

Петрофитные степи западной части ононской даурии
На правах рукописи
![]()
Головина Екатерина Олеговна
Петрофитные степи западной части Ононской Даурии
03.00.05 – «Ботаника»
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
Санкт-Петербург – 2009
Работа выполнена в Лаборатории географии и картографии растительности Учреждения Российской академии наук Ботанического института им. В. Л. Комарова РАН
Научный руководитель доктор биологических наук,
старший научный сотрудник
Сафронова Ирина Николаевна
Официальные оппоненты: доктор биологических наук,
профессор
Намзалов Бимба-Цырен Батомункуевич
кандидат биологических наук
Паршутина Людмила Петровна
Ведущая организация Московский государственный
университет им. М. В. Ломоносова
Защита состоится 3 июня 2009 г. в 14.00 часов на заседании диссертационного совета Д 002.211.01 при Учреждении Российской академии наук Ботаническом институте им. В. Л. Комарова РАН по адресу: 197376, г. Санкт-Петербург, ул. Профессора Попова, д. 2.
Тел.: (812) 346-47-06, факс (812) 346-36-43
С диссертацией можно ознакомиться в библиотеке Учреждения Российской академии наук Ботанического института им. В. Л. Комарова РАН
Автореферат разослан «……» апреля 2009 г.
Ученый секретарь
![]()
диссертационного совета
кандидат биологических наук О. Ю. Сизоненко
Введение
Актуальность темы. В условиях все увеличивающейся антропогенной нагрузки на биосферу важной предпосылкой ее сохранения служит выявление существующего биологического разнообразия на различных уровнях его организации, в том числе на уровне растительных сообществ (Юрцев, 1992). Актуальность геоботанических исследований возрастает одновременно с увеличением угрозы потери все большего числа компонентов растительного покрова. Сказанное в полной мере относится к Забайкалью и, в частности, к Ононской Даурии, растительность которой, включая и петрофитные степи, исследована все еще недостаточно детально. Изучение состава, структуры, экологии и закономерностей географического распространения петрофитных степей западной части Ононской Даурии позволит более подробно охарактеризовать растительный покров данного региона, что будет способствовать решению проблемы сохранения биоразнообразия.
Цели и задачи исследования. Цель работы – ботанико-географическая характеристика петрофитных степей западной части Ононской Даурии. Для достижения цели были поставлены следующие задачи:
- Выявление разнообразия петрофитных степей.
- Анализ видового и биоморфного состава петрофитно-степных фитоценозов.
- Создание классификационной схемы.
- Экологическая и географическая характеристика петрофитных степей.
Научная новизна работы. Впервые детально охарактеризованы разнообразие и экологическая приуроченность петрофитных степей западной части Ононской Даурии. Проведена их классификация; в результате впервые описаны 8 ассоциаций и 4 субассоциации. Впервые количественно проанализирован состав жизненных форм сосудистых растений петрофитных степей.
Практическая ценность результатов. Результаты работы использованы при инвентаризации растительного мира регионального заказника "Горная Степь" и проектируемой трансграничной (российско-монгольской) особо охраняемой природной территории "Истоки Амура" (Головина, 2006, 2007; Галанин и др., 2007; Беликович, Головина, Сафронова, 2007). Они могут быть применены также при осуществлении мероприятий по региональному экологическому мониторингу степных экосистем, при разработке и оптимизации системы мер по охране биоразнообразия и ценотического фонда Забайкалья и бассейна Амура, для целей крупномасштабного геоботанического картографирования и землеустройства, при преподавании дисциплин биологического и географического циклов в ВУЗах.
Апробация работы. Основные положения диссертации доложены на семинарах Лаборатории географии и картографии растительности Учреждения Российской академии наук Ботанического института им. В. Л. Комарова РАН (Санкт-Петербург, 2006, 2007, 2008), на IV Международном Симпозиуме «Степи Северной Евразии» (Оренбург, 2006), на совместном заседании секции геоботаники Русского Ботанического Общества и Лаборатории географии и картографии растительности (Санкт-Петербург, БИН РАН, 2007). По материалам диссертации опубликовано 8 научных работ, в том числе 1 в журнале из списка, рекомендованного ВАК РФ.
Объем и структура работы. Диссертация изложена на 210 страницах (160 – текст диссертации, 50 – приложения) и состоит из введения, 7 глав, выводов, заключения, списка использованной литературы, включающего 225 наименований (213 – на русском языке, 12 – на иностранных), и 2 приложений. Текст сопровожден 21 таблицей и иллюстрирован 36 рисунками.
Глава 1. Природные условия района исследований
Исследуемая территория находится в Восточном Забайкалье и представляет собой западную левобережную часть бассейна р. Онон, в среднем и нижнем течении (рис. 1). Ее рельеф определяют горные хребты, разделенные долинами рек и межгорными депрессиями. На юго-западе региона преобладают среднегорья высотой до 1400–1600 м (здесь и далее указана высота н. ур. м.), на северо-востоке – низкогорья, приподнятые до 1100 м. Отдельные вершины (г. Сохондо и др.) превышают 2000 м. Хребты сложены в основном сланцами и гранитами. Климат резко континентальный. Среднегодовая температура воздуха от –1.6 до –3.4°С. Годовое количество осадков 300–500 мм: 50–60% суммы выпадает в июле и августе.
В регионе выделяются высотные пояса растительности: лесостепной занимает высоты от 600–700 м до 900–1250 м, лесной простирается до 1650–1900 м, выше лежат подгольцовый и гольцовый пояса. В лесостепном поясе лиственничные, сосновые и березовые леса тяготеют к северным склонам. Степи характерны для вершин сопок и склонов всех экспозиций. На каменистых южных склонах развиты сообщества низкорослых деревьев и кустарников: Armeniaca sibirica, Spiraea aquilegifolia, Ulmus macrocarpa. В нижней полосе лесного пояса господствуют лиственничные леса, здесь же по щебнистым южным склонам встречаются степные фитоценозы. Во всех высотных поясах степи формируются на различных подтипах черноземных почв.
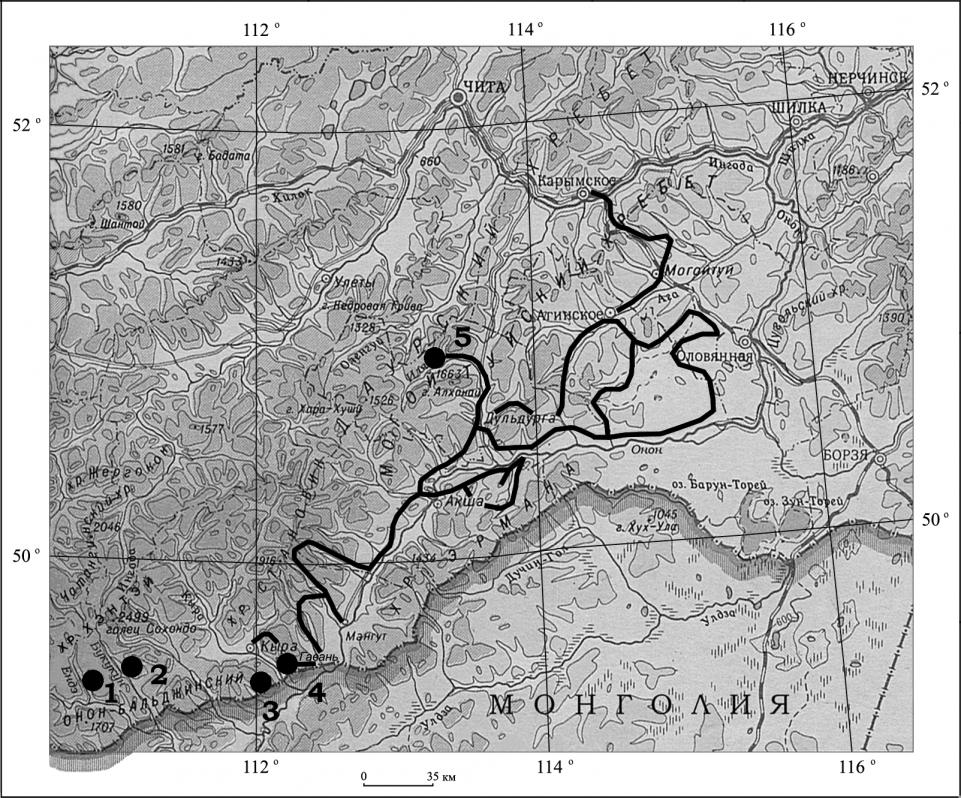
Рис. 1. Карта исследуемой территории.
![]() – маршруты экспедиции;
– маршруты экспедиции; ![]() – места полустационарных исследований: 1 – бассейн р. Ендэ; 2 – бассейн р. Букукун; 3 – региональный заказник «Горная Степь»; 4 – окрестности пос. Гавань Кыринского р-на Читинской обл.; 5 – бассейн р. Иля.
– места полустационарных исследований: 1 – бассейн р. Ендэ; 2 – бассейн р. Букукун; 3 – региональный заказник «Горная Степь»; 4 – окрестности пос. Гавань Кыринского р-на Читинской обл.; 5 – бассейн р. Иля.
Глава 2. История изучения степной растительности Восточного Забайкалья
Первые сведения о флоре и растительности Восточного Забайкалья получены благодаря экспедициям И. Г. Гмелина (1733–1743), П. С. Палласа (1768–1774), Н. С. Турчанинова (1828–1835), Г. И. Радде (1856); дальнейшее изучение растительного покрова региона велось в ходе работ, организованных Переселенческим управлением (Крашенинников, 1910, 1913; Новопокровский, 1912; Сукачев, 1912; Смирнов, 1914). В 1930–е годы растительность Читинской области изучала А. В. Куминова (1938, 1942). Интенсивные исследования степей Восточного Забайкалья начались с конца 1940-х годов, при этом большое внимание уделялось восточным районам территории (Рещиков, 1954; Пешкова 1958, 1963, 1968; Сергиевская, 1964; Алкучанский Говин, 1964; Горшкова, 1966). Ряд работ посвящен степям Восточного Забайкалья в целом (Зарубин, Фролова, 1980; Дулепова, 1993), а также отдельными формациям: нителистниковой (Сергиевская, 1959; Барицкая, 1979; Дулепова, 1981; Зарубин, Ионычева, Терентьева, 1982) и вострецовой (Блюменталь, Кирикова, 1960; Зарубин, 1969). Западная часть региона, петрофитные степи которой описаны в данной работе, оказалась охвачена маршрутами исследователей в значительной мере слабее, нежели восточная (Дулепова, 1983а,б; Зарубин, 1966а,б; Safronova, 2008) Фитоценотическое разнообразие и особенности распространенных здесь степных сообществ до сих пор оставались изучены недостаточно полно.
Согласно схемам ботанико-географического районирования (Лавренко, 1970; Пешкова, 1985; Лавренко и др., 1991) лесостепная часть Восточного Забайкалья, лежит в пределах Центральноазиатской подобласти степной области Евразии. Е. М. Лавренко (Лавренко и др., 1991), включает ее в состав Хангайско-Даурской горнолесостепной провинции, охватывающей территорию от Хангайской горной страны на западе до предгорий хребта Большой Хинган на востоке. Г. А. Пешкова (1966, 1985) обращает внимание на общность растительного покрова степей Восточного Забайкалья, Маньчжурии и северо-восточной Монголии и предлагает объединять горную лесостепь Восточного Забайкалья и территорий, расположенных к востоку, а не к западу от последнего. Согласно этому автору, бльшая часть исследуемого района лежит в пределах Маньчжурско-Даурской лесостепной провинции, простирающейся от горной лесостепи Читинской области до северо-восточного Китая. Мнение о единстве лесостепи Восточного Забайкалья, восточной Монголии и Маньчжурии высказывали и другие авторы (Колесников, 1955; Куренцова, 1962; Камелин, 1994).
Существует и иной подход к ботанико-географическому районированию региона, согласно которому горно-таежные и горно-степные пространства Забайкалья (Даурии) следует объединять в составе единого ботанико-географического выдела (Комаров, 1897; Крылов, 1919; Куваев, 1962; Галанин, Беликович, Галанина, 2008).
Глава 3. Материалы и методы исследования
Сбор полевых материалов проводился в 2005–2007 гг. в составе Даурского ботанического отряда Ботанического сада–института ДВО РАН по программе ДВО РАН «Комплексные экспедиционные исследования природной среды бассейна реки Амур» (рис. 1). Геоботанические описания (всего 314) выполнялись по стандартной методике (Раменский, 1938; Быков, 1957; Понятовская, 1964; Юнатов, 1964). Собрано около 1300 гербарных образцов. Степень щебнистости почв оценивалась глазомерно по трехбалльной шкале исходя из процента покрытия щебнем стенки почвенной прикопки. Для слабощебнистых почв эта величина составляет 10–30%, для среднещебнистых – 30–50%, для сильнощебнистых – 50–70%. Покрытие щебня свыше 70% характерно для грубоскелетных фрагментарных почв (Богатырев, 1954). Подразделение склонов по степени их крутизны принято по стандартной шкале (Почвенная съемка…, 1959).
В камеральных условиях описания заносились в базу данных ИБИС 5.1 (Зверев, 2007). Типизация растительных сообществ выполнена в соответствии с принципами эколого-фитоценотической классификации растительности. При выделении синтаксонов учитывались как доминантные, так и детерминантные виды. Флористическая однородность выделенных синтаксонов проверялась в объеме каждой из детерминантных групп с использованием критерия Кокрена (Василевич, 1985, 1995). Номенклатура сосудистых растений приводится по сводке С. К. Черепанова (1995), в отдельных случаях – в соответствии с «Флорой Центральной Сибири» (1979). Данные о географическом распространении видов приводятся по работам Л. И. Малышева и Г. А. Пешковой (1984), Г. А. Пешковой (2001), а также «Флоре Сибири» (1988–1997). При характеристике экологии видов использованы литературные данные (Горшкова, 1966; Методические указания…, 1974; Горшкова, Зверева, 1982). Принадлежность видов к поясно-зональным группам приведена согласно Л. И. Малышеву и Г. А. Пешковой (1984). Распределение видов сосудистых растений по жизненным формам проводилось на основе системы И. Г. Серебрякова (Серебряков, 1962) с уточнениями по Т. Г. Полозовой (Полозова, 1978, 1981; Кучеров, Милевская, Полозова, 2000). Данные о типах жизненных форм растений взяты из работ А. А. Горшковой (1966), А. А. Горшковой, Г. К. Зверевой (1982), И. В. Борисовой, Т. А. Поповой (1984).
Глава 4. Флористический состав петрофитных степей западной части
Ононской Даурии
4. 1. Таксономический состав флоры
По данным геоботанических описаний флора петрофитных степей западной части Ононской Даурии включает 210 видов, относящихся к 119 родам и 44 семействам.
В число многовидовых семейств входят Asteraceae, Rosaceae, Poaceae и Fabaceae (табл. 1). Существенная роль последнего, а также Brassicaceae, Liliaceae s. l., Apiaceae, Lamiaceae указывает на аридный характер флоры и отражает ее древнесредиземные связи (Малышев, 1972; Пешкова, 1972; Камелин, 1973). С другой стороны, высокий ранг семейств Rosaceae, Ranunculaceae, Cyperaceae типичен для бореальных флор (Толмачев, 1974). По-видимому, подобное сочетание аридных и бореальных черт закономерно для флор степных сообществ в пределах лесостепи (Банникова, 1986; Кыргыс, 2006).
Наибольшее число видов насчитывают роды Artemisia, Potentilla, Oxytropis, Astragalus и Allium, что типично для степных флор Байкальской Сибири и Южно-Сибирской горной области (Пешкова, 1972, 2001; Малышев, Пешкова, 1984; Банникова, 1986). Ведущая позиция этих родов связана с положением исследуемого региона в пределах либо вблизи от области крупных автохтонных центров их развития (Крашенинников, 1946; Попов, 1956; Положий, 1964; Фризен, 1988).
Таблица 1.
Семейства флоры петрофитных степей западной части Ононской Даурии,
преобладающие по числу видов
| Семейство | Число видов |
| Asteraceae | 36 |
| Rosaceae | 22 |
| Fabaceae | 19 |
| Poaceae | 19 |
| Brassicaceae | 9 |
| Caryophyllaceae | 9 |
| Ranunculaceae | 9 |
| Liliaceae s. l. | 9 |
| Apiaceae | 7 |
| Lamiaceae | 6 |
| Scrophulariaceae | 6 |
| Cyperaceae | 6 |
Максимальное фитоценотическое значение имеют представители семейств Asteraceae, Poaceae и Rosaceae. Из числа сложноцветных к доминантам и субдоминантам относятся Filifolium sibiricum, Artemisia monostachya, Arctogeron gramineum, из злаков – Festuca spp., Koeleria cristata, Stipa baicalensis, из розоцветных – Chamaerhodos trifida и виды Potentilla.
4. 2. Экологический состав флоры
По отношению к фактору увлажнения большинство растений флоры является мезоксеро- и ксеромезофитами, что соответствует положению изучаемых петрофитных степей в пределах лесостепного и нижней части лесного поясов. Именно к этой экологической группе относятся многие доминанты и субдоминанты (Filifolium sibiricum, Lespedeza juncea и др.). Мезофиты и ксерофиты принимают меньшее, при этом почти равное, участие в формировании видового состава флоры, но фитоценотическая роль ксерофитов заметно выше, что обусловлено приуроченностью многих петрофитно-степных сообществ к сухим крутым южным склонам.
По отношению к фактору богатства почв подавляющее большинство растений флоры является гликофитами. Это согласуется с тем, что исследуемые нами степи приурочены в основном к бескарбонатным черноземам, развитым на элюво-делювии горных пород, бедных основаниями.
4. 3. Поясно-зональный состав флоры
Основу флоры составляют горно-степные, лесостепные и собственно степные виды, на долю которых приходится около 90% всего видового состава. В спектрах поясно-зональных групп, составленных для каждого типа сообществ с учетом встречаемости видов, лидируют горно-степные растения, хорошо приспособленные к выживанию на щебнистых субстратах (Пешкова, 2001). Максимальных значений (свыше 70%) их доля достигает в наиболее ксерофитных типах степей, приуроченных к наименее мелкоземистым почвам, и отличающихся низким разнообразием представителей всех прочих поясно-зональных групп. Из числа последних заметную роль здесь играют лишь собственно степные растения, что обусловлено присутствием высоко константных видов с широкой экологической амплитудой: Koeleria cristata, Bupleurum scorzonerifolium, Rhaponticum uniflorum. Доля лесостепных растений возрастает по мере увеличения мелкоземистости и особенно увлажнения почв. В спектрах большинства типов сообществ они занимают 2-е либо 3-е место, в последнем случае лишь ненамного уступая собственно степным. Максимальную роль лесостепные виды играют в наиболее мезофитных фитоценозах, приуроченных к слабощебнистым почвам, например, в мятликово-разнотравных и стоповидноосоково-байкальскоковыльных, где отмечены растения, несвойственные прочим типам сообществ: Artemisia sericea, Vicia amoena и др. Доли представителей светлохвойно-лесной и монтанной групп обычно составляют 1–3%. Лишь в мятликово-разнотравных степях, занимающих небольшие участки на преимущественно облесенных склонах в лесном поясе, доля первых увеличивается до 10% за счет повышенной встречаемости Artemisia tanacetifolia, Spiraea media, Vicia unijuga и др.
Глава 5. Биоморфная структура петрофитных степей западной части
Ононской Даурии
Анализ биоморфной структуры петрофитных степей проведен двумя способами: для всех типов сообществ составлены спектры жизненных форм, «взвешенные» с учетом проективного покрытия видов либо с учетом их встречаемости (Лакин, 1980). При сопоставлении спектров жизненных форм, «взвешенных» по проективному покрытию видов, сравнение проведено только для эдификаторного травяного яруса.
5. 1. Анализ спектров жизненных форм, «взвешенных» с учетом
проективного покрытия растений
Ведущая роль в петрофитных степях западной части Ононской Даурии принадлежит стержнекорневым поликарпическим травам (рис. 2). Наибольшего обилия достигают многоглаво-каудексовые виды с рыхлой формой роста побегов (Filifolium sibiricum, Potentilla tanacetifolia и др.). Их доля обычно составляет 50–60%; они лидируют в спектрах большинства типов сообществ. Доля стержнекорневых подушковидных растений (Chamaerhodos trifida и др.) редко превышает 2–4%. Эта биоморфа наиболее конкурентоспособна на грубоскелетных фрагментарных почвах вершин сопок, где возрастает скорость ветра, и нередко наблюдаются усиленный прогрев, а также значительное иссушение приземного слоя воздуха и почвы. В развитых здесь хамеродосовых и горноколосниковых степях доля стержнекорневых подушковидных трав достигает 30–45%; данная биоморфа выходит на 1-е либо 2-е место в спектре. Участие одноглаво- и малоглаво-каудексовых и длиннокорневищно-стержнекорневых видов обычно невелико.
Плотнодерновинные травы (Festuca spp., Carex pediformis и др.) занимают, как правило, второе место в спектрах, лидируя лишь в байкальскоковыльных степях.
Стержнекорневые и плотнодерновинные растения являются основными компонентами, формирующими большинство типов петрофитных степей региона. При этом плотнодерновинные травы менее конкурентоспособны: резкие колебания влажности поверхностных горизонтов щебнистых почв затрудняют развитие их интенсивной корневой системы. Увлажнение нижних горизонтов щебнистых почв близко к стабильному, что обуславливает высокую конкурентную мощность стержнекорневых форм, способных использовать влагу из этих горизонтов (Вальтер, 1968; Юрцев, 1978). Кроме того, стержневой корень, в большей степени, нежели мочковатая корневая система, позволяет растениям закрепиться на подвижном субстрате и противостоять погребению щебнем на крутых склонах (Юрцев, 1978). В то же время в более засушливых районах Восточного Забайкалья отмечены дерновиннозлаковые петрофитные степи с господством ксерофильных типчаков, лучше приспособленных к перенесению недостатка влаги, чем доминирующее в изучаемых сообществах мезоксерофильное стержнекорневое разнотравье (Горшкова, 1971). Вероятно, стержнекорневые виды в Восточном Забайкалье получают конкурентное преимущество на щебнистых субстратах лишь в условиях достаточного увлажнения.
Обилие рыхлодерновинных трав возрастает в наиболее мезофитных типах степей на слабощебнистых почвах обычно за счет Poa botryoides. Исключение составляют байкальскоковыльные степи, где данная биоморфа, по-видимому, не выдерживает конкуренции с плотнодерновинной. Роль рыхлодерновинных форм повышается также в сообществах на каменистых эродированных склонах, где типичны Agropyron cristatum и Cleistogenes kitagawae.
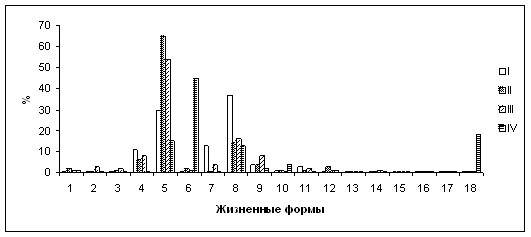
Рис. 2. Доли представителей различных жизненных форм в покрытии травостоя петрофитных степей.
Типы степей: I – стоповидноосоково-байкальскоковыльные; II – разнотравно-нителистниковые; III – разнотравно-спирейно-нителистниковые; IV – горноколосниково-хамеродосовые.
Жизненные формы: 1 – кустарнички простратные; 2 – полукустарники; 3 – полукустарнички; поликарпические травы: 4 – стержнекорневые одноглаво- и малоглаво-каудексовые; 5 – стержнекорневые многоглаво-каудексовые с рыхлой формой роста побегов; 6 – стержнекорневые многоглаво-каудексовые подушковидные; 7 – длиннокорневищно-стержнекорневые; 8 – плотнодерновинные; 9 – рыхлодерновинные; 10 – длиннокорневищные; 11 – короткокорневищные; 12 – луковичные и корневищно-луковичные; 13 – корнеклубневые; 14 – корнеотпрысковые; 15 – столонообразующие; монокарпические травы: 16 – однолетние; 17 – двулетние; 18 – многолетние.
Характерная черта петрофитных степей региона – малая фитоценотическая роль короткокорневищных трав, доля которых не превышает 3%, что обусловлено, прежде всего, сухостью верхних горизонтов щебнистых почв. Фитоценотическое значение длиннокорневищных и корневищнолуковичных видов столь же невелико.
Из числа монокарпических растений наибольшего обилия достигает многолетний столонообразующий суккулент Orostachys spinosa, высоко толерантный к колебаниям приземной температуры и почвенной влажности (Дулепова, 1993). Однако, являясь конкурентно слабым видом (Бялт, 1999), он «увеличивает» свою численность лишь в условиях, наименее благоприятных для растений прочих биоморф: на сухих вершинах сопок с грубоскелетными фрагментарными почвами. В спектре горноколосниковых степей, где O. spinosa находится вблизи от своего эколого-ценотического оптимума, многолетние монокарпики занимают первое место, в спектре хамеродосовых – второе.
Доля полукустарничков достигает 20% в холоднополынно-разнотравных степях, занимающих склоны, подверженные сильной гравитационной эрозии. Здесь обильна Artemisia frigida, которая, благодаря укоренению полегающих вегетативных побегов, высоко конкурентоспособна в этих условиях. Доля кустарничков составляет 15% в одноколосковополынных степях, развитых на крутых песчаных склонах, где сильно разрастается вегетативно-подвижный Thymus dahuricus, способный выдерживать погребение песком своих плагиотропных побегов. В спектрах остальных синтаксонов «вес» как полукустарничков, так и кустарничков обычно не превышает 3%.
5. 2. Анализ спектров жизненных форм, «взвешенных» с учетом
встречаемости растений
«Взвешенные» с учетом встречаемости видов, биоморфные спектры всех типов петрофитных степей региона сходны, что свидетельствует о единстве их генезиса и об их длительном существовании в условиях среды, близких к современным.
Во всех без исключения спектрах лидируют стержнекорневые поликарпические растения, доля которых составляет 45–60%. Роль короткокорневищных поликарпиков возрастает с увеличением увлажнения почв и содержания в них мелкозема. В большинстве типов сообществ доля этих видов составляет около 10%; в спектрах они, как правило, занимают второе место. Из числа прочих биоморф заметную роль играют плотно- и рыхлодерновинные, а также длиннокорневищные травы, доли которых колеблются от 3 до 11%, чаще в пределах 5–7%.
Биоморфные спектры исследуемых нами степей, «взвешенные» с учетом встречаемости видов, в общих чертах сходны с аналогичными спектрами степных флороценотипов Южной Сибири (Намзалов, 1994). По соотношению встречаемости различных биоморф петрофитные степи западной части Ононской Даурии наиболее близки к лугово-степному флороценотипу, хотя и отличаются от него намного большей ролью стержнекорневых форм. По-видимому, это обусловлено тем, что лугово-степной флороценотип, выделенный Б. Б. Намзаловым, включает, в том числе и плакорные варианты степных сообществ, для которых, по сравнению с петрофитными, типично повышенное разнообразие короткокорневищных видов и пониженное – стержнекорневых.
Глава 6. Единицы классификации и классификационная схема петрофитных степей западной части Ононской Даурии
К вопросу классификации степей Восточного Забайкалья обращались многие исследователи (Рещиков, 1954; Пешкова, 1985; Барицкая, 1979; Зарубин, Фролова, 1980; Арбузова, Дулепова, 1989; Дулепова, 1993; Королюк, 2002). Р. В. Камелиным (1987) на основе генетического подхода к классификации создана система флороценотипов растительности Монголии, в которой нашли свое место растительные сообщества, распространенные, в том числе и на исследуемой территории.
В литературе нет единого мнения о положении различных типов степей Восточного Забайкалья, прежде всего, нителистниковых и байкальскоковыльных в системах высших синтаксонов растительности. В рамках эколого-физиономического подхода разные авторы (Лавренко 1940, 1956; Юнатов, 1950; Рещиков, 1954, 1961; Пешкова, 1972, 1985 и др.) относят их к различным подтипам степного типа растительности. По мнению Р. В. Камелина (1987), реконструируемое флороценогенетическое родство нителистниковых и байкальскоковыльных сообществ отличает их от степей, что дает основание данному автору включать их в флороценотип восточноазиатских прерий.
Разработанная нами классификация петрофитных степей западной части Ононской Даурии (табл. 2) является эколого-фитоценотической доминантно-детерминантной (Василевич, 1985, 1995). Она дополняет новыми данными классификационные схемы упомянутых выше исследователей. Классификация имеет иерархическую структуру, в качестве ее единиц выбраны тип растительности, формация, ассоциация и субассоциация. Следуя Е. М. Лавренко (1940, 1956) и другим авторам мы рассматриваем исследуемые фитоценозы в составе степного типа растительности. Под названием петрофитные степи мы объединяем степные сообщества, формирующиеся на щебнистых маломощных почвах (Лавренко, 1940; Карамышева, 1960; Исаченко, Рачковская, 1961; Растительный покров Хакасии, 1976; Королюк, Макунина, 1998). Формация объединяет растительные сообщества с одинаковым видовым набором доминантов господствующего яруса; при выделении единиц данного ранга нами учитывалась фитоценотическая замещаемость видов. К одной ассоциации отнесены сообщества с общим видовым составом доминантов и детерминантов, а также сходной структурой. Субассоциации выделяются по особенностям флористического состава, а также по различиям в соотношениях субдоминантов и строении фитоценозов.
Таблица 2.
Схема классификации петрофитных степей западной части Ононской Даурии
| Формация | Ассоциация | Субассоциация |
| Filifolieta sibiricae |
| |
|
| |
| ||
| ||
| ||
| ||
|
Таблица 2 (продолжение).
| Формация | Ассоциация | Субассоциация |
| Filifolieta sibiricae |
| |
| ||
| ||
| ||
| ||
| Artemisieta monostachyae |
| |
| Pulsatilleto turczaninovii-Potentilleta acervatae |
| |
| ||
| Chamaerhodeta trifidae |
| |
| Orostachyeta spinosae |
| |
| Stipeta baicalensis |
|
Глава 7. Ботанико-географическая характеристика петрофитных степей
западной части Ононской Даурии
7. 1. Нителистниковые степи (Filifolieta sibiricae)
Нителистниковые степи играют большую роль в растительном покрове лесостепи Восточного Забайкалья, северо-восточной Монголии и северо-восточного Китая (Пешкова, 1966; Лавренко и др., 1991). В целом ареал формации простирается от Приангарья на западе до лесостепных островов Дальнего Востока (Пешкова, 1958; Ярошенко, 1958; Куренцова, 1962).
Среди всех описываемых типов петрофитных степей нителистниковые отличаются наибольшим разнообразием и максимальным спектром осваиваемых местообитаний. Они нередко доминируют в ландшафтах лесостепного пояса, но встречаются и в лесном, где строго приурочены к наиболее крутым щебнистым участкам южных склонов, обращенных в долины рек. Для каждого пояса характерен свой набор типов нителистниковых степей; не отмечено ни одного синтаксона, сообщества которого были бы распространены и в лесостепном, и в лесном поясах.
Нителистниковые степи лесостепного пояса
Отличительная черта нителистниковых степей лесостепного пояса – участие ряда видов (Scutellaria baicalensis, Dasiphora parvifolia, Festuca lenensis, Arctogeron gramineum, Potentilla leucophylla, Stipa baicalensis, Allium bidentatum, Adenophora gmelinii, Polygala tenuifolia, Cymbaria daurica и др.), не встречающихся, либо очень редких в аналогичных сообществах лесного пояса. Средняя высота травостоя 20–40 см, его общее проективное покрытие 30–60%. В травяном ярусе фитоценозов, развитых на сильнощебнистых почвах, выделяются два хорошо обособленных подъяруса; с увеличением содержания мелкозема в почве вертикальная дифференциация травостоя становится менее четкой. Характерно участие кустарника Dasiphora parvifolia. Из злаков наибольшую роль играют Festuca litvinovii и F. lenensis. Выделено 5 ассоциаций.
Сообщества асс. Herbo variae-Festuco litvinovii-Filifolietum sibiricae распространены в северо-восточной низкогорной части региона в диапазоне высот от 650 до 1000 м. Занимают в основном вершины сопок и верхние части склонов различной крутизны и экспозиции. Почвы сильнощебнистые, развитые на сланцах. Типично заметное участие петроксерофитов Pulsatilla tenuiloba, Arctogeron gramineum, Eremogone capillaris, а также петрофильного суккулента Orostachys spinosa, чьи розетки образуют небольшие скопления в наиболее щебнистых участках, тем самым обуславливая мозаичность фитоценозов. Из злаков наибольшее фитоценотическое значение имеют Festuca litvinovii, Koeleria cristata и Poa argunensis. К группе детерминантных видов, помимо P. argunensis и Orostachys spinosa, относятся также петроксерофильные горно-степные Potentilla sergievskajae и Crepis crocea (табл. 3). Нителистниковые степи с согосподством Festuca litvinovii до настоящего времени отмечены лишь в Восточном Забайкалье (Дулепова, 1993).
Сообщества асс. Herbo variae-Artemisio monostachyae-Filifolietum sibiricae nov. распространены в юго-западной среднегорной части региона на высоте от 1050 до 1250 м; приуроченность к элементам рельефа и почвам – как у предыдущей ассоциации. По сравнению с последней, разнотравно-одноколосковополынно-нителистниковые степи развиты в более влажных условиях, что связано с большей абсолютной высотой местности. В фитоценозах обилен петро- и псаммофильный мезоксерофит Artemisia monostachya, обычны петромезоксерофиты Phlojodicarpus sibiricus и Festuca sibirica. Кроме упомянутых видов, детерминантами ассоциации являются также петромезоксерофильные Potentilla verticillaris и Erysimum flavum. По различиям в видовом составе, а также в соотношениях субдоминантов и структуре сообществ, обусловленных неодинаковыми условиями локального увлажнения и различным содержанием почвенного мелкозема, в составе ассоциации выделены 4 субассоциации.
Таблица 3.
Фитоценотическая характеристика нителистниковых степей (фрагмент)
| Номера синтаксонов | 1 | 2.1 | 2.2 | 2.3 | 3 | 4 | 5 | |||||||
| Число описаний | 27 | 35 | 36 | 14 | 19 | 17 | 13 | |||||||
| Общее проективное покрытие, % | 50-60 | 50-55 | 50-55 | 45-50 | 50 | 55-60 | 50-65 | |||||||
| Общее число видов в ассоциации | 106 | 137 | 119 | 110 | 82 | |||||||||
| Среднее число видов на 100 м | 30 | 37 | 45 | 40 | 45 | 40 | 35 | |||||||
| Filifolium sibiricum | 9 | 100 | 10 | 100 | 9 | 100 | 10 | 100 | 9 | 100 | 25 | 100 | 15 | 100 |
| Polygonum angustifolium | 3 | 96 | 3 | 97 | 2 | 100 | 2 | 100 | 1 | 84 | 1 | 71 | + | 69 |
| Bupleurum scorzonerifolium | + | 70 | 1 | 94 | 2 | 97 | 3 | 86 | 3 | 95 | 3 | 94 | 2 | 92 |
| Koeleria cristata | 5 | 96 | 2 | 89 | 1 | 75 | 1 | 79 | + | 53 | 1 | 65 | 6 | 100 |
| Orostachys spinosa | 3 | 67 | + | 3 | + | 3 | + | 7 | + | 5 | ||||
| Poa argunensis | 1 | 85 | + | 51 | 1 | 47 | + | 36 | + | 26 | + | 35 | ||
| Potentilla sergievskajae | + | 19 | + | 7 | + | 5 | + | 12 | ||||||
| Crepis crocea | + | 26 | + | 7 | + | 12 | ||||||||
| Artemisia monostachya | + | 11 | 8 | 94 | 4 | 89 | 4 | 79 | + | 11 | + | 24 | 3 | 31 |
| Phlojodicarpus sibiricus | + | 22 | 3 | 89 | 3 | 100 | 3 | 79 | + | 16 | + | 35 | ||
| Festuca sibirica | + | 15 | 1 | 74 | 2 | 78 | 2 | 79 | 1 | 26 | + | 41 | ||
| Potentilla verticillaris | + | 37 | + | 57 | + | 50 | + | 50 | + | 32 | + | 47 | + | 15 |
| Erysimum flavum | + | 15 | + | 23 | + | 28 | + | 29 | + | 16 | ||||
| Thalictrum appendiculatum | + | 4 | + | 7 | + | 47 | + | 11 | + | 24 | + | 8 | ||
| Lupinaster pentaphyllus | + | 3 | + | 58 | + | 14 | + | 5 | ||||||
| Polygonum alopecuroides | + | 9 | + | 75 | + | 14 | ||||||||
| Potentilla acervata | + | 30 | + | 14 | + | 64 | + | 14 | + | 21 | + | 35 | 5 | 85 |
| Sanguisorba officinalis | + | 26 | + | 97 | + | 21 | + | 11 | + | 12 | + | 92 | ||
| Carex pediformis | + | 7 | + | 11 | 3 | 56 | + | 43 | 3 | 84 | + | 12 | + | 77 |
| Galium verum | + | 19 | + | 20 | 1 | 36 | 1 | 43 | 2 | 79 | + | 12 | + | 92 |
| Potentilla tanacetifolia | + | 37 | + | 54 | + | 28 | 2 | 79 | 2 | 89 | + | 24 | + | 8 |
| Patrinia rupestris | + | 7 | + | 9 | + | 6 | 1 | 71 | 1 | 89 | + | 38 | ||
| Vicia popovii | + | 7 | + | 17 | + | 28 | + | 71 | + | 63 | ||||
| Cleistogenes kitagawae | + | 9 | + | 57 | + | 53 | + | 47 | + | 8 | ||||
| Lespedeza juncea | 1 | 56 | + | 51 | + | 28 | 3 | 100 | 1 | 68 | 2 | 88 | ||
| Leontopodium leontopodioides | + | 26 | + | 20 | + | 19 | + | 29 | + | 63 | 1 | 82 | ||
| Spiraea pubescens | + | 7 | + | 3 | + | 19 | + | 21 | 5 | 63 | + | 6 | ||
| Clematis hexapetala | + | 7 | 1 | 63 | + | 18 | ||||||||
| Saussurea salicifolia | + | 67 | + | 17 | + | 11 | + | 57 | 1 | 84 | + | 6 | + | 15 |
| Lespedeza davurica | + | 11 | 1 | 36 | 1 | 63 | + | 12 | ||||||
| Spiraea aquilegifolia | + | 11 | + | 3 | + | 7 | 1 | 68 | + | 35 | ||||
| Artemisia gmelinii | + | 22 | + | 14 | 1 | 53 | + | 18 | ||||||
Примечание. Синтаксоны: 1 – асс. Herbo variae-Festuco litvinovii-Filifolietum sibiricae; 2 – асс. Herbo variae-Artemisio monostachyae-Filifolietum sibiricae: 2.1 – субасс. festucetosum lenensis, 2.2 – субасс. caricetosum pediformis, 2.3 – субасс. lespedezetosum junceae; 3 – асс. Herbo variae-Spiraeo pubescentis-Filifolietum sibiricae; 4 – асс. Herbo variae-Scutellario baicalensis-Filifolietum sibiricae; 5 – асс. Herbo variae-Artemisio macilentae -Filifolietum sibiricae.
В столбце слева приводится среднее проективное покрытие (в %), справа – постоянство (в %). «+» – среднее проективное покрытие вида менее 1%. В рамках – группы детерминантных видов.
Сообщества субасс. arctogeronetosum graminei nov. характерны для выпуклых вершин и взлобков на южных склонах. В них обильны петроксерофиты Arctogeron gramineum, Pulsatilla tenuiloba, Eremogone capillaris. Собственных детерминантных видов у этой субассоциации нет.
Фитоценозы субасс. festucetosum lenensis nov. приурочены к плоским либо западинным участкам вершин сопок. Здесь заметна роль Festuca lenensis и петрофильного Allium burjaticum. Собственных детерминантных видов данный синтаксон также не имеет, от субасс. arctogeronetosum graminei и lespedezetosum junceae он отличается видами, общими со следующей субассоциацией. В основном это горно-степные петромезоксерофиты: Allium burjaticum, Oxytropis mixotriche и др.
Степи субасс. caricetosum pediformis nov. развиты в верхних частях северных склонов. Их отличительной чертой является участие мезофильных и ксеромезофильных лугово-степных видов: Lupinaster pentaphyllus, Polygonum alopecuroides, Potentilla acervata, Carex pediformis, Adenophora tricuspidata и др.
Сообщества субассоциации lespedezetosum junceae nov. встречаются редко небольшими участками на каменистых западных и южных склонах с выходами горных пород. Для них характерен сильно разреженный кустарниковый ярус из Dasiphora parvifolia, Armeniaca sibirica и Spiraea pubescens. В травяном ярусе повышена роль Lespedeza juncea, Potentilla tanacetifolia, Patrinia rupestris. В качестве детерминантных выступают виды, тяготеющие к мелкоземистым почвам (Lespedeza juncea, Leontopodium leontopodioides, Saposhnikovia divaricata, Astragalus tenuis и др.) либо к каменистым эродированным участкам склонов (Patrinia rupestris, Potentilla tanacetifolia, Cleistogenes kitagawae, Vicia popovii).
Сообщества асс. Herbo variae-Spiraeo pubescentis-Filifolietum sibiricae nov. распространены по всей исследуемой территории на высоте 700–1200 м. Они характерны для умеренно крутых каменистых южных склонов со среднещебнистыми почвами и выходами сланцев. Разнотравно-спирейно-нителистниковые степи могут занимать протяженные участки, особенно в средней трети склонов. Типичен разреженный кустарниковый ярус высотой до 1.5 м, образованный Spiraea pubescens, S. aquilegifolia, Armeniaca sibirica и Dasiphora parvifolia. В травяном ярусе заметна роль петро- и эрозиофилов Potentilla tanacetifolia, Patrinia rupestris и других представителей ксеромезофильного разнотравья (Galium verum, Pulsatilla turzcaninovii), а также Carex pediformis и Stipa sibirica s. l. Возрастает встречаемость одно- и двулетников: Dontostemon integrifolius, Chamaerhodos erecta, Amethystea caerulea. Большинство детерминантных видов ассоциации (Spiraea pubescens, S. aquilegifolia, Artemisia gmelinii, Lespedeza davurica) более характерны для сообществ низкорослых деревьев и кустарников (Камелин, 1994), нежели для собственно степей.
Сообщества асс. Herbo variae-Scutellario baicalensis-Filifolietum sibiricae широко распространены в регионе на высоте от 700 до 1100 м; часто занимают значительные площади, определяя облик ландшафта. Приурочены в основном к пологим и покатым склонам, но встречаются также на вершинах и подгорных шлейфах. Почвы среднещебнистые, развитые на сланцах. В фитоценозах обильны Scutellaria baicalensis, Bupleurum scorzonerifolium, Lespedeza juncea, Stellera chamaejasme, Artemisia macilenta. Виды разнотравья либо принимают равное участие в сложении травостоя, либо преобладают лишь 2–3 из них. Из злаков, кроме широко распространенных типчаков, существенна роль Stipa sibirica s. l. и особенно S. baicalensis. Дифференциация от других ассоциаций негативна. Нителистниковые степи, близкие по видовому составу к рассматриваемым сообществам, характерны для Западного и Восточного Забайкалья, а также Восточной Монголии (Сергиевская, 1959; Рещиков, 1961; Дашням, 1966; Дулепова, 1993).
Сообщества асс. Herbo variae-Koelerio cristatae-Filifolietum sibiricae встречены на северо-западном макросклоне хр. Могойтуйского, где, по сравнению с прочими обследованными участками лесостепного пояса, формируются условия повышенного атмосферного увлажнения (Атлас Забайкалья, 1967). Они развиты на высоте 900–950 м в верхней половине склонов на песчано-дресвяном элюво-делювии гранитов. По обилию Koeleria cristata, Potentilla acervata, Galium verum, Pulsatilla turzcaninovii, Carex pediformis близки к нителистниковым степям лесного пояса. В то же время для сообществ типичны широко распространенные в лесостепном поясе Scutellaria baicalensis, Arctogeron gramineum, Potentilla leucophylla, Pulsatilla tenuiloba, Dasiphora parvifolia. Собственных детерминантных видов у ассоциации нет. Нителистниковые сообщества с согосподством Koeleria cristata встречаются в Восточном Забайкалье, в восточной части Монголии и в Приангарье (Сергиевская, 1959; Дашням, 1966; Барицкая, 1979; Номоконов и др., 1984).
Нителистниковые степи лесного пояса
Травостой нителистниковых степей лесного пояса более сомкнутый: его общее проективное покрытие обычно составляет 60–75%. Для всех типов фитоценозов характерен сильно разреженный подъярус высотой 80–100 см, образованный побегами Stipa sibirica s. l. Доминирующие растения принадлежат ко 2-му подъярусу высотой 30–45 см; роль низкорослых видов приземного 3-го подъяруса высотой 7–20 см чаще всего невелика. В большинстве сообществ обильны лугово-степные Potentilla acervata, Galium verum, Carex pediformis, лесная Sanguisorba officinalis. Из злаков наибольшую роль играют Koeleria cristata, Poa botryoides и Stipa sibirica s. l. Участие кустарников незначительно.
Выделено 4 ассоциации нителистниковых степей лесного пояса, сообщества которых строго приурочены к наиболее крутым участкам южных склонов, обращенным в долины рек. Взятые вместе, эти ассоциации намного четче дифференцируются по отсутствию видов, обычных в степях лесостепного пояса (Scutellaria baicalensis и др.), нежели собственной малочисленной и экологически разнородной детерминантной группой (Thalictrum petaloideum, Chamaerhodos erecta, Oxytropis myriophylla).
Сообщества асс. Herbo variae-Potentillo acervatae-Filifolietum sibiricae встречены в бассейне р. Букукун на высоте 1180–1250 м. Они занимают небольшие взлобки в нижней трети преимущественно облесенных склонов. Почвы среднещебнистые, развитые на сланцах. Существенное участие в сложении фитоценозов принимает ксеромезофильное разнотравье: Pulsatilla turczaninovii, Schizonepeta multifida, Allium senescens. Из злаков наиболее обильны Koeleria cristata и Poa botryoides. Детерминантами являются ксеромезофильные Delphinium grandiflorum, Artemisia desertorum, Potentilla matsoukana subsp. crebridens и более ксерофильные Papaver rubro-aurantiacum и Carex duriuscula. Нителистниковые степи с обилием Potentilla acervata ранее в том же районе были отмечены Б. И. Дулеповой (1981). Сведений о распространении сообществ данного типа в других районах Восточного Забайкалья, а также за его пределами, нам найти не удалось.
Сообщества асс. Herbo variae-Pulsatillo turczaninovii-Filifolietum sibiricae отмечены в бассейне р. Иля на высоте 900–950 м. Характерны для покатых и крутых склонов с выходами гранитов. Почвы среднещебнистые. Типична повышенная роль растений, тяготеющих к каменистым эродированным склонам: Potentilla tanacetifolia, Patrinia rupestris, Vicia popovii, Cleistogenes kitagawae. От прочих ассоциаций лесного пояса разнотравно-прострелово-нителистниковая дифференцируется негативно. Нителистниковые степи с согосподством Pulsatilla turczaninovii обычны как в Восточном, так и в Западном Забайкалье (Сергиевская, 1951, 1959).
Сообщества асс. Herbo variae-Lespedezo junceae-Filifolietum sibiricae описаны в бассейне р. Букукун на высоте 1150–1300 м. Характерны для протяженных крутых склонов, обычно полностью покрытых степью. Почвы слабощебнистые, развитые на сланцах. Среди прочих типов нителистниковых степей выделяются наиболее сомкнутым травостоем, в составе которого обильны петро- и эрозиофилы Patrinia rupestris, Potentilla tanacetifolia и Artemisia gmelinii. Характерная особенность – участие Cleistogenes kitagawae и Agropyron cristatum. От других ассоциаций нителистниковых степей лесного пояса отличается высокой встречаемостью Lespedeza juncea, Leontopodium leontopodioides, Spiraea aquilegifolia и Artemisia gmelinii. Нителистниковые степи, с согосподстовом Lespedeza juncea встречаются в Западном и Восточном Забайкалье, в том числе и в пределах лесостепного пояса (Барицкая, 1979; Дулепова, 1981, 1993). Вероятно, рассматриваемая ассоциация на протяжении своего ареала представлена рядом замещающих вариантов.
Сообщества асс. Herbo variae-Artemisio macilentae-Filifolietum sibiricae nov. встречены в бассейне р. Букукун на высоте 1200–1250 м, где развиты на песчано-дресвяном элюво-делювии гранитов в нижней половине покатых и умеренно крутых склонов. В фитоценозах заметную роль играет стелющийся кустарничек Thymus dahuricus, который нередко сильно разрастается и образует куртины до 1 м в диаметре. В отличие от прочих типов нителистниковых степей лесного пояса, здесь изредка встречаются Pulsatilla tenuiloba и Allium burjaticum, широко распространенные в лесостепном поясе и редкие в лесном. Присутствие этих видов в сообществах, вероятно, связано с формированием последних на песчаных почвах, прогревающихся быстрее и на большую глубину, чем суглинистые в тех же климатических условиях (Калинина, 1961). Детерминантной является малочисленная группа ксеромезо- и мезоксерофитов: Leontopodium conglobatum, Helictotrichon schellianum, Serratula centauroides.
7. 2. Одноколосковополынные степи (Artemisieta monostachyae)
Степи с преобладанием Artemisia monostachya распространены в Восточном Забайкалье вблизи от высотно-климатического предела формирования степной растительности (Дулепова, 1983, 1993).
Сообщества асс. Koelerio cristatae-Artemisietum monostachyae nov. отмечены в лесном поясе в бассейне р. Ендэ на высоте 1250–1300 м, где приурочены к крутым южным склонам. Почвы песчано-дресвяные, развитые на элюво-делювии гранита. Обильны Koeleria cristata, Festuca litvinovii, Thymus dahuricus; существенную роль играет Eremogone capillaris. Детерминантами ассоциации являются мезоксеро- и ксеромезофиты Pedicularis striata, Heteropappus biennis, Gentiana decumbens, ксерофит Oxytropis oxyphylla и др. Характерная черта – низкая встречаемость или отсутствие многих ксеромезо- и мезофильных видов (Carex pediformis, Galium verum, Potentilla acervata, Sanguisorba officinalis и др.), обычных практически во всех прочих типах степей, развитых в пределах лесного пояса.
7. 3. Полидоминантные разнотравные степи
(Pulsatilleto turczaninovii-Potentilleta acervatae)
В исследуемом районе полидоминантные разнотравные степи с согосподством ксеромезофильных Potentilla acervata, Pulsatilla turczaninovii, Galium verum и др. распространены в лесном поясе, где занимают южные склоны различной крутизны.
Сообщества асс. Poo botryoides-Pulsatilleto turczaninovii-Potentilletum acervatae nov. встречаются в бассейне р. Иля на высоте 900–1000 м. Занимают небольшие участки в нижней части преимущественно облесенных склонов, отличающиеся повышенным увлажнением и накоплением мелкозема. Почвы слабощебнистые, подстилаемые гранитами. Основу травостоя, наряду с доминантами, составляют Galium verum, Stellera chamaejasme, Sanguisorba officinalis, Carex pediformis. Отличительная особенность – участие мезофитов Artemisia tanacetifolia, Vicia amoena, а также лесных кустарников Spiraea media, Rosa davurica. Помимо Artemisia tanacetifolia и Spiraea media, детерминантами выступают горно-степные и лугово-степные ксеромезофиты: Leontopodium conglobatum, Polygala sibirica, Artemisia sericea и др.
В том же районе, где и предыдущий синтаксон, впервые описана асс. Artemisio frigidae-Pulsatilleto turczaninovii-Potentilletum acervatae, сообщества которой приурочены к крутым южным склонам с выходами гранитов. Почва среднещебнистая; вследствие активности эрозионных процессов, поверхность ее покрыта незакрепленным, подвижным щебнем. Типична повышенная роль Potentilla tanacetifolia, и петроксерофильного полукустарничка Artemisia frigida, обилие которого, благодаря биоморфологическим особенностям возрастает на осыпающихся склонах (Дулепова, 1993). К детерминантным видам, кроме Artemisia frigida, относятся растения, тяготеющие к щебнистым и/или песчаным почвам: мезоксерофиты Patrinia rupestris, Chamaerhodos erecta, ксерофиты Allium tenuissimum, Alyssum obovatum и др.
7. 4. Хамеродосовые степи (Chamaerhodeta trifidae)
В Восточном Забайкалье сообщества с преобладанием Chamaerhodos trifida наиболее типичны для Онон-Аргунских степей (Алкучанский Говин, 1964; Флора Сибири, 1988). По мнению Т. И. Исаченко (1965) они представляют собой одну из начальных стадий формирования степной растительности на каменистом субстрате.
Сообщества асс. Orostachyo spinosae-Chamaerhodetum trifidae nov. распространены в лесостепном поясе на высоте 700–850 м в северо-восточной части региона. Формируются на грубоскелетных фрагментарных почвах вершинных и привершинных инсолируемых участков сопок, сложенных сланцами и туфами. Господствующие петроксерофиты (Chamaerhodos trifida, Orostachys spinosa, Arctogeron gramineum, Eremogone capillaris) сосредоточены в приземном подъярусе травостоя; обычны также Festuca litvinovii и Pulsatilla tenuiloba. Детерминантами ассоциации являются ее доминант Chamaerhodos trifida, горно-степные ксерофиты Potentilla sericea, Alyssum obovatum, и ксеромезофит Patrinia sibirica, по каменным россыпям и скалам встречающийся как в лесостепном, так и в лесном поясах и в высокогорьях.
В пределах района исследований для сообществ с господством Chamaerhodos trifida прослеживается западная граница ареала, что, по-видимому, связано с эколого-климатическими требованиями эдификатора.
7. 5. Горноколосниковые степи (Orostachyeta spinosae)
Горноколосниковые степи за пределами Восточного Забайкалья отмечены в Туве и на юго-восточном Алтае, где они являются редкими (Намзалов, 1994), а также в бассейне р. Индигирки (Юрцев, 1981).
Сообщества асс. Arctogeronto graminei-Orostachyetum spinosae nov. встречаются в лесостепном поясе на высоте 800–1200 м по всей исследуемой территории, чаще на северо-востоке. Приурочены к вершинам сопок с грубоскелетными фрагментарными почвами, подстилаемыми сланцами. По структуре и преобладающим видам арктогероново-горноколосниковые степи близки к сообществам предыдущей ассоциации, от которой отличаются повышенной встречаемостью Filifolium sibiricum, Potentilla tanacetifolia, Pulsatilla turczaninovii, тяготеющих к относительно мелкоземистым субстратам. Вероятно, данные фитоценозы, по сравнению с горноколосниково-хамеродосовыми, приурочены к более развитым почвам с большим содержанием мелкозема.
7. 6. Байкальскоковыльные степи (Stipeta baicalensis)
Stipa baicalensis является доминантом луговых и разнотравно-дерновиннозлаковых степей, распространенных в Хангайско-Даурской горнолесостепной, Маньчжурской лесостепной и Монгольской степной провинциях Центральноазиатской степной подобласти (Волкова, 1988; Лавренко и др., 1991). За их пределами байкальскоковыльные степи встречаются в предгорных районах Прихубсугулья (Карамышева, 1981), приангарской лесостепи (Номоконов и др., 1984) и в предгорьях, окружающих Приханкайскую равнину (Куренцова, 1962).
Нами выделена одна ассоциация байкальскоковыльных степей – Carici pediformis-Stipetum baicalensis. Ее сообщества распространены на слабощебнистых почвах в лесостепном поясе на высотах 1000–1200 м. Занимают подгорные шлейфы и вогнутые нижние части склонов, где происходит накопление влаги и мелкозема. Наряду с мезофильными и ксеромезофильными Sanguisorba officinalis, Hemerocallis minor, Galium verum, обильны мезоксерофиты Filifolium sibiricum, Adenophora gmelinii, Scutellaria baicalensis. Обычны малочисленные кустарники: Dasiphora parvifolia, Spiraea aquilegifolia, Rosa davurica. Отличительная черта – присутствие мезоксерофита Pulsatilla multifida и мезофита Veratrum nigrum, нехарактерных для прочих типов петрофитных степей. Байкальскоковыльные степи с согосподством Carex pediformis отмечены на территории от Восточного Хангая до Восточного Забайкалья (Горная лесостепь…, 1983; Карамышева, 1981; Дулепова, 1993).
7. 7. Экологические взаимоотношения петрофитных степей
западной части Ононской Даурии
Основными прямодействующими экологическими факторами, определяющими региональное разнообразие петрофитных степей, являются увлажнение и теплообеспеченность, которые изменяются в зависимости как от абсолютной высоты местности, так и от локальных особенностей местообитаний. Согласно значениям среднего фитоценотического статуса выделенных ассоциаций по градиенту увлажнения (50.7–53.9), изучаемые петрофитные степи развиты в условиях увлажнения, которое может быть охарактеризовано как либо лугостепное, либо типичное для сухих и свежих лугов (Методические указания…, 1974). Это полностью согласуется с положением сообществ в пределах лесостепного и нижней части лесного поясов.
Из числа косвенно действующих факторов наибольшее значение имеет гранулометрический состав почвы. В ряде случаев выявляется существенная роль крутизны склонов. Обусловленная этим фактором степень подвижности субстрата оказывает большое влияние на видовой состав растительности. На крутых щебнистых склонах конкурентное преимущество получают растения, биоморфы которых позволяют им выдерживать погребение щебнем и противостоять смыву.
Амплитуды среднего фитоценотического статуса ассоциаций по градиентам богатства–засоленности почв и пастбищной дигрессии (соответственно 12.6–12.9 и 3.6–4.0) указывают на то, что исследуемые фитоценозы приурочены к почвам, близким по степени богатства, и испытывают слабое влияние выпаса.
Соподчиненность ассоциаций петрофитных степей в зависимости от тех или иных экологических факторов представлена на рис. 3.
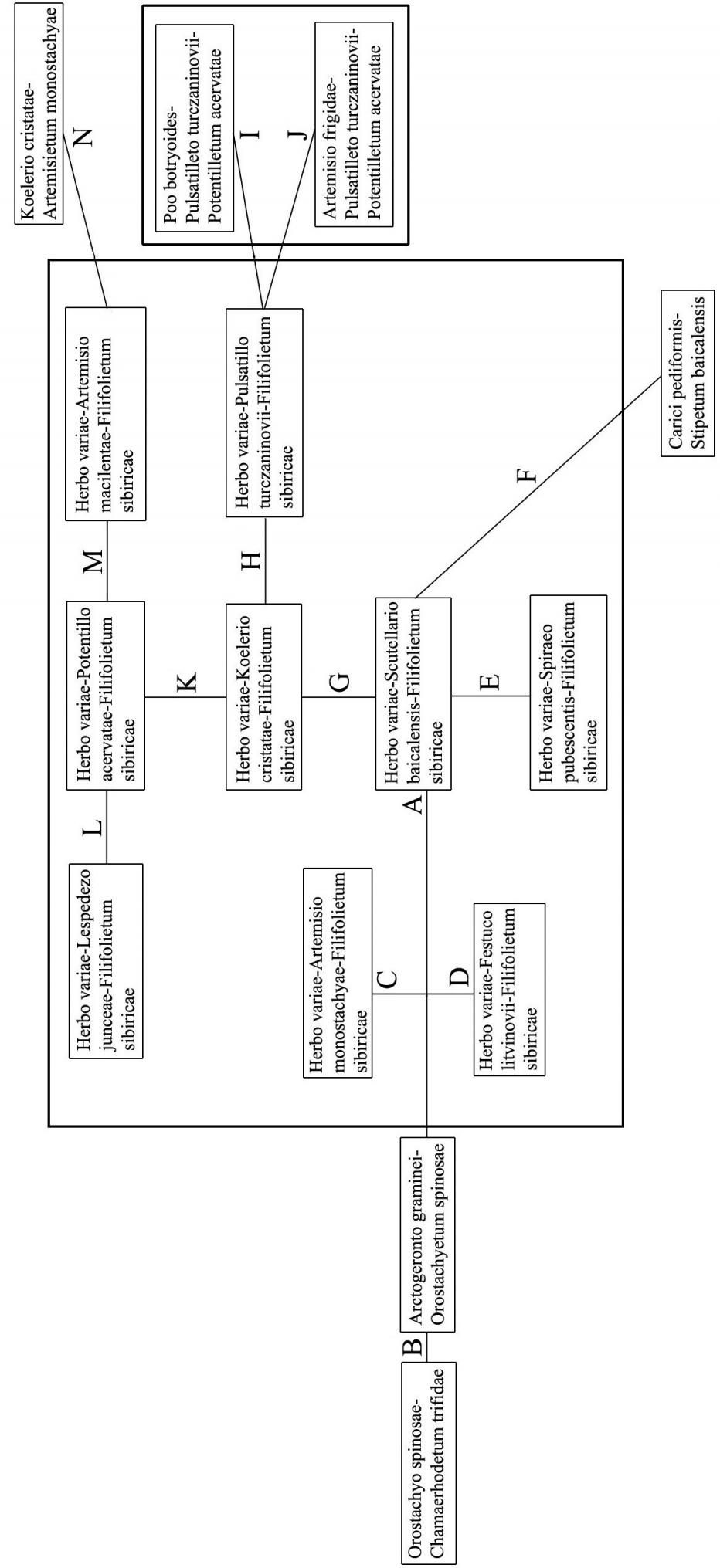
| Рис. 3. Эколого-фитоценотическая схема петрофитных степей западной части Ононской Даурии. |
Выводы
1. Для петрофитно-степной растительности западной части Ононской Даурии характерен высокий уровень синтаксономического разнообразия. Она представлена нителистниковой (Filifolieta sibiricae), полидоминантной разнотравной (Pulsatilleto turczaninovii-Potentilleta acervatae), одноколосковополынной (Artemisieta monostachyae), хамеродосовой (Chamaerhodeta trifidae), горноколосниковой (Orostachyeta spinosae) и байкальскоковыльной (Stipeta baicalensis) формациями, объединяющими 15 ассоциаций и 4 субассоциации.
2. Флора изученных петрофитных степей насчитывает 210 видов сосудистых растений. Спектр семейств, преобладающих по числу видов, указывает на то, что она сочетает черты как аридных, так и бореальных флор. Максимальное фитоценотическое значение имеют представители семейств Asteraceae, Poaceae и Rosaceae.
3. Для всех типов петрофитных степей исследуемого региона характерны сходные соотношения встречаемости различных жизненных форм сосудистых растений, близкие к тем, которые выявляются для подтипа луговых степей. Ведущая роль в сложении петрофитно-степных фитоценозов принадлежит стержнекорневым поликарпическим травам в силу адаптивных преимуществ этой биоморфы на щебнистых, в том числе подвижных субстратах.
4. В районе исследований петрофитные степи распространены в двух высотных поясах: лесостепном и лесном. Все ассоциации характеризуются четкой приуроченностью к тому или иному высотному поясу. Только в лесостепном поясе отмечены горноколосниковые, хамеродосовые и байкальскоковыльные степи; только в лесном – полидоминантные разнотравные и одноколосковополынные степи. В пределах обоих высотных поясов формируются сообщества нителистниковой формации, отличающейся максимальным разнообразием и наиболее широкой экологической амплитудой.
5. Сообщества выделенных ассоциаций различаются по характеру географического распространения. Широко распространены в Восточном Забайкалье, а также за его пределами фитоценозы ассоциаций Herbo variae-Scutellario baicalensis-Filifolietum sibiricae, Herbo variae-Koelerio cristatae-Filifolietum sibiricae, Herbo variae-Pulsatillo turczaninovii-Filifolietum sibiricae, Herbo variae-Lespedezo junceae-Filifolietum sibiricae. Только на территории Восточного Забайкалья до настоящего времени отмечены сообщества ассоциаций Herbo variae-Festuco litvinovii-Filifolietum sibiricae, Herbo variae-Potentillo acervatae-Filifolietum sibiricae, а также ассоциаций, впервые описанных нами. На западной границе своего распространения в исследуемом регионе находятся фитоценозы ассоциации Orostachyo spinosae-Chamaerhodetum trifidae.
6. Основными факторами, определяющими разнообразие петрофитных степей, являются увлажнение, теплообеспеченность, а также гранулометрический состав почвы и крутизна склона.
Список работ, опубликованных по теме диссертации
1. Головина Е. О. Нителистниковые степи западной части Ононской Даурии // Бот. журн. 2009. Т. 94. № 2. С. 244–266.
2. Головина Е. О. О нителистниковых (Filifolium sibiricum (L.) Kitam.) сообществах заказника «Горная степь» // Растения в муссонном климате: Мат-лы IV междунар. конф., г. Владивосток, 10–13 октября 2006 г. Владивосток, 2007. С. 111–113.
3. Головина Е. О. Разнообразие степных сообществ заказника «Горная степь» (Кыринский район Читинской области) // Степи Северной Евразии: Мат-лы IV междунар. симпоз. Оренбург, 2006. С. 196–197.
4. Головина Е. О. Характерные черты петрофитных степей юго-восточного макросклона хребта Становик (Читинская область) // Актуальные проблемы геоботаники: Мат-лы III Всероссийск. школы-конф. Петрозаводск, 2007. С. 152–154.
5. Головина Е. О. О степных сообществах Национального парка «Алханай» (Агинский Бурятский Автономный округ) // Мат-лы междунар. науч. конф. «Биологическое разнообразие азиатских степей». Казахстан, г. Костанай, 3–4 апреля 2007 г. Костанай, 2007. С. 39–40.
6. Беликович А. В., Головина Е. О., Сафронова И. Н. Основные черты растительности Онон-Бальджинского Национального Парка (Монголия) // Растительный и животный мир трансграничной особо охраняемой территории. Тр. Сохондинского заповедника. Вып. 2. Чита, 2007. С. 79–89.
7. Галанин А. В., Беликович А. В., Сафронова И. Н., Роенко Е. Н., Головина Е. О., Коробков А. А. Флора и растительность заказника «Горная степь» // Растительный и животный мир трансграничной особо охраняемой территории. Тр. Сохондинского заповедника. Вып. 2. Чита, 2007. С. 34–78.
8. Сафронова И. Н., Головина Е. О. О степях в верховьях р. Онон (долина реки Галзутый, Читинская область) // Мат-лы междунар. науч. конф. «Биологическое разнообразие азиатских степей». Казахстан, г. Костанай, 3–4 апреля 2007 г. Костанай, 2007. С. 118–120.