

Биологические особенности и экология представителей сем. onagraceae (кипрейные) в мурманской области
На правах рукописи
Асминг Светлана Викторовна
Биологические особенности и экология представителей сем. Onagraceae (кипрейные)
в Мурманской области
03.00.05 – ботаника
Автореферат диссертации на соискание учёной степени кандидата
биологических наук
Петрозаводск
2007
Работа выполнена на кафедре биологии Кольского филиала Петрозаводского государственного университета
Научный руководитель
кандидат биологических наук, доцент,
Похилько Анна Антоновна
Официальные оппоненты:
доктор биологических наук, старший научный сотрудник,
Кузнецов Олег Леонидович,
институт биологии Кар НЦ РАН,
кандидат биологических наук, доцент,
Дьячкова Тамара Юрьевна,
Петрозаводский государственный университет
Ведущая организация
Полярно-альпийский ботанический сад-институт Кольского научного центра РАН
Защита диссертации состоится 28 ноября 2007 г. в 14 часов на заседании диссертационного совета Д 212.190.01 при Петрозаводском государственном университете по адресу: 185910, Республика Карелия, Петрозаводск, пр. Ленина, 33, эколого-биологический факультет, ауд. 137 теоретического корпуса.
С диссертацией можно ознакомиться в библиотеке Петрозаводского государственного университета
Автореферат разослан «___» октября 2007 г.
Ученый секретарь
диссертационного совета
Шкляревич Г.А.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования. Из 36 родов сем. Onagraceae, распространённых главным образом, в умеренной зоне в Мурманской области произрастают 3 – Chamaenerion (Rafin.) Rafin., Circaea L. и Epilobium L..
На территории Кольского полуострова встречается 1 вид рода Chamaenerion – Ch. angustifolium (L.) Scop., 1 вид рода Circaea – С. alpina L. и 8 видов рода Epilobium – E. adenocaulon Hausskn. (заносный вид), E. alpinum L., E. alsinifolium Vill., E. collinum C. C. Gmel., E. davuricum Fisch. ex Hornem., E. hornemannii Reichenb., E. lactiflorum Hausskn. и E. palustre L. Б. А. Мишкин (1953) выделяет также E. lapponicum (Wahlenb.) Hausskn.
Наиболее детально изучен Ch. angustifolium. Исследованием его номенклатуры, систематики, морфологии, географического распространения, онтогенеза, сезонного развития, способов размножения и распространения, экологии, фитоценологии, консортивных связей занимались многие отечественные и зарубежные ученые (Данилов, 1938; Забелкин, Уланова, 1995; Myerscough, 1980; Stockling, 1992 и др.).
В отличие от Chamaenerion 2 других рода изучены недостаточно полно. В частности, не исследованы онтогенез, возрастные спектры и жизненность популяций, морфолого-биологические особенности и экология этих растений. Ревизия рода Epilobium в Мурманской области не проводилась, что является актуальным в настоящее время.
Circaea alpina и 3 вида рода Epilobium (E. alsinifolium, E. davuricum, E. lactiflorum) являются редкими и нуждающимися в охране растениями на территории Мурманской области. Они занесены в Красные книги разных рангов: Красная книга Мурманской области (1979, 1990, 2004), Красная книга Архангельской области (1995), Red Data Book of East Fennoscandia (1998) и др.
Circaea alpina и некоторые виды рода Epilobium обладают полезными свойствами, являются лекарственными, кормовыми и декоративными растениями (Растительные ресурсы СССР…, 1987).
Circaea alpina, E. alsinifolium и E. davuricum – кальцифильные виды, указывающие на наличие пород с присутствием извести (Шляков, 1959; Kytvuori, 1962; Раменская, 1983). Эти растения могут служить индикаторами плодородия почв, способствовать выявлению ценных сообществ и мест концентрации редких видов при составлении региональных Красных книг.
Цель и задачи исследования. Целью работы является детальное изучение морфолого-биологических особенностей и экологии вышеназванных видов как в естественных местах произрастания, так и в условиях культуры. Особое внимание уделено Circaea alpina на северном пределе распространения этого чрезвычайно труднокультивируемого вида.
Для достижения этой цели необходимо было решить следующие задачи:
1) определить устойчивые морфологические признаки, характеризующие близкие виды, и уточнить таксон, к которому принадлежит тот или иной вид; 2) изучить онтогенез, возрастной состав и жизненность особей в популяциях Circaea alpina и видов рода Epilobium в естественных местах обитания и в условиях культуры; 3) определить экологический оптимум рассматриваемых видов и их географическое распространение в пределах Мурманской области; 4) изучить способы возобновления, семенную продуктивность, всхожесть и скорость прорастания семян.
Такие подробные исследования помогут выяснить причины редкости этих видов.
Научная новизна. Впервые изучены онтогенез, возрастные спектры, экологические особенности, семенная продуктивность и способы возобновления, всхожесть семян, скорость развития видов рода Epilobium и Circaea alpina. Определено современное состояние популяций изучаемых видов, подтверждены их местонахождения 30-40 гг. XX века, дан прогноз жизнеспособности особей в ценопопуляциях в регионе.
Практическая ценность. Дихотомический ключ для определения видов рода Epilobium представляет интерес для ботаников, работников природоохранных организаций, учителей, школьников. Изучение диагностических признаков, онтогенеза, строения монокарпического побега, возрастных спектров и жизненности ценопопуляций исследуемых видов вносит определённый вклад в систематику, биологию и экологию растений, результаты исследования могут использоваться в курсах для студентов биологических специальностей.
Исследование современного состояния популяций представителей сем. Onagraceae имеет большое значение для прогнозирования их жизнеспособности и сохранения популяций. Сведения о географическом распространении изученных растений могут быть внесены в базы данных по редким видам.
Материалы нашей работы могут быть использованы Комитетом природных ресурсов и окружающей среды Мурманской области при проектировании особо охраняемых природных территорий (ООПТ), проведении экологических экспертиз и при отведении участков земли под различные виды деятельности.
Апробация работы. Материалы диссертации доложены и обсуждены на конференции молодых учёных «Сбалансированное природопользование на примере освоения минеральных ресурсов» в 2004 г. (г. Апатиты), VII научной конференции Кольского филиала Петрозаводского государственного университета в 2004 г. (г. Апатиты), на Международной конференции Петрозаводского государственного университета «Структурно-функциональные особенности биосистем Севера» в 2005 г. (г. Петрозаводск).
Благодарности. Выражаю благодарность за терпеливое руководство – к.б.н., доценту кафедры биологии КФ ПетрГУ А.А. Похилько, за ценные советы - д.б.н., зав. лабораторией микробиологии Института проблем промышленной экологии Севера КНЦ РАН
Г.А. Евдокимовой, а также к.б.н., старшему научному сотруднику Полярно-альпийского ботанического сада-института КНЦ РАН О.А. Белкиной и к.б.н., научному сотруднику А.Ю. Лихачёву за определение видов мохообразных.
Публикации. По теме диссертации опубликовано 7 работ, в печати 1 работа.
Структура и объём работы. Диссертация состоит из введения, 4 глав, выводов, списка литературы, изложена на 109 страницах машинописного текста, содержит 38 таблиц и 51 рисунок. Список использованной литературы включает 86 источников, в том числе 21 иностранный.
1. ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Объектами исследования являются 8 видов рода Epilobium (E. adenocaulon, E. alpinum,
E. alsinifolium, E. collinum, E. davuricum, E. hornemannii, E. lactiflorum, E. palustre) и Circaea alpina (сем. Onagraceae).
Работа по изучению онтогенеза, систематики, распространения и экологии вышеназванных видов проводилась в течение 2002-2004 гг.
Использовались различные методы исследования: географический, систематический, онтогенетический, методы популяционного мониторинга, разработанные Л.Б. Заугольновой (1994), И.М. Ермаковой и др. (1987) и Т.Г. Соколовой (1987).
Материал собран в разных районах Мурманской области – на юго-западе (Кандалакшский район, окрестности пос. Вуориярви, ущелье Пюхякуру) и в центральной части (Кировский и Апатитский район).
Для составления карт ареалов изучаемых видов рода Epilobium и Circaea alpina в Мурманской области критически просмотрен гербарий ПАБСИ (PABGI), Ботанического Института им. В.Л. Комарова (LE), сборы студентов и аспирантов Кольского филиала Петрозаводского государственного университета в окрестностях с. Умба, пос. Сосновка и др., собственные сборы из различных районов Мурманской области и литературные данные (Мишкин, 1953; Шляков, 1959; Раменская, Андреева, 1982; Раменская, 1983; Белкина и др., 1991). Просмотрено около 1000 гербарных листов, более 4000 гербарных образцов. Карты общего распространения исследуемых видов приводятся по Хультену и Фрису (Hulten & Fries, 1986).
Возрастные спектры ценопопуляций видов рода Epilobium и Circaea alpinа изучались на постоянных и полустационарных площадках. При исследовании жизненного состояния ценопопуляций Circaea alpina использовались два метода: выделение трёх уровней жизненного состояния и многобалльная оценка жизненности (Ермакова и др., 1987). При изучении жизненности ценопопуляций видов рода Epilobium применялся только метод многобалльной оценки.
Во всех местах обитания закладывалось 20-30 площадок размером 1 м2, на которых подсчитывалось количество особей и собирался гербарный материал. Статистическая обработка проводилась в Microsoft Office Excel 2003.
Поскольку переход видов рода Epilobium из одного возрастного состояния в другое в прегенеративный период проходит быстро и эти виды являются вегетативно-подвижными растениями, все вегетативные группы – p, j, im, v были объединены в группу вегетативных, а все генеративные – g1, g2, g3 – в генеративную. Рассмотрены соответственно только эти группы.
Учитывались параметры популяций – проективное покрытие (%), обилие (по Друде), число особей разных возрастных групп (%), соотношение генеративных и вегетативных особей (%). Сделано около 200 геоботанических описаний в местах произрастания исследуемых видов. Названия видов сосудистых растений приведены по А.К. Черепанову (1995), мохообразных по
М.С. Игнатову, О.М. Афониной (1992).
Материал для изучения онтогенеза видов рода Epilobium и Circaea alpina собирался методом эколого-популяционной выборки. Из разных мест произрастания выбирались разновозрастные особи 20-30 экземпляров, которые фиксировались в 75%-м спирте или гербаризировались. Изучение онтогенеза проводилось в естественных местах обитания и в лабораторных условиях. Возрастные состояния выделялись по А.А. Уранову (1975). Описания соцветий приведены по А.А. Фёдорову, З.Т. Артюшенко (1979). Для изучения первых этапов онтогенеза в лабораторных условиях проведены опыты по определению всхожести семян
E. hornemannii и E. palustre, собранных в разных местах обитания. Семена E. hornemannii собраны в трёх местах обитания (берег оз. Имандра, Тик-губа; Хибинский горный массив, подножье горы Кукисвумчорр, берег ручья; Хибинский горный массив, берег ручья, влажное место), семена
E. palustre из двух мест обитания (предгорье Хибин; Хибинский горный массив, окрестности
г. Кировска, берег р. Белая в среднем течении; окрестности горы Белой (5 км на юго-запад от
ст. Апатиты, влажное болотистое место)).
Для изучения первых этапов развития семена (по 100 шт.) проращивали при комнатной температуре на свету, стратифицировали в течение 14 дней при -1-0оС. Морфология семян, ёмкость и строение почек изучалось по общепринятой методике с использованием светового микроскопа МБС–10, х56.
2. СИСТЕМАТИЧЕСКИЙ ОБЗОР ПРЕДСТАВИТЕЛЕЙ СЕМ. ONAGRACEAE, ПРОИЗРАСТАЮЩИХ В МУРМАНСКОЙ ОБЛАСТИ
Сем. Onagraceae содержит около 500 видов, 36 родов, встречающихся главным образом в умеренной зоне Нового Света, преимущественно в западных областях Северной Америки и Мексики. Некоторые роды приурочены к определенным странам. Только немногие из родов распространены по всему свету, сюда относится в первую очередь pод Epilobium, содержащий около 250 видов; это единственный род из сем. Onagraceae, который представлен в Австралии. В пределах бывшего СССР он является наиболее распространённым - 53 вида (Штейнберг, 1949). Меньшее распространение имеют: Chamaenerion, представленный 7 видами, Circaea – 6 видами и Ludwigia L. – 2 видами (Скворцов, 1989). Как заносное встречается Oenothera L. (Штейнберг, 1949; Скворцов, 1994).
На территории Мурманской области произрастают 3 рода: Chamaenerion, Circaea и Epilobium. Род Chamaenerion представлен 1 видом - Ch. angustifolium, который является наиболее изученным из представителей сем. Onagraceae.
Из 7 видов рода Circaea, свойственных северной умеренной и холодной зоне, в пределах бывшего СССР встречается 6 (Штейнберг, 1949). На территории региона представлен только 1 вид Circaea alpina (Шляков, 1959).
Ранее считалось, что многие виды рода Epilobium образуют гибриды, что затрудняет правильное определение вида (Syrinki, 1933; Штейнберг, 1949; Шляков, 1959). Однако монограф этого рода А.К. Скворцов (2005) считает, что виды рода Epilobium в своём большинстве имеют очень чёткие характерные признаки. По его мнению, в России гибриды встречаются не часто.
Авторы, ранее исследовавшие систематику изучаемых нами видов, – Е.И. Штейнберг (1949), Р.Н. Шляков (1959), А.К. Скворцов (1980), Д.П. Воробьёв (1981), М.Л. Раменская (1982), Н.В. Власова (1996) – применяли для определения разные признаки: форма рыльца, длина черешков, форма и поверхность семян, органы перезимовки и возобновления, наличие/отсутствие на стебле листовых линий, форма листьев. В работе представлено подробное сравнение ключей разных авторов для определения исследуемых видов, их достоинств и недостатков.
Проведён анализ морфологических и других диагностических признаков, характеризующих тот или иной вид рода Epilobium. Для определения видов рода Epilobium, произрастающих на территории Мурманской области, нами предложен дихотомический ключ, который дополняет ключи Р.Н. Шлякова (1953) и А.К. Скворцова (1980, 1991) и подтверждает диагностические признаки, применяемые другими авторами.
В основу ключа, как и у других монографов этого рода, положена форма рыльца.
Виды рода Epilobium, произрастающие на территории Мурманской области, обладают как устойчивыми диагностическими признаками, так и дополнительными.
Устойчивыми признаками являются: ветвление и высота побега (для E. alpinum); органы перезимовки и возобновления (E. alsinifolium, E. davuricum, E. hornemannii, E. palustre) в период их формирования; опушение верхушки стебля, цветоножки, чашечки и коробочки (E. adenocaulon); расстояние между узлами для E. alsinifolium и E. hornemannii; форма и размеры листьев
(E. alpinum, E. alsinifolium, E. davuricum, E. hornemannii, E. lactiflorum и E. palustre); длина и ширина черешка (E. alsinifolium, E. hornemannii); форма рыльца (E. collinum); окраска и опушение коробочки (E. davuricum и E. palustre) в период плодоношения; поверхность семян
(E. adenocaulon, E. alsinifolium и E. hornemannii) в период рассеивания семян.
К дополнительным признакам относятся: ветвление и высота побега (E. davuricum и
E. palustre), окраска растения (E. davuricum, E. lactiflorum и E. palustre); характер листорасположения (E. alsinifolium и E. hornemannii); размеры цветков (E. alsinifolium,
E. hornemannii и E. lactiflorum) в период цветения; окраска цветков (E. davuricum, E. lactiflorum и E. palustre) в период цветения; форма и размеры семян (E. davuricum и E. palustre) в период рассеивания семян.
3. ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ, ФИТОЦЕНОЛОГИЯ И ЭКОЛОГИЯ ПРЕДСТАВИТЕЛЕЙ СЕМ. ONAGRACEAE
Рассмотренные виды относятся к разным географическим элементам. Представители родов Chamaenerion, Circaea, 2 вида рода Epilobium являются бореальными, в том числе 3 циркумполярные (Ch. angustifolium, Circaea alpina, E. palustre), 1 североамериканский
(E. adenocaulon). 3 вида рода Epilobium – гипоарктические (циркумполярные E. davuricum,
E. hornemannii и европейский E. alsinifolium), 1 – плюризональный европейский (E. collinum), 1 - арктический амфиатлантический (E. lactiflorum) и 1 арктоальпийский евразиатский (E. alpinum).
В Мурманской области из сем. Onagraceae довольно часто встречаются Ch. angustifolium,
Е. palustre и Е. hornemannii, произрастающие практически во всех районах. E. hornemannii и
Е. palustre часто растут вместе, являются широко распространенными и неприхотливыми видами, но Е. hornemannii обладает более широкой экологической амплитудой и большими биологическими возможностями. Этот вид часто можно встретить на нарушенных местах обитания, в более сухих по отношению к влаге, т. е. потенциальные биологические возможности Е. hornemannii выше, чем у Е. palustre.
Помимо широко распространённых представителей сем. Onagraceae на территории Мурманской области есть редкие виды – Circaea alpina, Е. alsinifolium и Е. davuricum, приуроченные к определённым условиям (в частности к выходам карбонатных пород). Перечисленные виды, а также E. alpinum и E. lactiflorum весьма требовательны к целостности окружающей среды и чувствительны к антропогенной нагрузке, произрастают в местах, удалённых от человеческой деятельности. Кроме вышеупомянутых видов, на территорию Кольского полуострова из Карелии заходит более южный представитель рода Epilobium –
E. collinum и заносный североамериканский E. adenocaulon.
Если рассматривать распространение в Мурманской области рода Epilobium в целом, можно сделать вывод о том, что он встречается и в центральной части и на периферии области (рис. 1). Распространение рода Epilobium приурочено практически ко всем горным массивам, так как среди его видов есть арктические (E. lactiflorum), арктоальпийские (E. alpinum) и гипоарктические (E. alsinifolium, Е. davuricum, Е. hornemannii), предпочитающие нивальные луговины, места позднего схода снега. Многие виды рода Epilobium обладают гидроморфными признаками. В восточном и северо-западном районах видов рода Epilobium описано меньше.
Большинство видов рода Epilobium сконцентрировано в центральной части области (от 3 до 6 видов), на юго-востоке (3-5) и на севере (2-5). В остальных районах число видов колеблется от 1 до 3. В некоторых местах вообще не отмечено присутствие видов рода Epilobium (рис. 1).
Виды рода Epilobium - это растения первичных и вторичных открытых местообитаний как естественного происхождения (ключевые болота, берега рек, ручьёв, нивальные луговины), так и искусственного. Ненарушенные, первичные местообитания, на которых отмечено присутствие видов рода Epilobium, встречаются на территории Мурманской области в альпийском и субальпийском горных поясах. Вместе с ними произрастают виды рода Salix (S. caprea,
S. phylicifolia, S. starkeana), Agrostis (A. canina, A. stolonifera), Carex (C. bigelowii, C. cespitosa,
C. cinerea, C. vesicaria), Equisetum (E. palustre, E. pratense, E. sylvaticum), Galium (G. palustre,
G. uliginosum), Poa (P. pratensis, P. trivialis), Cirsium heterophyllum, Crepis paludosa, Deschampsia cespitosa, Elymus caninus, Montia fontana, Saussurea alpina, Solidago lapponica, а также мохообразные - Bryum pseudotriquetrum, Calliergon stramineum, Dicranella palustris, Palustriella commutata, Philonotis fontana, виды рода Rhyzomnium (R. magnifolium, R pseudopunctatum), Sphagnum fuscum и др. В лесах виды рода Epilobium приурочены к влажным местам близ ручьёв, рек. Здесь также встречаются лесные виды – Vaccinium vitis-idaea, виды рода Melampyrum
(M. pratense, M. sylvaticum), Orthilia secunda, Pyrola rotundifolia, Trientalis europaea и др.
В специфических местах обитания (эвтрофное болото южного Прихибинья, ущелья Пюхякуру и Рускеакуру) виды рода Epilobium произрастают с кальцифильными видами, являющимися редкими на территории Мурманской области: Circaea alpina, E. davuricum, Equisetum scirpoides и др.

Рис. 1. Частота встречаемости видов рода Epilobium в Мурманской области
(0-7 – число видов)
В местах, подверженных антропогенной нагрузке, совместно с видами рода Epilobium растут: Deschampsia cespitosa, Lepidotheca suaveolens, Potentilla norvegica, Rumex confertus, Taraxacum officinale, Tussilago farfara, которые во флоре Мурманской области относятся к адвентивным.
Зафиксировано около 100 видов высших сосудистых растений и 25 видов мохообразных, произрастающих совместно с видами рода Epilobium.
Широко распространённые E. hornemannii и E. palustre часто растут вместе в схожих местах обитания, поэтому набор сопутствующих видов у них почти одинаков. Господствуют травянистые многолетние растения и мохообразные. Эти 2 вида весьма часто сопутствуют редким видам этого рода (E. alsinifolium, E. davuricum, E. lactiflorum) и Circaea alpina.
Circaea alpina занимает в семействе обособленное положение, имеет ограниченное распространение в Андах, происхождением связана с неморальными лесами (Раменская, 1983). В области известно только одно место нахождения Circaea alpina – на юго-западе Кандалакшского района (ущелья Пюхякуру и Рускеакуру, территория комплексного регионального заказника «Кутса»). Circaea alpina – мезофит, сциофит, кальцефил. Этот вид обладает узкой экологической амплитудой, произрастает в лесах определённого типа – тенистых темнохвойных мшистых лесах, в пойменных кустарниках, по замшелым склонам, расщелинам скал. Circaea alpina – вид первичных ненарушенных местообитаний естественного происхождения (разнотравно-моховые прирусловые ивняки, ельники папоротниково-разнотравно-зеленомошные). Предпочитает среднее и избыточное увлажнение, растёт по берегам ручьёв, на затенённых влажных скалах, на влажной перегнойной почве в тенистых сырых местах (Попов, 1957; Шляков, 1959; Скворцов, 1974; Беглянова, 1979; Воробьёв, 1981; Рысин, Рысина, 1987; Власова, 1996). На это растение оказывают резко отрицательное влияние уплотнение почвы и механическое повреждение наземных органов (Рысин, Рысина, 1978).
Проективное покрытие Circaea alpina на затенённых местах достигает 65-85%, на более освещённых – 35-40%. Особи формируют корневище и столоны, захватывая небольшие пространства.
Сопутствующими видами Circaea alpina являются Salix spp., Athyrium filix-femina, Dryopteris carthusiana, Matteuccia struthiopteris, другие представители сем. Onagraceae
(Ch. angustifolium, E. hornemannii, E. palustre), Equisetum pratense, Comarum palustre, Filipendula ulmaria, Geum rivale, Actaea erythrocarpa, Caltha palustris и др. Всего выявлено 38 видов сосудистых растений, произрастающих в этом фитоценозе.
Circaea alpina растёт на моховых подушках, в моховом покрове наблюдается разнообразие мохообразных (25 видов): Aulacomnium palustre, Brachythecium latifolium, Bryum pseudotriquetrum, Climacium dendroides, Martschantsia polymorpha, Mnium stellare, Plagiomnium ellipticum, Plagiothecium denticulatum, Pleurozium schreberi, Pohlia wahlenbergii, Polytrichastrum alpinum, Pseudobryum cinclidioides, Rhodobryum roseum, Rhytidiadelphus triquetrus, Sphagnum warnstorfii, Warnstorfia fluitans и др. Проективное покрытие сосудистыми растениями составляет от 50 до 95%, мохообразными от 70 до 100%. Общее проективное покрытие 70-100%.
4. ОНТОГЕНЕЗ, СТРОЕНИЕ МОНОКАРПИЧЕСКОГО ПОБЕГА, ВОЗРАСТНОЙ СОСТАВ, ЖИЗНЕННОСТЬ ПОПУЛЯЦИЙ И СЕЗОННОЕ РАЗВИТИЕ CIRCAEA ALPINA И ВИДОВ РОДА EPILOBIUM
4.1. Онтогенез, строение монокарпического побега, возрастной состав, жизненность особей в популяции и сезонное развитие Circaea alpina
4.1.1. Онтогенез Circaea alpina
Circaea alpina – многолетнее травянистое растение с укороченным гипогеогенным корневищем, в узлах которого образуются почки возобновления и столоны. На северном пределе распространения (Мурманская область) Circaea alpina, с нашей точки зрения, меняет жизненную форму. Некоторые авторы, занимавшиеся ранее Circaea alpina, относят её к столонно-клубневым растениям с небольшим жизненным циклом (Рысин, Рысина, 1987; Барыкина, 2000). Наши исследования показывают, что на территории региона Circaea alpina является многолетним растением, подземные органы которого в крайних условиях существования сохраняются в течение нескольких лет.
Семена с прямым зародышем без эндосперма. Сведения о прорастании семян этого вида нам не известны. Семена Circaea lutetiana при температуре 20оС почти не прорастают ни на свету, ни в темноте. Вероятно, необходима скарификация и холодная стратификация. Семена с сильным экзогенным и глубоким физиологическим покоем (Николаева, Разумова, 1985; Kinzel, 1913). Эксперименты по всхожести свежесобранных семян Circaea alpina не дали положительных результатов.
Проростки (p) Circaea alpina имеют первичный корешок и две семядоли овально-яйцевидной формы, которые не отмирают после образования настоящих листьев в течение
4-6 дней (рис. 2). На конусе нарастания закладываются листовые зачатки. К моменту развёртывания первых листьев семядоли 0.2-0.25 см длины, 0.15-0.2 см ширины, овальной (яйцевидной) формы. Длина гипокотиля варьирует от 1.5 до 3.5(4.5) см. Зародышевый корешок функционирует до перехода растения в имматурное состояние.
Переход в ювенильное (j) состояние характеризуется образованием первой пары настоящих листьев (рис. 2). Ювенильные растения Circaea alpina – однопобеговые, длина побегов 2.5-4.5 см. Длина междоузлия варьирует от 0.7 до 1.5 см. На этой стадии формируются первая и вторая пары настоящих листьев. Первая пара 0.2-0.25 см длиной, 0.15-0.2 см шириной, овальной формы, цельнокрайные, на длинном черешке. Вторая пара 0.5-(0.8) см длиной, 0.4-(0.7) см шириной, яйцевидной (широкояйцевидной) формы, цельнокрайные, черешок короче, чем у первой пары. Главный корень сохраняется, формируются боковые корни первого порядка.
Побеги имматурных (im) особей Circaea alpina длиной 3.8-4.5 см, с 2 междоузлиями и
3 парами листьев (рис. 2). Листья на длинных черешках, становятся похожи на взрослые, приобретая сердцевидную форму. В зависимости от расположения на побеге варьируют размеры и форма листьев, количество зубцов. Первая пара листьев 0.2-0.25 см длиной, 0.15-0.2 см шириной, овальной формы, цельнокрайные, на длинном черешке. В основании первого узла начинают формироваться 2 боковых побега 1-2 см длиной, 2 пары листьев. Вторая пара листьев 0.8-1 см длиной, 0.7-(1) см шириной, яйцевидной (широкояйцевидной) формы с 2 зубцами. Происходит отмирание главного корня, образуется гипогеогенное корневище 0.4-1.0 см длиной, 1.5-2.5 см толщиной. В пазухах листьев и на корневище закладываются почки возобновления (1-2).
Переход в виргинильное (v) состояние сопровождается интенсивным нарастанием надземной и подземной частей растений (рис. 2). Виргинильные особи Circaea alpina имеют побеги длиной 6.5-10 см и 3 междоузлия. Формируются боковые побеги второго порядка (1-2). Строение боковых побегов идентично главному. Листья в количестве 3-4 пар, крупные, сердцевидно-яйцевидной формы, имеют хорошо выраженные зубцы по всему краю: первая пара листьев овальной-яйцевидной формы, 0.6–1.3 см длиной, 0.5-1.0 см шириной; вторая и последующие пары листьев сердцевидной-яйцевидной формы с 3-4(5) зубцами и выемкой у основания, (0.5)0.9-1.1(1.3) см длиной, (0.5)0.7-1(1.2) см шириной. Длина корневища 0.7-2.5 см, толщина 1.6-2.5 см. В отличие от имматурных, у виргинильных особей на корневище, состоящем из 2-4(5) междоузлий, образуются уже не единичные, а многочисленные почки возобновления, которые закладываются равномерно на всём протяжении корневища (по 1 в каждом узле). У второго междоузлия появляются придаточные корни (0.5-1 см длиной).
С образованием первого генеративного побега растения Circaea alpina переходят в генеративное состояние.
Молодые генеративные (g1) особи Circaea alpina имеют побеги длиной 3.5-10 см и
3-4 междоузлия (рис. 3). Листья в количестве 3-4 пар, крупные, с хорошо выраженными зубцами по краю. Первая пара листьев отмирает, в их пазухах развиваются боковые побеги. Вторая и последующие пары листьев сердцевидной-яйцевидной (широкояйцевидной) формы, 3-4-зубчатые, с выемчатым основанием, (0.7)1.3-1.5 см длиной, (0.8)1.3-0.9(1.5) см шириной. Развиваются
1-2 боковых побега 4 см длиной, несущие по 3–4 пары листьев. На верхушке побега формируется простое соцветие – кисть (средней длины, пяти-многоцветковая, редкая, верхушечная, прямая, цилиндрическая). На корневище, помимо почек возобновления, образуются столоны. Столоны подземные, состоят из нескольких пар мясистых бесцветных чешуй (2-3), междоузлий (2-3) и верхушечной почки. Длина столонов варьирует от 7.5 до 13.5 см. В пазухах чешуй столона находятся почки возобновления (1-2). Верхушечная почка столона состоит из чешуй (2), листовых бугорков (2) и конуса нарастания.

Рис. 2. Особи Circaea alpina в прегенеративный период (бп – боковой побег,
вп – верхушечная почка, г/к – гипокотиль, зк – зародышевый корешок, к – корневище,
м – междоузлия, о/л – отмершие листья, пк – придаточные корни, с/д – семядоли, э/к – эпикотиль, 1 л – первая пара листьев, 2 л – вторая пара листьев и т. д.)
Средневозрастные (g2) особи. Побеги длиной 6.5-12.5 см, имеют 5-7 междоузлий (рис. 3). На верхушке побега сформировано кистевидное соцветие, состоящее из главной оси и 2-4 боковых паракладиев, каждый из которых несёт по (5)6-8(10) цветков. Листорасположение может быть как супротивным, так и очередным. Листья в количестве 4-5 пар, крупные, сердцевидно-яйцевидной формы, имеют хорошо выраженные зубцы по краю листа: первая пара листьев отмерла; вторая пара листьев 0.6-0.7 см длиной, 0.7-0.9 см шириной, листья широкояйцевидной формы, цельнокрайные; третья и последующие пары листьев (2.3)3-(2.9)4 см длиной, (1.8)2.5-(2.1)3.1 см шириной, сердцевидно-яйцевидной формы, с (4)5(6) зубцами, с выемчатым основанием. Длина корневища 0.9-3.0 см, толщина 1.8-2.7 см. На корневище продолжают формироваться почки возобновления и столоны длиной 6-9 см, имеющие по 7 междоузлий и 3-5 пар плёнчатых листьев. Происходит вегетативное размножение. За счёт образования столонов особь разрастается, увеличивается занимаемая ею площадь. В следующий вегетационный период столоны отмирают, связь с материнским растением прерывается, дочерние особи начинают самостоятельное существование.
У старых генеративных (g3) особей Circaea alpina начинается упрощение структуры надземных и подземных частей. Побеги длиной 5.5-10.5 см с 3-4 междоузлиями. Листья в количестве 4(5) пар, менее крупные, чем у средневозрастных особей, сердцевидно-яйцевидной формы, зубцы по краю листа становятся менее выраженными: первая пара листьев отмерла; вторая и последующие пары (2.1)2.7-(2.7)3.8 см длиной, (1.5)2.3-(1.9)3.8 см шириной, сердцевидно-яйцевидной формы, с (3)4(5) зубцами, с выемчатым основанием. Длина корневища 0.7-2.5 см, толщина 1.6-2.5 см. Отмирают старые участки корневища, столоны продолжают формироваться, но уже в меньшем количестве (1-2).
Сенильные (s) и субсенильные (ss) особи Circaea alpina в природе не обнаружены. Это объясняется тем, что Circaea alpina является вегетативно-подвижным растением. Благодаря наличию корневищ и формирующимся на них столонам особи постоянно омолаживаются и расселяются по территории.

Рис. 3. Генеративные особи Circaea alpina (бп – боковой побег, к – корневище,
м – междоузлия, о/л – отмершие листья, пк – придаточные корни, пл – плоды, с – соцветие,
ст – столоны, ц – цветки, 1 л – первая пара листьев, 2 л – вторая пара листьев и т. д.)
4.1.2. Строение монокарпического побега Circaea alpina
У особей разных возрастных состояний Circaea alpina во второй половине вегетационного сезона (конец июля – начало августа) начинают формироваться почки возобновления (рис. 4). В количестве от 4 до 10 они располагаются в пазухах бесцветных пленчатых чешуй на коротком гипогеогенном корневище, состоящем из нескольких междоузлий (2-4(5)). Наиболее хорошо сформированная почка находится у первого междоузлия корневища, в пазухе первой пары пленчатых чешуй; здесь же появляются столоны. Закладывается по 2 почки, но развивается обычно одна. Она состоит из бесцветных чешуй (1-2) с широкими основаниями, 2 листовых зачатков и конуса нарастания. Размеры почек: 0.35-1.5 мм длина и 0.2-0.8 мм ширина.
Почки возобновления после периода покоя в следующий вегетационный сезон дают начало новым метамерам подземных побегов с чешуевидными листьями. К концу августа рост подземных побегов замедляется, на корневище развиваются придаточные корни, а в пазухах чешуевидных листьев формируются почки возобновления (1-3). Весной из почки формируется надземный побег с листьями, расположенными супротивно. Побег заканчивается соцветием. Цветоносный побег осенью отмирает, его базальная часть включается в систему подземных корневищ, нарастающих симподиально.
У основания надземных побегов находятся почки столонов (1-3). Столон состоит из нескольких пар чешуй (2-3), междоузлий (2-3) и верхушечной почки. Длина столона 7-13.5 см.
В пазухах чешуй столона расположены почки (1-2). Верхушечная почка состоит из чешуй (2), листовых бугорков (2) и конуса нарастания.
Таким образом, многолетний поликарпик Circaea alpina формирует побеги дициклического типа, т. е. длительность их развития от пробуждения начальной почки до цветения и отмирания составляет 2 вегетационных периода.

Рис. 4. Строение почек и столонов Circaea alpina
4.1.3. Возрастной состав популяций Circaea alpina
Популяция Circaea alpina в ущелье Пюхякуру разделена озером на 2 части – западную и восточную. Они расположены относительно недалеко друг от друга (расстояние 500 м), располагаются не на одном уровне. Западная часть популяции занимает площадь 15 м2, восточная – 10 м2. Расселение вида идет сверху вниз по ущелью (с запада на восток).
Возрастной спектр восточной части популяции двухвершинный, неполночленный: отсутствуют проростки (p), сенильные (s) и субсенильные (ss) особи, преобладают генеративные – 54%. Среди особей прегенеративного периода господствуют виргинильные (v) – 25%, среди генеративных средневозрастные (g2) – 31%. Значительно меньше молодых (g1) – 7% и старых генеративных особей (g3) – 16%. В процентном соотношении молодых генеративных и средневозрастных особей больше, чем старых генеративных. Плотность восточной части популяции составляет 38 шт. на 1 м2. В целом в восточной части популяции прегенеративная и генеративная часть примерно одинаковы (46 и 54% соответственно) (рис. 5).
Возрастной спектр западной части популяции существенно отличается от возрастного спектра восточной присутствием проростков (p) – 2%, большим процентом ювенильных (j) – 16%, имматурных (im) – 20% и виргинильных (v) – 39% особей. Генеративная часть представлена главным образом средневозрастными особями (g2) – 13%, процент молодых и старых генеративных одинаков (по 5%), сенильные (s) и субсенильные (ss) особи отсутствуют. Плотность западной части популяции составляет 34 шт. на 1 м2. В целом в западной части популяции преобладают особи прегенеративного периода (78%).

Рис. 5. Возрастные спектры популяции Circaea alpina (Кандалакшский район, окрестности пос. Вуориярви, ущелье Пюхякуру)
Таким образом, в возрастном спектре восточной части популяции Circaea alpina преобладают генеративные (54%), а в западной – особи прегенеративного периода (81%), т. е. последняя моложе. Популяции располагаются не на одном уровне, и поэтому расселение идёт сверху вниз по ущелью.
Так как обе части популяции Circaea alpina расположены относительно недалеко друг от друга, они были объединены в одну. Тогда возрастные спектры выглядят следующим образом. Возрастные спектры популяции Circaea alpina в Мурманской области неполночленные, двухвершинные, левосторонние: господствуют особи прегенеративного периода (v) – 32%. Приблизительно на одном уровне находятся ювенильные (j) – 11%, имматурные (im) – 17%, средневозрастные (g2) – 15% и старые генеративные особи (g3) – 18%. Проростки (p) и молодые генеративные особи (g1) составляют небольшую долю спектра (1 и 6%), сенильные (s) и субсенильные (ss) особи отсутствуют. Плотность популяции 37 шт. на 1 м2. В целом в популяции Circaea alpina в Мурманской области преобладают особи прегенеративного периода (61%).
Важной оценкой состояния ценопопуляции является её виталитетный спектр. В ценопопуляциях Circaea alpina господствуют особи прегенеративного периода. Отсутствие субсенильных и сенильных особей, вероятно, можно объяснить тем, что Circaea alpina является вегетативно подвижным растением и при появлении корневищ и столонов происходит образование новых побегов и особей. В изученной ценопопуляции наибольшее число растений средних размеров.
Изученная локальная популяция разновозрастная с относительно высокой жизненностью. Преобладание особей в вегетативном состоянии говорит о семенном возобновлении Circaea alpina в естественных местах произрастания, а также о расселении этого вида. В составе ценопопуляции значительную роль играют способные к плодоношению генеративные растения.
При сравнении восточной и западной частей ценопопуляции выявляются возможности вида к расселению: идёт распространение семян по ущелью с востока на запад, возможно, потоками воды, при таянии снега и при выпадении атмосферных осадков.
Более высокая выживаемость молодых растений в западной части популяции в сочетании с низкой репродуктивностью (низкое число вегетативных растений в расчёте на одно генеративное), вероятно, объясняется её недавним появлением. В восточной, более старой части низкое выживание молодых растений сочетается с относительно большей репродуктивностью, это приводит к снижению плотности популяции.
Тип возрастного спектра ценопопуляции Circaea alpina в оптимальных условиях является равновесным и соответствует нормальному типу ценопопуляции. Изученная популяция достаточно стабильна.
4.1.4. Жизненность особей в популяции Circaea alpina в Мурманской области
Для изучения жизненности особей в популяции были использованы два метода.
1. Выделение трёх уровней жизненного состояния в онтогенезе особей и применение этого метода для характеристики популяции Circaea alpina.
Первый уровень – низкая жизненность. Растения с небольшими размерами побегов (высота побега 1.3-4.5 см) и листьев (длина листовой пластинки 0.45-1.3 см, ширина листовой пластинки 0.3-0.9 см), с небольшим количеством листьев (2-6) и репродуктивных органов (1 цветок/плод), отсутствием боковых побегов.
Второй уровень – средняя жизненность. Растения имеют нормальные размеры побегов (высота побега: 2.5-13.0 см) и листьев (длина листовой пластинки 0.75-2.6 см, ширина листовой пластинки 0.55-1.9 см), большое количество листьев (4-11), репродуктивных органов
(4-6 цветков/плодов) и боковых побегов (1-2).
Третий уровень – высокая жизненность. Растения с максимальными размерами побегов (высота побега: 5.5-18.0 см) и листьев (длина листовой пластинки 1.5-3.4 см, ширина листовой пластинки 1.0-2.5 см), с большим количеством листьев (8-14), репродуктивных органов
(8-10 цветков/плодов) и боковых побегов (1-3).
В исследованной ценопопуляции Circaea alpina в Мурманской области во всех возрастных группах преобладают особи с максимальным уровнем жизненности. Большую часть ценопопуляции составляют растения средних и максимальных размеров, то есть 2-го и 3-го уровней жизненности (рис. 6). Плотность особей в среднем 7.6 шт/м2.

Рис. 6. Возрастной спектр популяции Circaea alpina в Мурманской области с учётом дифференциации возрастных групп по уровням (1-3-й) жизненности особей
В прегенеративном периоде самая высокая жизненность наблюдается у виргинильных растений (62% – 3-го, 33% – 2-го и 5% – 1-го уровня жизненности) и наименьшая у ювенильных (36% – 3-го, 43% – 2-го и 21% – 1-го уровня жизненности). В генеративном периоде более мощными являются средневозрастные растения (76% – 3-го, 21% – 2-го и 3% – 1-го уровня жизненности), наименьшая жизненность у молодых (44% – 3-го, 44% – 2-го и 12% – 1-го уровня жизненности).
Жизненность особей прегенеративной и генеративной части одинаковая. Особи 3-го и 2-го уровня жизненности составляют 52-52 и 37-42% соответственно. В целом можно сказать, что популяция находится на оптимальном уровне существования, в нормальных условиях окружающей среды.
2. Метод многобалльной оценки жизненности особи и его применение для характеристики популяции Circaea alpina.
При втором подходе к изучению жизненного состояния особей Circaea alpina разработана трёхбалльная шкала, которую можно в дальнейшем использовать для вычисления различных критериев жизненности всей ценопопуляции. В основу шкалы было взято 4 признака – высота побега, количество листьев, длина и ширина листовой пластинки, а также количество репродуктивных органов (цветков/плодов) для генеративных особей, по которым были проанализированы особи всех возрастных групп.
В ценопопуляции преобладают особи со 2-м баллом жизненности, меньше особей с 1-м баллом и очень мало с 3-м баллом, причем по всем возрастным группам. Если рассматривать отдельно особей прегенеративного периода, то наибольшей жизненностью обладают ювенильные особи (j), наименьшей – виргинильные (v). Особи прегенеративной и генеративной части показывают примерно одинаковый разброс по уровням жизненности, у генеративных много особей со 2-м баллом, меньше с 1-м и с 3-м, а у вегетативных тоже много со 2-м, но больше с 1-м и меньше с 3-м.
Средний балл жизненности выражает средний уровень жизненности популяции и отдельных ее возрастных состояний, он позволяет представить средний облик растений Circaea alpina. Самым большим средним баллом жизненности обладают генеративные особи (2.0), самым маленьким – ювенильные (j) (1.7), у особей остальных групп он одинаков (1.8).
Оба метода оценки жизненности особей в популяциях показали, что жизненное состояние популяции Circaea alpina в Мурманской области невысокое. В целом популяция обладает средним баллом жизненности, т. е. она и не процветает и не угнетена – находится в пределах нормы. Конечно, говорить об оптимальных условиях существования этого вида нельзя, так как Мурманская область является северным пределом его распространения.
4.1.5. Сезонное развитие Circaea alpina в Мурманской области
Circaea alpina – летнезелёное растение. Вегетация Circaea alpina начинается после таяния снега в конце мая – начале июня (иногда в середине июня). Период цветения небольшой и длится всего 10 дней в годы с высокой температурой воздуха, как в 2003 году. Первые зрелые семена появляются в конце первой декады августа. Созревание плодов и семян идёт в течение 10-15 дней. Семена созревают не каждый год. Отмирание растений происходит в конце августа – начале сентября, но может быть и раньше, в зависимости от погодных условий.
4.2. Онтогенез, строение монокарпического побега, возрастной состав, жизненность особей в популяциях и сезонное развитие видов рода Epilobium
4.2.1. Всхожесть семян видов рода Epilobium
Скорость и длительность прорастания семян у исследованных видов рода Epilobium различны. Семена всех видов прорастают на 7-9-й день, максимальный процент всхожести наблюдается на 15-25-й у нестратифицированных семян и на 69-90-й у семян, подвергавшихся действию низких температур.
Высокой всхожестью обладают семена E. hornemannii во всех вариантах опыта, собранные в разных местах обитания (20-75% проросших семян). Стратификация низкими температурами отрицательно влияет на всхожесть семян этого вида (1-32%). Семена E. palustre, по сравнению с E. hornemannii, имеют низкую всхожесть (2-7%) в опытах при комнатной температуре, их всхожесть повышалась после воздействия низкими температурами (25-56%). В природе семена видов рода Epilobium прорастают на следующий год.
В результате поставленного нами эксперимента по всхожести семян видов рода Epilobium было установлено, что семена сохраняют всхожесть более одного года – от 1 до 3 лет с изменением количества проросших семян с 50-70 % до 20-35 %.
4.2.2. Онтогенез видов рода Epilobium
Ввиду того, что возрастное развитие разных видов рода Epilobium незначительно отличается, онтогенез рассматривается на примере одного вида – E. hornemannii. Для других видов указаны лишь существенные характерные отличия.
E. hornemannii – многолетнее травянистое растение с коротким гипогеогенным корневищем, в узлах которого образуются почки возобновления. В отличие от E. hornemannii, корневище E. alpinum удлинённое (ползучее). У E. alsinifolium и E. palustre на корневище помимо почек возобновления, образуются также специализированные органы вегетативного размножения – столоны (подземные у E. palustre и надземные у E. alsinifolium).
Проростки (p) имеют белый тонкий зародышевый корешок, длиной 14-31 мм и две семядоли, которые не отмирают после образования настоящих листьев в течение 5-7 дней. Зародышевый корешок функционирует до перехода растения в имматурное состояние. Семядоли овальной формы, 0.8-1.2 мм длины, 0.6-1 мм ширины. Длина гипокотиля 6.3-16.3 мм. На конусе нарастания закладываются листовые зачатки. Проросток не превышает по высоте 20 мм, обычно – 9-18 мм.
Проростки E. hornemannii, а также E. palustre появляются и в ненарушенных местах обитания, и в подверженных антропогенной нагрузке. Одним из главных условий для прорастания является достаточное увлажнение почвы. Проростки других видов (E. alpinum, E. alsinifolium,
E. davuricum, E. lactiflorum), более требовательных к целостности окружающей среды, встречаются только на средненарушенных участках, в открытых достаточно увлажненных местах с умеренным плодородием почвы и малым количеством конкурентов.
Переход в ювенильное (j) состояние характеризуется образованием первой пары настоящих листьев. Ювенильные растения E. hornemannii – однопобеговые, длина побегов 4.5-6.5(8.5) см. Первое и второе междоузлия укорочены (1.5-2.5 мм). На этой стадии формируются первая и вторая пары настоящих листьев. Первая пара овальной формы, с цельным краем, на коротком черешке, 0.2-0.25 см длиной, 0.1-0.15 см шириной. Вторая пара яйцевидной формы, с цельным краем, 0.5-0.8 см длиной, 0.4-0.6(0.7) см шириной. Листья располагаются супротивно. Главный корень сохраняется, формируются боковые корни первого порядка длиной 3.5-5.5 см.
Побеги имматурных (im) особей длиной (6.5)8.0-10.5 см с 2 междоузлиями и 3 парами листьев. Листорасположение меняется с супротивного на спиральное, кроме E. alsinifolium,
E. davuricum и E. palustre. Листья на длинных черешках (1.5-2.5 мм длиной), становятся похожи на взрослые, приобретая яйцевидную форму. В зависимости от расположения на побеге варьируют размеры и форма листьев. Первая пара листьев овальной формы, цельнокрайные, 0.4-0.5 см длиной, 0.25-0.35 см шириной. Вторая и последующие пары листьев 1.5-2.5 см длиной, 0.8-1.2 см шириной, эллиптической (яйцевидной) формы. У основания главного побега в пазухах первой и второй пары листьев закладываются почки возобновления (1-2). Главный корень отмирает, образуется гипогеогенное корневище 0.4-1.0 см длиной, 1.5-2.5 см толщиной.
На ранних этапах онтогенеза у имматурных особей всех исследованных видов рода Epilobium форма листьев отличается и является постоянным диагностическим признаком: линейная (E. davuricum), узколанцетная (ланцетная) (E. palustre), широколанцетная
(E. adenocaulon), овальная с закруглённой верхушкой (E. alpinum), яйцевидная (E. alsinifolium,
E. collinum).
В отличие от всех остальных видов, у особей E. alpinum корневище не короткое, а длинное (ползучее).
Переход в виргинильное (v) состояние сопровождается интенсивным нарастанием надземной и подземной частей растений. Виргинильные особи E. hornemannii имеют побеги длиной 5.0-12.0 см и 3-4 междоузлия. Листья в количестве 3-4 пар, вполне сформированы, приобретают взрослую форму. Первая пара листьев овальной формы, цельнокрайные, 0.5-2.0 см длиной, 0.3-1.0 см шириной. Вторая и последующие пары листьев яйцевидной формы, 0.65-2.40 см длиной, 0.25-1.30 см шириной. Появляется хорошо выраженное сетчатое жилкование листьев. Идёт нарастание корневища – длина 0.5-1.0 см, толщина 0.8-1.5 см. В отличие от имматурных, у виргинильных особей на корневище образуются уже не единичные, а многочисленные почки возобновления. У второго междоузлия появляются придаточные корни (0.5-1 см длиной).
Средняя длина побега у других видов: 1.5-4.5 см (E. alpinum), 5.0-10.5 см (E. davuricum), 6.0-11.0 см (E. lactiflorum), 10.5-20.0 см (E. palustre). Также отличаются форма и размеры листьев: линейные 0.69 см длиной, 0.17 см шириной (E. davuricum), ланцетные 1.94 см длиной, 0.31 см шириной (E. palustre), широколанцетные (E. adenocaulon), овальные 0.74 см длиной, 0.31 см шириной (E. alpinum) и яйцевидные (E. alsinifolium, E. collinum, E. lactiflorum). Длина листовой пластинки E. lactiflorum 2.47 см, ширина 0.81 см.
У E. alsinifolium и E. palustre в пазухах низовых листьев у основания побега формируются специализированные органы вегетативного размножения – столоны.
С образованием первого генеративного побега растения переходят в генеративное (g) состояние. Особи E. hornemannii характеризуются максимальным развитием надземных и подземных частей. Генеративные побеги достигают 15.3 см (5.0-31.0) высоты. Листья в количестве 3-4 пар, яйцевидной формы, 2.08 см длиной, 0.88 см шириной. Низовые листья отмирают, в их пазухах формируются боковые почки. На верхушке побега формируется простое кистевидное соцветие, несущее (3)5-6 цветков. Преобладают придаточные корни, образующиеся на 1-3 междоузлиях стебля.
Средняя длина побега у других видов колеблется от 1.5 см у E. alpinum, 4.0 см
у E. davuricum, 5.0 см у E. lactiflorum, 6.3 см у E. palustre до 11.5 см у E. alpinum, 11.5 см
у E. davuricum, 17.0 см у E. lactiflorum, 33.0 см у E. palustre. Форма листьев такая же, как
у виргинильных особей, но размеры несколько больше: 2.36 см длина, 0.45 см ширина
(E. palustre); 1.13 см длина, 0.45 см ширина (E. alpinum); 2.45 см длина, 0.81 см ширина
(E. lactiflorum).
Длина корневища от 0.9 до 3.0 см, толщина от 1.8 до 2.7 см у E. collinum, E. davuricum,
E. hornemannii, E. lactiflorum, E. palustre, кроме ползучего у E. alpinum (4.5-7.5 см длиной). На корневище продолжают формироваться почки возобновления, а у видов E. alsinifolium и E. palustre и столоны длиной 6-9 см, имеющие по 7 междоузлий и 3-5 пар плёнчатых листьев. Происходит вегетативное размножение. За счёт образования корневища и столонов особи разрастаются, увеличивается занимаемая ими площадь. В следующий вегетационный период связь с материнским растением прерывается, дочерние особи начинают самостоятельное существование.
4.2.3. Строение монокарпического побега видов рода Epilobium
Почки возобновления у изученных видов рода Epilobium начинают формироваться во второй половине вегетационного сезона. В количестве от 4 до 10 они появляются на корневище, состоящем из нескольких междоузлий (3-4), в пазухах мясистых бурых листьев в конце июля – начале августа. Наиболее хорошо сформированная почка находится у первого междоузлия корневища (в пазухе первой пары отмерших листьев), здесь же у видов E. alsinifolium и E. palustre и появляются столоны. Почка состоит из бесцветных чешуй (1-2), двух листовых зачатков и конуса нарастания. Размеры почки: 0.35-1.5 мм длина и 0.2-0.8 мм ширина.
Столон имеет несколько междоузлий (2-4), 2-3 пары бурых чешуевидных листьев, третья-четвёртая пара листьев – светлая, мясистая (первая пара почки). В почке несколько пар листьев
(3-6) и конус нарастания. Размеры почки: 0.4-2.5 мм длина и 0.4-1.4 мм ширина.
В отличие от Circaea alpina, у видов рода Epilobium длительность развития монокарпических побегов от пробуждения начальной почки до цветения и отмирания составляет 1 вегетационный период, т. е. побеги моноциклического типа. Побеги, образующиеся из почек на концах столонов или из придаточных почек на корневище, сначала проходят внутрипочечное развитие (в этот же вегетационный период), а внепочечное уже в следующем году.
Длиннокорневищный E. alpinum и столонообразующие E. alsinifolium и E. palustre интенсивно расселяются, в то время как короткокорневищные E. adenocaulon, E. collinum,
E. davuricum, E. hornemannii и E. lactiflorum распространяются хуже.
4.2.4.-4.2.5. Возрастной состав и жизненность особей в популяциях видов рода Epilobium
Возрастные спектры популяций видов рода Epilobium изучены нами неравномерно. Наиболее полно исследованы популяции E. hornemannii, E. palustre и E. palustre var. lapponicum – широко распространённые в регионе, в то время как E. davuricum, встречающийся в специфических местообитаниях, значительно меньше. Тем не менее даже по имеющимся данным можно сделать выводы о возрастном составе популяций.
Для изучения жизненного состояния особей в популяциях видов рода Epilobium использовался метод многобалльной оценки. В результате разработаны трёхбалльные шкалы, которые можно в дальнейшем использовать для вычисления различных критериев жизненности всей ценопопуляции.
Результаты обследованных локальных популяций 4 таксонов представлены на рис. 7.
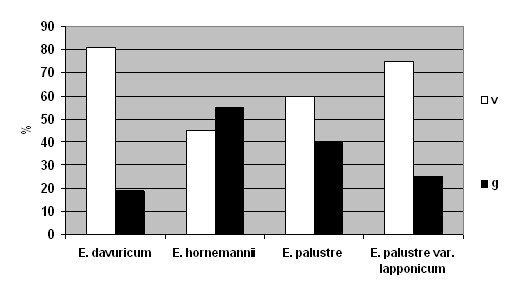
Рис. 7. Возрастные спектры популяций видов рода Epilobium
Сравнивая ценопопуляции разных видов рода Epilobium, можно сделать вывод о том, что возрастные спектры E. davuricum, E. palustre и E. palustre var. lapponicum сходны: в них преобладают особи прегенеративного периода (78-84%). На долю генеративной части приходится 16-22%. Популяции данных видов молодые.
В спектре E. hornemannii генеративные особи составляют больше 50%. Можно сделать вывод, что многие особи этого вида успевают достигнуть зрелости (зацвести и образовать плоды и семена), следовательно семенное размножение у него лучше, чем у других видов. Тем самым
E. hornemannii является более жизнеспособным по сравнению с остальными видами.
Анализируя жизненность вегетативных особей разных видов рода Epilobium, следует отметить, что особи всех видов обладают невысокой жизненностью (1-2-й уровни). Самая высокая жизненность у особей E. hornemannii (2-3), меньше – у E. alpinum (2), E. palustre (2) и
E. palustre var. lapponicum (2), самая низкая жизненность у E. lactiflorum (1).
У генеративных особей жизненность ниже, чем у особей прегенеративного периода. Это, вероятно, можно объяснить тем, что в годы исследований вегетационные периоды были аномально жаркими.
Самая высокая жизненность у особей E. alpinum (3), средняя – у E. davuricum (2) и
E. lactiflorum (2), низкая – у E. hornemannii (1-2), E. palustre (1-2) и E. palustre var. lapponicum (1-2).
4.2.6. Сезонное развитие видов рода Epilobium в Мурманской области
Виды рода Epilobium – летнезелёные растения. Фенологическое развитие продолжается в течение 2-2,5 месяцев. Вегетация начинается после таяния снега в конце мая – начале июня, в некоторые годы в середине июня. Продолжительность созревания семян короче, чем у
Сircaea alpina. Семена созревают ежегодно. В конце августа начинается пожелтение листьев. Отмирание растений происходит в конце августа – начале сентября.
Продолжительность фенологических фаз исследованных видов рода Epilobium идентична. Межфазные периоды короткие.
ВЫВОДЫ
1. Для определения видов рода Epilobium, произрастающих в Мурманской области, предложен дихотомический ключ, который дополняет ключи Р. Н. Шлякова (1953) и
А. К. Скворцова (1980, 1991) и подтверждает диагностические признаки, используемые другими авторами. В основу ключа, как и у других монографов этого рода, положена форма рыльца.
2. Исследованные представители сем. Onagraceae относятся к разным географическим элементам: бореальным (циркумполярные Ch. angustifolium, Circaea alpina, E. palustre, североамериканский E. adenocaulon); гипоарктическим (циркумполярные E. davuricum,
E. hornemannii и европейский E. alsinifolium); плюризональным (европейский E. collinum); арктическим (амфиатлантический E. lactiflorum) и арктоальпийским (евразиатский E. alpinum).
3. В Мурманской области наряду с широко распространёнными видами (Ch. angustifolium, Е. hornemannii и Е. palustre) произрастают и редкие (Circaea alpina, Е. alsinifolium, Е. davuricum и E. lactiflorum), приуроченные к определённым условиям обитания. Эти виды, а также E. alpinum требовательны к целостности окружающей среды и чувствительны к антропогенной нагрузке, встречаются в местах, удалённых от человеческой деятельности.
4. Виды рода Epilobium произрастают во всех районах Мурманской области. Большинство видов (5-6) сконцентрировано в центре, на юго-востоке и на севере. В остальных районах число видов колеблется от 1 до 3.
5. Виды рода Epilobium - растения первичных и вторичных местообитаний как естественного происхождения (ключевые болота, берега рек, ручьёв, нивальные луговины), так и искусственного. Зафиксировано около 100 видов высших сосудистых растений и 25 видов мохообразных, сопутствующих им.
Экологическая амплитуда исследованных видов довольно широкая, изученные виды произрастают на территории Мурманской области в альпийском, субальпийском горных поясах и в лесном поясе. Большинство видов рода Epilobium часто произрастают вместе: широко распространённые (E. hornemannii и E. palustre) и редкие (E. alsinifolium, E. davuricum и
E. lactiflorum).
6. Известно только одно место нахождения Circaea alpina в Мурманской области – окрестности пос. Вуориярви Кандалакшского района. Circaea alpina – мезофит, сциофит, кальцифил, обладает узкой экологической амплитудой, произрастает в тенистых ельниках папоротниково-разнотравно-зеленомошных, прирусловых ивняках. Сопутствующими видами Circaea alpina являются 38 видов сосудистых растений и 25 мохообразных, в том числе виды рода Epilobium (E. hornemannii, E. palustre).
7. Семена Circaea alpina обладают сильным экзогенным и глубоким физиологическим покоем. Опыты по проращиванию семян Circaea alpina в лабораторных условиях не дали положительных результатов. Семена видов рода Epilobium, напротив, имеют высокую всхожесть, разную скорость и длительность прорастания. Высокой всхожестью характеризуются семена
E. hornemannii (до 75%), низкой E. palustre (до 7%). Стратификация низкими температурами отрицательно сказывается на всхожести семян E. hornemannii (32%), напротив всхожесть семян
E. palustre повышалась после воздействия низкими температурами (до 56%).
8. На основании анализа онтогенеза, ритма развития, способов размножения исследованные виды рода Epilobium и Circaea alpina относятся к многолетним травянистым корневищным растениям: большинство является короткокорневищными (E. adenocaulon, E. collinum,
E. davuricum, E. hornemannii, E. lactiflorum), в том числе столонообразующие E. alsinifolium,
E. palustre и Circaea alpina, исключение составляет длиннокорневищный E. alpinum.
9. Длительность развития побегов различна у изученных видов. Circaea alpina формирует монокарпические побеги дициклического типа, виды рода Epilobium моноциклического.
10. Большинство представителей сем. Onagraceae в Мурманской области достаточно хорошо адаптированы к экстремальным условиям существования, размножаются вегетативным и генеративным способом. Семенное размножение играет немаловажную роль в расселении всех исследованных видов рода Epilobium. По способам возобновления изученные виды можно разделить на следующие группы:
- виды, одинаково хорошо размножающиеся как вегетативным способом участками корневища, так и семенами (E. alpinum, E. adenocaulon, E. collinum, E. hornemannii, E. lactiflorum);
- виды, одинаково хорошо размножающиеся как вегетативным способом при помощи специализированных органов (столонов), так и семенами (E. palustre);
- виды, возобновляющиеся преимущественно вегетативно при помощи специализированных органов (столонов) (Circaea alpina, E. alsinifolium);
- виды, возобновляющиеся преимущественно вегетативно участками короткого корневища (E. davuricum).
11. Возрастные спектры изученных популяций Circaea alpina, E. davuricum и E. palustre неполночленные левостороннего типа, в них преобладают особи прегенеративного периода. В отличие от этих видов в спектре E. hornemannii генеративные особи составляют больше 50%, следовательно, этот вид является более жизнеспособным.
12. Жизненное состояние особей в популяциях Circaea alpina и видов рода Epilobium в Мурманской области относительно благополучное. В целом популяции находятся в пределах нормы, преобладают растения средних размеров.
13. Сезонный ритм развития исследованных видов рода Epilobium и Circaea alpina в условиях Мурманской области относительно короткий и длится 2.5 месяца. Вегетация у большинства видов начинается в конце мая – начале июня и заканчивается в конце августа – начале сентября. Исключение составляет Circaea alpina, семена этого вида, находящегося на северном пределе ареала, в некоторые годы не успевают созреть. У видов рода Epilobium период плодоношения короче, чем у Circaea alpina.
СПИСОК ПУБЛИКАЦИЙ ПО ТЕМЕ ДИССЕРТАЦИИ
- Асминг, С.В. Экология и возрастные спектры популяций некоторых видов рода Epilobium L. Кольского полуострова // Кольский полуостров на пороге третьего тысячелетия: проблемы экологии. – Апатиты : Изд. КНЦ РАН, 2003. – С. 84-90.
- Асминг, С.В. Влияние антропогенного фактора и метеорологических условий на жизненность популяции Epilobium davuricum Fisch. ex Hornem. на эвтрофном болоте южного Прихибинья // Материалы школы молодых учёных «Сбалансированное природопользование на примере освоения минеральных ресурсов». – Апатиты : Изд. КНЦ РАН, 2004. – С.138-141.
- Асминг, С.В. Возрастные спектры и экология Circaea alpina L. в Мурманской области //
VII Научная конференция КФ ПетрГУ. – Апатиты, 2004. – С. 35-36. - Асминг, С.В. Материалы к биологии некоторых видов рода Epilobium L., произрастающих в Мурманской области // Материалы докл. II Научной школы молодых специалистов РАН «Комплексность использования минерально-сырьевых ресурсов – основа повышения экологической безопасности региона». – Апатиты : Изд. КНЦ РАН, 2005. – С. 99-103.
- Асминг, С.В. Жизненность ценопопуляций некоторых видов рода Epilobium L., произрастающих в Мурманской области // Материалы междунар. конф. «Структурно-функциональные особенности биосистем Севера». – Петрозаводск : Изд. ПетрГУ, 2005. – С. 27-29.
- Асминг, С.В. Жизненность особей разных возрастных состояний ценопопуляции Circaea alpina L. в Мурманской области // Материалы международного контактного форума по сохранению местообитаний в Баренцевом регионе «International compact forum on habitat conservation in the Barents region». – Сыктывкар, 2005. – С. 13-14.
- Асминг, С.В. Формирование жизненной формы представителей семейства Onagraceae в условиях Крайнего Севера (Мурманская область) // Материалы II Всеросс. науч. конф. «Принципы и способы сохранения биоразнообразия». – Йошкар-Ола, 2006. – С. 27-28.
- Асминг, С.В. Морфолого-биологические особенности и экология видов рода Epilobium L. в Мурманской области // Вест. Оренбургского гос. ун-та. – Оренбург : Изд. ОГУ, 2008 (в печати).