

Экологическая структура зоопланктона временных водоемов саратовской области
На правах рукописи
Евдокимов Николай Анатольевич
ЭКОЛОГИЧЕСКАЯ СТРУКТУРА ЗООПЛАНКТОНА
ВРЕМЕННЫХ ВОДОЕМОВ САРАТОВСКОЙ ОБЛАСТИ
03.00.16 – экология
Автореферат диссертации на соискание ученой степени
кандидата биологических наук
Саратов – 2006
Работа выполнена в государственном образовательном
учреждении высшего профессионального образования
«Саратовский государственный университет им. Н.Г. Чернышевского»
на кафедре морфологии и экологии животных
Научный руководитель: кандидат биологических наук, доцент
Ермохин Михаил Валентинович
Официальные оппоненты: доктор биологических наук, профессор
Сачков Сергей Анатольевич
кандидат биологических наук,
старший научный сотрудник
Вехов Николай Владимирович
Ведущая организация: Институт биологии внутренних вод
им. И.Д. Папанина РАН
Защита состоится « 14 » сентября 2006 г. в 10 часов на заседании диссертационного совета Д 212.243.13 при государственном образовательном учреждении высшего профессионального образования «Саратовский государственный университет им. Н.Г. Чернышевского» по адресу:
410012, г. Саратов, ул. Астраханская, д. 83
С диссертацией можно ознакомиться в Зональной научной библиотеке ГОУ ВПО «Саратовский ГУ».
Автореферат разослан «_7_» августа 2006 г.
Ученый секретарь
диссертационного совета С.А. Невский
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования. Временные водоемы – уникальный тип водных экосистем. Небольшая продолжительность существования и малые размеры позволяют использовать их как модельные объекты в фундаментальных исследованиях закономерностей формирования популяций и сообществ гидробионтов.
Строгая избирательность, способность к миграциям в пространстве и времени – основа экологической стратегии обитателей эфемерных биотопов (Бигон и др., 1989). Они определяют высокий уровень эндемизма и специализации видов. В процессе эволюции временных водоемов как особого типа экосистем они стали рефугиумом реликтовых и наиболее консервативных элементов фауны низших ракообразных, например, голых жаброногов. Поэтому, даже в антропогенно трансформированном ландшафте, временные водоемы образуют очаги высокого биоразнообразия (Simovich, 1998).
Зоопланктон временных водоемов – важное звено в формировании внутри и межэкосистемных потоков вещества и энергии. Он удобен для оценки влияния абиотических факторов на экологическую структуру сообщества. Хотя его изучение во временных водоемах России проводилось еще с начала XX века, большинство работ были посвящены фауне и редко структуре сообществ. Зоопланктон временных водоемов богат по видовому составу, но недостаточно изучен, поскольку традиционно внимание планктонологов России было сконцентрировано на крупных хозяйственно значимых водоемах (Волгоградское водохранилище…, 1979). Кроме того, гидрологическая сеть юго-востока Европейской части России подверглась сильной трансформации, что привело к формированию благоприятных условий для возникновения густой сети временных водоемов в ландшафтах, где ранее их существование было невозможно или сильно ограничено аридным климатом. Во второй половине XX в. временные водоемы стали распространенным типом водных экосистем водораздельных ландшафтов Заволжья Саратовской области.
Принято считать, что экологическая структура временных водоемов проста. Однако в ходе анализа структуры планктонных сообществ были выявлены специфические особенности, демонстрирующие ее сложность и специфичность. Выделены механизмы стабилизации сообществ в предсказуемо нестабильной среде: сложные способы избегания конкуренции (Simovich, 1998), диапауза (Алексеев, 1990), банк яиц в грунте (Gomez et al., 2002).
Исследования трофической структуры зоопланктона временных водоемов находится на начальной стадии, что связано с необходимостью выявления пищевой специализации специфичных для этого типа водоемов структурообразующих видов. Как правило, это узкоареальные эндемичные виды, изменяющие пищевую специализацию в постэмбриональном развитии и имеющие широкие пищевые спектры.
Цель и задачи исследования: Цель работы – выявление закономерностей формирования экологической структуры зоопланктона временных водоемов различных природных зон на примере территории Саратовской области. Для достижения поставленной цели необходимо было решить следующие задачи:
1. Установить закономерности формирования видового состава зоопланктона временных водоемов и разработать их типологию по фаунистическим особенностям;
2. Описать динамику популяций структурообразующих видов и выделить сезонные группировки низших ракообразных;
3. Изучить сезонную динамику количественного развития зоопланктона с определением ведущих факторов, оказывающих влияние на его формирование в основных типах временных водоемов;
4. Установить основные трофические группировки зоопланктона на основе анализа трофического статуса структурообразующих видов с учетом его изменения в процессе постэмбрионального развития;
5. Выявить закономерности сезонной динамики трофической структуры зоопланктона.
Научная новизна. Впервые рассмотрена структура зоопланктона на стадии подледной лужи, специфической для гидрологического режима временных водоемов севера Евразии. Определены особенности структуры и фенологии популяций 6 массовых видов зоопланктона временных водоемах степной зоны, выделены сезонные группировки Anostraca и Calanoida. Для Hemidiaptomus hungaricus Kiefer, 1933 установлено крайне низкое значение условного физиологического нуля. Выявлены особенности и общие закономерности сезонной динамики показателей экологической структуры зоопланктона временных водоемов степной зоны и интразональных пойменных ландшафтов. У 10 видов Anostraca и Calanoida проведен сравнительный анализ морфофункциональных особенностей придатков, участвующих в обработке пищи. Впервые описан половой диморфизм в строении мандибул Branchinecta minuta S. Smirnov, 1948, B. orientalis G.O. Sars, 1901, Pristicephalus josephinae Grube, 1853, Chirocephalus horribilis S. Smirnov, 1948, Drepanosurus birostratus Fischer, 1851 и различия их трофического статуса. Установлены закономерности смены типа питания в процессе постэмбрионального развития двух массовых видов каланоид. Впервые выявлены отличительные особенности трофической структуры зоопланктона временных водоемов различных типов в степных и сухостепных ландшафтах.
Научно-практическая значимость. Установленные особенности фенологии и структуры популяций ранее неизученных видов, позволили вскрыть механизмы их адаптаций к гидрологическому режиму. На основе уравнений, описывающих связь структуры популяций и сообществ с параметрами водоемов, возможна разработка математических моделей для прогнозирования реакции сообщества на различные воздействия. Результаты исследований были использованы при определении статуса и разработке мер охраны 12 редких видов низших ракообразных, предлагаемых к внесению во 2-е издание Красной книги Саратовской области. Установленные закономерности влияния факторов среды на структуру сообществ могут быть использованы в оценке их видового разнообразия при проектировании особо охраняемых природных территорий. Материалы включены в программы практикумов по зоологии беспозвоночных, популяционной экологии и большого спецпрактикума Саратовского государственного университета.
Апробация работы. Результаты исследований докладывались на XII Международной конференции молодых ученых «Биология внутренних вод: проблемы экологии и биоразнообразия» (ИБВВ РАН, Борок, 2002), Всероссийской научной конференции «Охрана растительного и животного мира Поволжья и сопредельных территорий» (ПГПУ, Пенза, 2003), Международной конференции «Трофические связи в водных сообществах и экосистемах» (ИБВВ РАН, Борок, 2003), Международном совещании «Биоразнообразие экосистем Поволжья: прошлое, современное состояние, будущее» (ИПЭЭ РАН, Саратов, 2005); научных конференциях Саратовского государственного университета (Саратов, 2001–2005 гг.).
Публикации. Результаты исследования опубликованы в 9 работах.
Декларация личного участия автора. Материал диссертации собран и обработан автором лично. Участие автора в совместных публикациях составляло от 30 до 90%.
Объем и структура диссертации. Работа изложена на 256 страницах и состоит из введения, 6 глав, выводов, списка литературных источников и приложений. В работе приведено 22 рисунка и 37 таблиц. Список цитированной литературы включает 186 источников, в том числе 71 на иностранных языках.
Основные положения, выносимые на защиту:
1. С увеличением продолжительности существования водоема растет общее количество видов, среднее число видов в пробе и доля видов Branchiopoda, по сравнению с Copepoda. При этом Copepoda составляют основу видового разнообразия в водоемах с продолжительностью существования менее 100–120 сут.
2. Гидрологические параметры временных водоемов определяют структуру популяций гидробионтов. Длина тела голых жаброногов в момент наступления половой зрелости зависит от средней глубины водоема на момент его максимального заполнения, а у каланоид – от средней температуры воды в период науплиального развития.
3. Роль различных групп низших ракообразных в формировании зоопланктона временных водоемов убывает в ряду: веслоногие (циклопоиды и каланоиды) – голые жаброноги – ветвистоусые ракообразные.
4. Изменение трофического статуса голых жаброногов и каланоид в процессе постэмбрионального развития сопряжено с динамикой количественного развития зоопланктона временных водоемов в последней декаде мая. Переход к облигатному или факультативному хищничеству происходит при достижении пика биомассы зоопланктона.
5. С увеличением продолжительности существования водоема доминирование в зоопланктоне переходит от собирателей-хватателей к фильтраторам-хватателям, а затем к фильтраторам. Биомасса зоопланктона формируется в основном за счет крупных видов фильтраторов-хватателей.
СОДЕРЖАНИЕ РАБОТЫ
Во введении обосновывается актуальность исследования, отмечается практическая и теоретическая значимость работы, формулируются основные цель и задачи исследования.
Глава 1. ОСОБЕННОСТИ ЭКОСИСТЕМ ВРЕМЕННЫХ ВОДОЕМОВ И АДАПТАЦИИ ЗООПЛАНКТОНА К СРЕДЕ ОБИТАНИЯ
(обзор литературы)
В обзоре литературы показано своеобразие абиотических факторов среды во временных водоемах, общие для биотопов данного типа свойства, обусловливающие обеспеченность пищевыми ресурсами. Обсуждены методические допущения при выделении зоопланктона временных водоемов. Рассмотрены характерные для зоопланктона временных водоемов адаптивные стратегии и механизмы их реализации. Сформулированы возможные причины формирования его высокого видового разнообразия и эндемизма во временных водоемах.
Глава 2. МАТЕРИАЛ, МЕТОДИКИ ИССЛЕДОВАНИЯ И ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА РАЙОНА ИССЛЕДОВАНИЙ
Материал собран в 1997–2003 гг. на трех участках в различных природных зонах Саратовской области: лесостепном, степном и сухостепном. Видовой состав сообществ определяли в 20 водоемах, а исследования структуры – на примере 13 водоемов.
Исследования водоемов начинали с первой ранневесенней оттепели (рис. 1).

Рис. 1. Динамика среднесуточной температуры воздуха и температуры воды на начальных этапах формирования водоема (№ 10; 1999 г.): пунктирная линия – температура воды, сплошная – температура воздуха. С 10 по 25 марта – стадия подледного водоема
Анализ морфометрических показателей и температурного режима водоемов проводили по общепринятым методикам (Близняк, 1952; Мордухай-Болтовской, 1975; Муравейский, 1960). Карбонатную жесткость воды (34 пробы) определяли по общепринятой методике (Экологический мониторинг, 2005).
Структуру популяций изучали по методике Н.В.Вехова (1989). Жизненные циклы описывали в 1999 и 2001 гг. в степных водоемах по методике Е.В.Боруцкого (1960). Измерение средней температуры воды при науплиальном развитии Cyclopoida проводили в сезон, предшествующий определению средних размеров половозрелых рачков. Для характеристики популяций использовали длину тела рачков среднюю (Lср), на момент полового созревания (L0 или L12), максимальную (Lmax), плотность (N), биомассу (B), условный физиологический ноль (t0), сумму эффективных температур (K), соотношение полов и размерно-возрастных групп.
Способ добывания пищи голых жаброногов определяли, используя три признака: 1. Морфология мандибул и торакопод; 2. Содержимое кишечника; 3. Поведение при добывании пищи в природе и в лабораторных условиях (Fryer, 1983; Mura, 1991). Проанализировано около 80 экз. рачков (самцы и самки 9 видов), содержимое кишечников определено у 46 экз. (самцы и самки 6 видов).
Способ добывания пищи Calanoida на различных стадиях постэмбрионального развития определяли по общепринятым методикам (Монаков, 1976; Сажина, 1985). Изучено 50 экз. половозрелых рачков (8 видов) и около 100 экз. науплиусов и копеподитов Hemidiaptomus rylovi Charin, 1928 и H. hungaricus.
Количественные пробы объемом 50 л отбирали сетью Апштейна и обрабатывали по общепринятой методике (Мордухай-Болтовской, 1959; Методические рекомендации..., 1982). Объем материала составил 577 количественных и 154 качественных пробы. При анализе фауны использовали следующие показатели: общее число видов (S) и среднее их число в стандартной пробе (Sср), встречаемость (p), индекс сходства Жаккара (IJ), соотношение числа видов Branchiopoda и Copepoda (SBr./SCop.). Типологию водоемов по сходству видового состава проводили с применением статистических методов, реализованных в программе “Кластерный анализ” (XYZ Soft, 2001).
Структуру сообществ анализировали в 2002–2003 гг. Среднюю индивидуальную массу особей определяли по справочным таблицам (Балушкина, Винберг, 1978). Структуру сообществ анализировали по общеизвестным показателям: видовому разнообразию (HN и HB), индексу доминирования Симпсона (D), индивидуальному весу особи (w), числу структурообразующих видов (n), доли численности (NAn.:NCla.:NCal.:NCycl.) и биомассы (BAn.:BCla.:BCal.:BCycl.) таксона от общей численности (биомассы), соотношению биомассы Cyclopoida и Calanoida (BCycl./BCal.), отношению минимальной численности и биомассы к максимальным за сезон (Nmin/Nmax и Bmin/Bmax соответственно). При анализе трофической структуры выделялись трофические группировки, различающиеся по способу добывания пищи. Статистическую обработку проводили по общепринятым статистическим методам, реализованным в пакете программ Statistica 6.0 (StatSoft, 2001).
Глава 3. ВИДОВОЙ СОСТАВ ЗООПЛАНКТОНА ВРЕМЕННЫХ ВОДОЕМОВ
Видовой состав зоопланктона временных водоемов Саратовской области включает 68 видов низших ракообразных, в том числе: голых жаброногов – 9, ветвистоусых ракообразных – 25, каланоид – 10 и циклопоид – 24 вида. Группировка голых жаброногов Саратовской области одна из наиболее богатых по видовому составу среди аналогичных сообществ на территории бывшего СССР, причем для большинства видов характерна низкая частота встречаемости.
Планктонные ракообразные представлены видами 4-х фаунистико-географических комплексов: северного, холодноводного и тепловодного умеренных широт и южного. В фауне голых жаброногов отмечено 4 пресноводных и 4 вида, характерных для слабоминерализованных водоемов; положение B. minuta из-за слабой изученности не определено. Cladocera и Cyclopoida содержат мало специализированных видов, обитающих только во временных водоемах, тогда как среди Calanoida доля таких видов высока, а среди Anostraca – все виды специфичны для временных водоемов.
Наименьшее число видов характерно для водоемов долины р. Медведицы (33 вида). Гораздо выше оно в водоемах степей Заволжья (39 видов) и достигает пика в сухих степях (52 вида). Увеличение числа видов происходит за счет трех таксономических групп: Anostraca, Cladocera и Calanoida. Наибольшая общность видового состава обнаружена между водоемами степей и сухих степей Заволжья, которые сильно отличаются от лесостепных водоемов долины р. Медведицы. Для голых жаброногов характерна высокая специализация и узкая ландшафтная приуроченность.
Роль пресноводных видов наиболее значима в фауне водоемов поймы Медведицы. В направлении с северо-запада на юго-восток увеличивается доля видов фауны слабоминерализованных водоемов. На фауну водоемов в сухой степи влияет усиление аридности и значительное воздействие на гидрологический режим временных водоемов вод, поступающих из оросительных каналов.
Общее число видов в отдельных водоемах составляет 4–34 вида, среднее число видов в пробе – 2–13 видов. Общее число видов в некоторых временных водоемах превышает таковое для постоянных. В связи с межгодичными изменениями гидрологического режима обнаружены существенные отличия видового состава зоопланктона. Наиболее выражены они в водоемах сухих степей, заполняемых из оросительных каналов, а в водоемах на степных водоразделах количество видов в разные годы изменяется незначительно.
Число видов основных групп и соотношение между Branchiopoda и Copepoda сильно зависят от продолжительности существования и морфометрических показателей водоемов. Выявлены зависимости общего числа видов и среднего числа видов в пробе от продолжительности существования и площади поверхности временных водоемов в период максимального заполнения (табл. 1).
Число видов Anostraca и Calanoida увеличивается незначительно (до 5 и 7 соответственно), то у Cladocera и Cyclopoida оно изменяется от 1 до 18 и от 3
Таблица 1
Влияние некоторых гидрологических и морфометрических характеристик водоема
на количественные показатели фауны зоопланктона по результатам
множественного регрессионного анализа
| Характеристика | Sср. | S | SBr./SCop. | SCl. | SCal. | SCycl. | ||||||
| r | p | r | p | r | p | r | p | r | p | r | p | |
| T | – | – | 0.72 | 0.004 | 0.74 | 0.003 | 0.73 | 0.003 | – | – | 0.53 | 0.05 |
| V | – | – | – | – | 0.58 | 0.03 | 0.66 | 0.01 | – | – | – | – |
| Sh | 0.62 | 0.02 | 0.60 | 0.03 | – | – | 0.69 | 0.01 | 0.67 | 0.01 | – | – |
Примечание: T – продолжительность существования водоема, V – объем водоема на момент максимального заполнения, Sh – площадь поверхности водоема на момент максимального заполнения, SCl. – число видов Cladocera, SCal. – число видов Calanoida, SCycl. – число видов Cyclopoida.
до 12 соответственно. В постоянных водоемах преобладают виды Calanoida, в то время как в пересыхающих водоемах превалируют виды Cyclopoida.
На основании фаунистического сходства выделено 7 групп водоемов: I. Водоемы пойм; II. Водоемы надпойменных террас; III. Временные водоемы степных водоразделов; IV. Лиманы; V. Лиманы, связанные с оросительными системами; VI. Копаные пруды; VII. Лужи.
Видовой состав постоянных водоемов сухих степей по качественному составу не имеет существенных отличий от временных. Наиболее своеобразна фауна лиманов, где выявлены голые жаброноги P. josephinae, C. horribilis, В. orientalis, Branchipus schaefferi Fischer, 1834, каланоиды H. rylovi, Diaptomus mirus Lilljeborg, 1889 и Arctodiaptomus sp.
Глава 4. ВЛИЯНИЕ ОСОБЕННОСТЕЙ ВРЕМЕННЫХ ВОДОЕМОВ
НА ФЕНОЛОГИЮ И СТРУКТУРУ ПОПУЛЯЦИЙ МАССОВЫХ ВИДОВ ЗООПЛАНКТОНА
Голые жаброноги объединяются в 3 сезонные группировки: 1) ранневесеннюю (B. orientalis, Tanymastix stagnalis Linne, 1758, D. birostratus); 2) весеннюю (B. minuta, P. josephinae, Branchinecta ferox Milne-Edwards, 1840); 3) весенне-летнюю (B. schaefferi, C. horribilis, Streptocephalus torvicornis Waga, 1842). Нижний температурный порог жизнедеятельности различных видов находится в диапазоне от 2 до 10°С. Сроки прохождения этапов жизненного цикла, продолжительность жизни, длина тела рачков на момент наступления половой зрелости и максимальная длина тела определяются особенностями гидрологического и температурного режима водоемов. Установлена положительная зависимость средней длины тела C. horribilis на момент наступления половой зрелости от средней глубины водоема (r=0.98, p=0.03). На примере водоемов сухой степи выявлена зависимость средней длины половозрелых особей P. josephinae от гидрологического режима водоема, формирующегося в различные годы при изменении объемов подачи воды. Наибольшую плотность и биомассу популяции T. stagnalis имеют в лужах, в лиманах – C. horribilis.
Развитие ряда массовых видов Cladocera начинается при температуре воды близкой с таковой для видов Anostraca. Однако устойчивый рост численности популяций начинается лишь при достижении 15–20°С. В массовом количестве во временных водоемах развиваются популяции Daphnia hyalina Leydig, 1860; D. pulex Leydig, 1860; D. magna Straus, 1820; Moina brachiata Jurine, 1820; Simocephalus vetulus O.F. Muller, 1776; Ceriodaphnia reticulata Jurine, 1820 в весенне-летний период с высокой температурой воды. Продолжительность этапов жизненного цикла определяют многие факторы: продолжительность существования водоема, особенности гидрологического и температурного режимов, зарастание растительностью, развитие популяций других видов. Минимальные сроки существования популяций Cladocera выявлены на примере S. vetulus – 1.5 мес. У Cladocera обнаружен нерегулярный характер развития популяций в течение ряда лет. Для сезонной динамики плотности характерен экспоненциальный рост. Наибольшая плотность и биомасса выявлена у фитофильных видов S. vetulus (водоемы V группы), C. reticulata и прудового вида D. hyalina (водоемы III группы). В водоеме № 10 плотность C. reticulata и S. vetulus подвержена циклическим изменениям с периодом в 7–8 сут.
Массовые виды Calanoida можно разделить на три сезонных группировки: 1) весеннюю (моноциклические виды H. rylovi, H. hungaricus и D. mirus); 2) переходную весенне-летнюю (полициклические виды Eudiaptomus transylvanicus Daday, 1890; E. vulgaris Schmeil, 1898; Arctodiaptomus dentifer Smirnov, 1928 и Arctodiaptomus sp.); 3) летнюю (Acanthodiaptomus denticornis Wierzejski, 1887 и Lovenula alluaudi Guerne et Richard, 1890). Виды весенней группы отличаются наиболее низким температурным порогом активации яиц и крайне специфическими условиями прохождения начальных этапов жизненного цикла. Для H. hungaricus характерно низкое значение условного физиологического нуля –4–4.5°С. Нижний температурный порог видов весенне-летней и летних групп находился в пределах от 4 до 10°С. Эти виды (E. vulgaris, A. denticornis) формируют группу со средними значениями условного физиологического нуля (3–4°С) (табл. 2).
Таблица 2
Условный физиологический ноль, температурные пороги (tmin, tmax) и
сумма градусодней, необходимых для развития различных видов Calanoida
| Название вида | to, °C | tmin, °C | tmax, °C | K, градусодни | ||
| науплиусы | копеподиты | взрослые рачки | ||||
| H. hungaricus | –4 | 0 | 20 | 164–212 | 194–284 | 293–600 |
| D. mirus | –4* | 0 | 20 | 140–170 | 180–220 | 250–500 |
| H. rylovi | –4* | 0 | 25 | 150–265 | 380–433 | 600–1000 |
| Arctodiaptomus sp. | – | 4–6 | 30 | – | – | – |
| E. vulgaris | 3** | 8 | 30 | 120 | 220 | – |
| A. denticornis*** | 4 | 10 | 22 | 218–319 | 397 | – |
*Экстраполяция данных, **указано оценочное значение, ***по Иванова, 1985
Продолжительность этапов жизненного цикла и число генераций (у полициклических видов) определяется, с одной стороны, суммой эффективных температур, с другой – особенностями температурного и гидрологического режимов водоемов. Последние два фактора также детерминируют среднюю длину тела взрослых рачков H. rylovi, H. hungaricus, D. mirus, E. vulgaris и Arctodiaptomus sp. Средняя длина тела находится в зависимости от средней температуры воды, при которой происходит развитие науплиусов (рис. 2). Данная зависимость подтверждает ранее установленный факт (Афанасьева, 1975), что именно средняя температура воды, при которой происходит рост науплиусов, определяет среднюю длину тела половозрелых рачков в популяции.
В сезонной динамике плотности популяций Calanoida выделяется два крайних типа: экспоненциальное убывание и рост. Первый тип наиболее характерен для весенних видов в водоемах со скоростью падения уровня воды 0.5–1 см/сут. Науплиусы таких видов имеют высокую плотность (60–80 тыс. экз./м3) и синхронно выходят из диапаузы в начале марта – апреле. Плотность копеподитов снижается на порядок по сравнению с науплиусами, а у половозрелых особей составляет от нескольких тысяч до десятков экз./м3. В водоемах со скоростью падения уровня воды 4–10 см/сут естественная убыль рачков перекрывается концентрированием в оставшемся объеме водной массы, в результате чего наблюдается экспоненциальный рост плотности популяции.

Рис. 2. Зависимость средней длины тела половозрелых рачков (L12) от средней температуры воды, при которой происходит развитие науплиусов (tn) пяти массовых видов Calanoida: H. rylovi, H. hungaricus, D. mirus, E. vulgaris, Arctodiaptomus sp.
Динамика плотности популяций полициклических видов характеризуется, как правило, экспоненциальным ростом. Для весенних генераций весенне-летних видов свойственна низкая плотность популяции и растянутость сроков выхода науплиусов. У летнего вида A. denticornis науплиусы наблюдаются с последней декады апреля до середины июля. Сезонная динамика плотности популяций летних генераций полициклических и летних видов в значительной мере определяется характером конкурентного взаимодействия с представителями летнего планктонного сообщества. При отсутствии конкуренции плотность летних генераций E. vulgaris и Arctodiaptomus sp. достигает 300–600 тыс. экз./м3. При доминировании конкурирующих видов Cladocera средняя плотность Calanoida ниже – 100–200 экз./м3.
Сезонная динамика биомассы детерминирована значительным количеством факторов, например, половой структурой популяций. У весенних видов H. hungaricus, D. mirus при увеличении продолжительности существования водоема доля самцов снижается от 86 до 65%. В популяциях E. vulgaris количество самцов и самок в среднем равно, а у летних видов L. alluaudi и A. denticornis самки преобладают.
Другим структурным показателем, влияющим на биомассу, является возрастная структура популяции. Моноциклические весенние виды, в силу синхронности развития, имеют наиболее простую возрастную структуру популяции. В летних генерациях Arctodiaptomus sp. одновременно встречаются представители всех стадий жизненного цикла, при этом основную ее часть составляют копеподиты. В популяциях летних генераций E. vulgaris, в отличие от Arctodiaptomus sp., характерно преобладание половозрелых рачков над копеподитами.
В сезонной динамике биомассы популяций H. hungaricus, D. mirus, H. rylovi можно выделить две фазы. Первая фаза характеризуется экспоненциальным ростом до достижения пика биомассы, приходящегося на момент наступления половой зрелости. Вторая фаза – вымирание половозрелых рачков, продолжающаяся в течение длительного периода времени. Наибольшая плотность и биомасса популяций H. hungaricus, H. rylovi, E. vulgaris наблюдается в водоемах III группы с высокой степенью постоянства гидрологического и температурного режимов в течение ряда лет, у Arctodiaptomus sp. – в водоемах V группы, а у D. mirus – в водоемах IV группы.
У Cyclopoida по сезонной динамике их роли в сообществах можно выделить только группу типичных обитателей временных водоемов (Cyclops strenuus Fischer, 1851; C. furcifer Claus, 1857; Metacyclops minutus Claus, 1863, D. bisetosus), которые являются наиболее многочисленными. В некоторых временных водоемах C. furcifer вместе с копеподитами D. bisetosus формируют основу сообщества подледных луж. Выход из диапаузы IV копеподитов происходит при крайне низкой температуре (0–0.5°С), сразу после начала заполнения котловины водоема водой.
Продолжительность жизненного цикла моноциклических C. furcifer и D. bisetosus определяются, как правило, особенностями температурного режима водоема. Число генераций у полициклических C. strenuus и M. minutus, варьирует в зависимости от продолжительности существования водоема. Средняя длина тела половозрелых C. furcifer и M. minutus находится в экспоненциальной зависимости от средней температуры воды в период развития науплиусов.
Сезонная динамика плотности популяций Cyclopoida аппроксимируется кривой экспоненциального роста. При этом, чем менее продолжительны сроки существования водоема, тем более высока суммарная плотность копеподитов Cyclopoida. Средняя плотность половозрелых особей весенних генераций изменяется в пределах 0.1–10.0 тыс. экз./м3. Наибольшая плотность формируется в сильно усыхающих водоемах – 70 тыс. экз./м3. В лужах наиболее велика плотность и биомасса половозрелых C. furcifer (3–5 тыс. экз./м3), во временных водоемах поймы – C. strenuus, в лиманах половозрелых D. bisetosus (8–10 тыс. экз./м3), половозрелых особей весенней генерации M. minutus – в лужах, летних генераций – в водоемах V группы.
Глава 5. СТРУКТУРА И СЕЗОННАЯ ДИНАМИКА ЗООПЛАНКТОНА ОСНОВНЫХ ГРУПП ВРЕМЕННЫХ ВОДОЕМОВ
В формировании зоопланктона можно выделить три фенологических периода: раннюю весну, весну, лето. Ранневесенний период в большинстве типов водоемов отличается наименьшим средним числом видов, основу которого составляют Cyclopoida. Они определяют также численность сообщества, а наибольшую биомассу формируют Calanoida и Anostraca. Однако численность, биомасса и общее видовое разнообразие в это время наименьшие. Весенний период характеризуется наибольшим средним числом видов, основу которого также составляют Cyclopoida. Это обусловлено одновременным присутствием в сообществе популяций видов из различных сезонных группировок. Весенний период выделяется наибольшим числом структурообразующих видов. Основу численности сообщества составляют Cyclopoida, биомассу почти в равной мере формируют Anostraca и Cyclopoida. В весенний период преобладают относительно крупные виды Anostraca и Calanoida из весенней группировки, поэтому индивидуальный вес особей был максимальным, а биомасса сообщества достигала наибольшего значения за сезон (рис. 3).
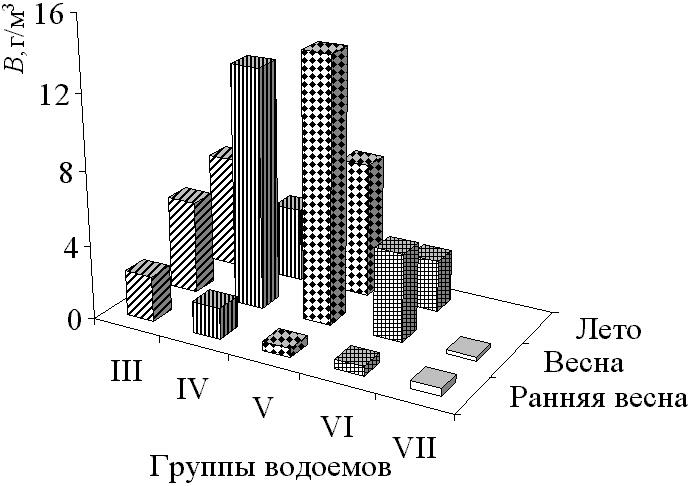
Рис. 3. Сезонная динамика биомассы в различных группах временных водоемов
В летний период происходит незначительное уменьшение среднего числа видов, а основу его составляют Cladocera. Calanoida и Cladocera, в равной мере, участвуют в формировании облика сообщества. В водоемах с незначительным антропогенным влиянием численность и видовое разнообразие выше, а средний вес организмов наименьший. Таким образом, наибольшие значения числа видов, биомассы и индивидуального веса особей достигаются в весенний период. Общая тенденция в сезонной динамике численности и видового разнообразия, рассчитанного по численности – рост с летним пиком. Численность сообщества определяют Cyclopoida, биомассу Anostraca и Calanoida.
Результаты корреляционного анализа показали, что средние за сезон показатели зоопланктона зависят, в первую очередь, от продолжительности существования временных водоемов. Наиболее значимые связи этого параметра выявлены с числом видов Cladocera, числом структурообразующих видов (по биомассе) и средним за сезон числом видов одновременно присутствующих в сообществе. Ряд значимых связей выявлены с площадью и максимальной глубиной водоема на момент его заполнения (табл. 3).
Зоопланктон водоемов различных групп значительно различается по среднему числу видов, одновременно присутствующих в сообществе в течение сезона. Наиболее велико значение этого показателя в лиманах и лиманах, связанных с оросительными системами, а наименьшее – в зоопланктоне луж. Для лиманов выявлено наибольшее число видов Anostraca, Calanoida и Cyclopoida. Лиманы, связанные с оросительными системами, отличаются наибольшим числом видов Cladocera и наименьшим Anostraca.
Таблица 3
Корреляционные зависимости структурных показателей
зоопланктона от морфометрических параметров водоема
| Показатели | Параметр | ||||
| T | hср | hmax | V | Sh | |
| Sср.An. | – | – | – | –0.51 | – |
| Sср.Cl. | 0.79 | – | – | – | 0.61 |
| Sср.Cal. | – | –0.49 | –0.54 | – | – |
| Sср. | 0.59 | – | – | – | 0.48 |
| N | 0.52 | – | – | – | – |
| B | 0.56 | – | – | – | – |
| NCycl./NCal. | –0.56 | – | – | – | – |
| BCycl./BCal. | –0.49 | – | – | – | – |
| HN | 0.48 | – | – | – | 0.47 |
| HB | – | – | –0.52 | – | – |
| nN | 0.55 | – | –0.50 | – | – |
| nB | 0.62 | – | – | – | 0.52 |
Примечание: Sср.An. – среднее число видов Anostraca, Sср.Cl. – среднее число видов Cladocera, Sср.Cal. – среднее число видов Calanoida, nN – число структурообразующих видов (по численности), nB – число структурообразующих видов (по биомассе), hср – средняя глубина на момент максимального заполнения, hmax – максимальная глубина на момент максимального заполнения.
По кривым ранжирования было выявлено 2 крайних типа структуры зоопланктона. Первый тип характерен для сообществ водоемов V группы, отличающихся максимальным числом структурообразующих видов, высокой выравненностью их численности и биомассы. Второй тип кривых выявлен для зоопланктона луж, характеризующегося минимальным числом структурообразующих видов, крайне неоднородной численностью и биомассой.
В большинстве временных водоемов доминантами по численности выступают различные виды Cyclopoida и лишь в некоторых доминируют Cladocera. По биомассе приблизительно в равном количестве временных водоемов доминируют Calanoida, а в других – Cyclopoida. Незначительно им в качестве доминантов уступают Anostraca.
Отношение Nmin/Nmax, характеризующее сезонные изменения численности, зависит от динамики численности массовых видов. Наибольшие сезонные изменения выявлены в водоемах с незначительным сукцессионным возрастом. Значительные сезонные вариации численности происходят в водоемах, находящихся под влиянием оросительной системы.
Средний индивидуальный вес особей находится в пределах от 0.005 до 2.615 мг, что обусловлено доминированием во временных водоемах, по сравнению с постоянными, относительно крупных видов низших ракообразных. Как следствие, амплитуда динамики средней биомассы зоопланктона за сезон составляет 0.029–33.139 г/м3.
Отношение Bmin/Bmax – интегральная величина, характеризующая сообщество и свидетельствующая о его продуктивности (Алимов, 2000). Наибольшая вариабельность этого показателя отмечена для водоемов с искусственно регулируемым уровнем воды, наименьшая – для лиманов. Основу численности зоопланктона в большинстве водоемов составляют Cyclopoida. Биомассу определяли различные таксоны, в зависимости от группы временных водоемов (табл. 4). Наиболее высокое видовое разнообразие и стабильность структурных параметров выявлены в сухостепных лиманах.
Таблица 4
Средняя за сезон доля биомассы основных таксонов зоопланктона
в различных группах временных водоемов, %
| Таксон | Группа водоемов | ||||
| III | IV | V | VI | VII | |
| Anostraca | 13.1 (5.9–29.5) | 42.9 (0.5–92.5) | 0.03 (0–0.1) | 6.7 | 38.0 (0–82.1) |
| Cladocera | 9.7 (1.0–22.1) | 14.3 (0–30.9) | 69.9 (49.8–89.9) | 2.6 | 0 |
| Calanoida | 68.7 (41.2–88.0) | 29.5 (6.8–55.3) | 10.5 (8.9–12.1) | 31.8 | 10.7 (1.6–29.6) |
| Cyclopoida | 8.6 (1.4–30.2) | 13.2 (0.7–25.5) | 19.6 (1.1–38.0) | 58.9 | 51.4 (5.9–98.1) |
Примечание: в скобках указан диапазон изменения показателя
Глава 6. ТРОФИЧЕСКАЯ СТРУКТУРА ЗООПЛАНКТОНА ВРЕМЕННЫХ ВОДОЕМОВ
Трофическая структура сообществ зоопланктона временных водоемов изучена чрезвычайно слабо. Во-первых, структуру зоопланктона определяют ранее не изученные виды или виды, трофический статус которых нуждается в уточнении. Во-вторых, динамика условий среды определяет высокую изменчивость кормовой базы. В-третьих, размеры пищевых объектов сильно варьируют. Пищевая специализация Cladocera и Cyclopoida и закономерности ее изменения изучены достаточно полно (Монаков, 1976). Анализу трофической структуры зоопланктона временных водоемов в рамках нашей работы предшествует установление трофического статуса массовых видов Anostraca и Calanoida.
Голые жаброноги производят захват и обработку пищи с использованием торакопод и мандибул. В строении торакопод выделяли три функционально важных морфологических показателя: 1) относительная длина экзоподита (отношение длины части экзоподита, выступающей за дистальный край эндоподита, к длине торакоподы); 2) степень развития и форма эндоподита; 3) строение и вооружение II–V эндитов (рис. 4).
По строению экзоподита голых жаброногов можно разделить на 3 группы: 1. У B. minuta, B. schaefferi, S. torvicornis относительная длина экзоподита близка к 30%; 2. У B. orientalis, P. josephinae, D. birostratus, T. stagnalis, C. horribilis – 15–25%; 3. У B. ferox – близок к 0, т.е. экзоподит практически не выступает над эндоподитом, а их дистальные края находятся почти на одной линии. Степень подвижности и приуроченность к верхним слоям воды, установленная наблюдениями в природных водоемах, снижается в этом ряду от первой группе к третьей.


Рис. 4. 5-я торакальная конечность самки B. minuta (а) и её дистальные (III–V) эндиты (б): 1 – эндиты, 2 – эндоподит, 3 – экзоподит, 4 – эпиподит, 5 – экзиты, 6 – основание конечности, 7 – длина торакоподы, 8 – длина части экзоподита, выступающей за дистальный край эндоподита, 9 – III эндит, 10 – IV эндит, 11 – V эндит, 12 – передний ряд вооружения, 13 – задний ряд
По строению эндоподита также можно выделить 3 группы голых жаброногов. 1. У B. ferox, B. orientalis, P. josephinae, самок B. minuta эндоподит не выступает за линию эндитов. 2. У T. stagnalis, B. schaefferi, S. torvicornis, самцов B. minuta, самок C. horribilis и D. birostratus эндоподит выступает за линию эндитов, а его внутренний край образует тупой угол с осью конечности. 3. У самцов C. horribilis и D. birostratus лопатообразный эндоподит сильно склеротизирован и значительно выступает за линию эндитов.
Строение и вооружение II–V эндитов позволяет судить о способности противостоять активному сопротивлению потенциального пищевого объекта и удержать его. В результате сравнительного анализа по этому признаку сформировано три группы. В ряду групп: 1) виды рода Branchinecta; 2) P. josephinae, C. horribilis, D. birostratus; 3) T. stagnalis, B. schaеfferi, S. torvicornis происходит уменьшение общей ширины II–V эндитов, уменьшается количество шипов в переднем ряду вооружения и увеличивается число щетинок в заднем ряду (в целом ослабляется жесткость структур, слагающих вооружение эндитов) (табл. 5). Способность к активному захвату и удержанию подвижной жертвы уменьшается.
Таблица 5
Вооружение II–V эндитов 5-ой пары торакопод самцов голых жаброногов
| Название вида | Эндиты | |||||||
| II | III | IV | V | |||||
| a. | p. | a. | p. | a. | p. | a. | p. | |
| B. ferox | 1 | 9 | 10 | 5 | 11 | 4 | 17 | 2 |
| B. orientalis | 2 | 9 | 7 | 3 | 8 | 2 | 6 | 2 |
| B. minuta | 2 | 8 | 6 | 3 | 6 | 2 | 4 | 2 |
| P. josephinae | 1 | 20 | 2 | 3 | 2 | 2 | 6 | 2 |
| C. horribilis | 2 | 16 | 2 | 3 | 2 | 2 | 4 | 2 |
| D. birostratus | 1 | 15 | 2 | 3 | 2 | 2 | 2 | 2 |
| T. stagnalis | 2 | 20 | 2 | 3 | 2 | 2 | 1 | 2 |
| B. schaefferi | 2 | 24 | 2 | 3 | 2 | 2 | 1 | 2 |
| S. torvicornis | 2 | 26 | 2 | 3 | 2 | 2 | 1 | 2 |
Примечание: a. – передний ряд; p. – задний ряд
Строение мандибул голых жаброногов имеет ряд специфических черт, затрудняющих их типологию. Для сравнительного анализа использовали особенности строения задне-дорзальной области молярной поверхности левой мандибулы, свидетельствующей по данным Д. Фрайера (1983) о размерах потенциальных пищевых объектов и характере их обработки (рис. 5).
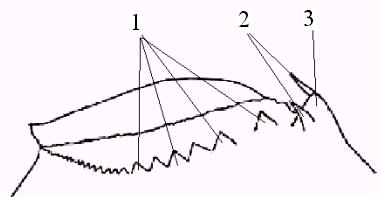
Рис. 5. Строение задне-дорзальной области левой мандибулы самца B. minuta: 1 – группа зубцов задне-дорзальной борозды, 2 – апикальные задние шипы, 3 – апикальный задний зубец
По этому признаку все изученные виды можно разделить на 4 группы: 1) B. ferox, B. orientalis, B. minuta; 2) B. schaefferi, T. stagnalis, C. horribilis, P. josephinae; 3) D. birostratus; 4) S. torvicornis. В ряду четырех групп мощный апикальный задний зубец (зубцы) заменяются шипами, которые у последнего вида исчезают, количество зубцов в борозде увеличивается, но они уменьшаются в размерах. Подобные изменения структуры мандибул ведут к уменьшению размеров потенциальных пищевых объектов, а характер обработки изменяется от прокалывания и разрыва покровов к механическому перетиранию пищевых объектов.
Выявленная ранее морфофункциональная зависимость пищевой специализации от особенностей строения плавательных ног и мандибул рачков (Fryer, 1966, 1983; Mura, 1995) позволяет нам, с большой долей достоверности, сопоставить выявленные типы строения с определенными типами функционирования. Результаты морфофункционального анализа подтверждаются данными кластерного исследования и содержимого кишечников.
Таким образом, сравнительный морфофункциональный анализ мандибул и торакопод взрослых рачков, прямые наблюдения за пищевым поведением рачков свидетельствует, что в сообществе существуют пять трофических групп голых жаброногов (табл. 6).
Таблица 6
Трофические группировки голых жаброногов
| Способ захвата пищи | Трофический уровень | Основная пища | Название вида |
| Фильтрация | II | Фитопланктон, детрит | S. torvicornis, B. schaefferi, T. stagnalis |
| Соскабливание, фильтрация, захват | II–III | Детрит, фитопланктон, зоопланктон | P. josephinae, самцы C. horribilis, D. birostratus |
| Фильтрация и захват | II–III | Фито- и зоопланктон | самки C. horribilis, D. birostratus |
| Захват и фильтрация | III | Зоо- и фитопланктон | самки B. minuta |
| Захват и соскабливание | III | Зоо- и фитопланктон, детрит | B. orientalis, B. ferox, самцы B. minuta |
Для установления типа захвата пищи ранее не изученных каланоид L. alluaudi, D. mirus, H. rylovi, H. hungaricus, Arctodiaptomus sp. использован анализ морфофункциональных особенностей ротовых придатков: мандибул, максиллул, максилл, максиллипед. По совокупности этих признаков можно выделить три основные морфофункциональные группы видов: 1) фильтраторы (E. vulgaris, A. denticornis, E. affinis и Arctodiaptomus sp.), 2) фильтраторы-хвататели (L. alluaudi, H. hungaricus и D. mirus), 3) активные хвататели, способные к фильтрации (H. rylovi). Полученные результаты подтверждены наблюдениями пищевого поведения Calanoida в полевых и лабораторных условиях. Проведенный анализ позволяет выделить среди Calanoida 3 группы по типу захвата пищи (табл. 7).
В постэмбриональном развитии H. hungaricus и H. rylovi последовательно сменяются несколько типов строения и функционирования ротовых аппаратов, сопровождающиеся сменой типа питания. В постэмбриональном развитии H. rylovi выявлено 3 способа питания: фильтрация (науплиус II – копеподит I), фильтрация + пассивный захват (копеподиты I–II), активный захват + фильтрация (копеподиты III–VI). В постэмбриональном развитии H. hungaricus выделе-
Таблица 7
Трофические группировки Calanoida
| Способ захвата пищи | Трофический уровень | Основная пища | Название вида |
| Фильтрация | II | Детрит, бактерио-, фитопланктон | E. vulgaris, A. denticornis, Arctodiaptomus sp. |
| Фильтрация и захват | II–III | Фито-зоопланктон | E. affinis, D. mirus, H. hungaricus, L. alluaudi |
| Активный захват и фильтрация | III | Зоопланктон и фитопланктон | H. rylovi |
но два способа питания: фильтрация (науплиус II – копеподит IV), фильтрация + пассивный захват (копеподиты IV–VI).
Трофическая структура зоопланктона временных водоемов определяется количеством и доступностью пищи, а также зависит от видовой структуры сообщества. Действие первого фактора обусловлено сезонной динамикой численности и биомассы популяций. У гидробионтов с продолжительным жизненным циклом два раза происходит закономерное изменение трофического статуса (табл. 8, 9). По нашим данным, экологическим стимулом к смене стратегии питания становится увеличение плотности популяций при высыхании водоема в конце весеннего периода. При этом заметно возрастает частота контактов рачков с потенциальными жертвами, поэтому становится эффективной стратегия факультативного или даже облигатного хищничества среди видов Anostraca и Calanoida.
Таблица 8
Сезонная динамика трофического статуса Anostraca

Условные обозначения: ![]() – фильтрация,
– фильтрация, ![]() – факультативное хищничество,
– факультативное хищничество, ![]() – хищничество
– хищничество
Разнообразия вариантов структуры популяций доминирующих видов и сообществ в целом, а также абиотических условий среды в различных типах временных водоемов определяют существенные отличия в трофической структуре зоопланктона. Основу видового состава в ранневесенний период составляют преимущественно активные хвататели. Однако основная тенденция в
Таблица 9
Сезонная динамика трофического статуса Calanoida

Условные обозначения см. табл. 8
формировании трофической структуры зоопланктона временных водоемов состоит в увеличение числа фильтраторов в течение вегетационного сезона.
Специфическими трофическими группировками зоопланктона временных водоемов можно считать фильтраторов-хватателей и хватателей-соскабливателей. Они присутствуют в ранневесенний и весенний период (за исключением L. alluaudi). Наибольшее число видов фильтраторов-хватателей обнаружено в весенний период. Оно обусловлено изменением трофического статуса особей в процессе постэмбрионального развития и вступлением в эту группу видов каланоид и весенне-раннелетних видов голых жаброногов достигших соответствующей стадии развития. В ранневесенний и весенний периоды основу трофической структуры (по численности) составляют собиратели-хвататели, а в летний период – фильтраторы.
В большинстве водоемов средний индивидуальный вес собирателей, собирателей-хватателей и хватателей в течение сезона изменяется незначительно. В весенний период фильтраторы включают Anostraca, крупных прудовых Cladocera и Calanoida, поэтому их средний индивидуальный вес значительно выше, чем в постоянных водоемах – 0.15–2.14 мг, как у фильтраторов-хватателей и хватателей-соскабливателей – 0.20–3.85 мг и 1.40–5.86 мг соответственно. Наибольший средний индивидуальный вес имеют хищные голые жаброноги из рода Branchinecta. Таким образом, ранневесенние и весенние виды Anostraca, Cladocera и Calanoida при низкой численности определяют формирование трофической структуры по биомассе.
В ранневесенний и весенний период биомассу зоопланктона формируют различные трофические группировки в зависимости от типа водоема, а в летний – фильтраторы.
Характер сезонной динамики разнообразия трофических группировок определяет присутствие фильтраторов-хватателей и хватателей-соскабливателей. Разнообразие трофических группировок по численности характеризуется весенним пиком, связанным с одновременным присутствием в это время видов из нескольких сезонных группировок, различающихся по пищевой специализации, но близких по численности. Разнообразие трофических группировок по биомассе убывает в течение сезона: ранней весной они многообразны, а их биомассы относительно выровнены; летом трофическая структура наименее разнообразна, поскольку фильтраторы-хвататели и хвататели-соскабливатели не участвуют в формировании сообществ, а фильтраторы доминируют по численности и биомассе.
Основные показатели трофической структуры зоопланктона находятся в зависимости от особенностей водоема. Наиболее значимы связи с продолжительностью существования водоема, например среднее число видов-фильтраторов и собирателей.
Анализ градиентных рядов водоемов по абиотическим факторам позволяет обнаружить закономерную смену преобладающих трофических группировок. Водоемы V группы находятся под значительным влиянием зоопланктона, поступающего из оросительных каналов, сформированного преимущественного из фильтраторов. Здесь выявлено наибольшее их число – 8 видов. Трофическая структура зоопланктона лиманов отличается наибольшим числом видов фильтраторов-хватателей и хватателей. Присутствие значительного числа видов фильтраторов-хватателей напрямую связано с формированием высокой биомассы зоопланктона, что подтверждается существованием значимой связи между этими показателями. В лужах число видов-фильтраторов минимально. Соотношение трофических группировок по биомассе отличается в водоемах различных групп (табл. 10).
Таблица 10
Средняя за сезон доля от общей биомассы трофических группировок
зоопланктона временных водоемов различных групп, %
| Трофические группировки | Группы водоемов | |||||
| III | IV | V | VI | VII | ||
| фильтрация | 16.79 (6.90-28.22) | 36.04 (0.30-70.74) | 77.35 (56.08-98.62) | 34.62 | 30.22 (1.93-91.63) | |
| собирание | 0.28 (0.02-0.94) | 0.10 (0-0.25) | 0.36 (0.19-0.52) | – | – | |
| фильтрация-захват | 62.35 (32.77-86.26) | 41.64 (11.02-97.52) | 0.13 (0.09-0.16) | – | 9.34 0-46.10) | |
| собирание-захват | 2.80 (0.59-9.66) | 5.92 (1.19-12.11) | 19.15 (0.69-37.60) | 51.71 | 34.90 (3.34-89.25) | |
| захват | 17.79 (2.27-55.49) | 9.27 (0.98-13.70) | 3.00 (0.37-5.63) | 5.99 | 25.53 (1.13-85.57) | |
| захват-соскабливание | – | 7.03 (0-21.08) | 0.02 (0-0.04) | 7.69 | – | |
Примечания: в скобках указан размах варьирования показателя
Доля фильтраторов наиболее велика в лиманах, связанных с оросительными системами. В водоемах III и IV групп выявлена высокая биомасса фильтраторов-хватателей. Они наиболее характерны для временных водоемов со средней продолжительностью существования. При этом в водоемах III группы фильтраторы-хвататели представлены преимущественно Calanoida, для развития которых важен значительный объем свободной от растительности водной массы; в лиманах с наименьшей площадью свободного водного зеркала фильтраторы-хвататели Calanoida малочисленны. Функцию фильтраторов-хватателей здесь выполняют Anostraca, поскольку зарастание водоема не препятствует развитию их популяций. В водоемах VII группы преобладают собиратели-хвататели.
ВЫВОДЫ
1. Видовой состав зоопланктона временных водоемов Саратовской области включает 68 видов низших ракообразных, в том числе: голых жаброногов – 9, ветвистоусых ракообразных – 25, каланоид – 10 и циклопоид – 24 видов. Число видов растет с северо-запада на юго-восток в ряду: временные водоемы лесостепи, степи и сухой степи (33, 39 и 52 вида). В долинах рек Правобережья значимы интразональные элементы фауны из северного и холодноводного комплексов умеренных широт, а в лиманах сухой степи – виды из фауны постоянных водоемов. Количество видов растет при увеличении продолжительности существования водоемов.
2. Выделено 7 групп временных водоемов, различающихся по видовому составу зоопланктона: водоемы поймы; водоемы надпойменных террас; водоемы водоразделов; лиманы; лиманы, связанные с оросительной системой; копаные пруды; лужи.
3. Голые жаброноги и каланоиды формируют по 3 сезонные группировки: голые жаброноги – ранневесеннюю, весеннюю и весенне-летнюю, а каланоиды – весеннюю, весенне-летнюю и летнюю. Популяции различных сезонных группировок каланоид отличаются по синхронности выхода науплиусов из диапаузы, размерно-возрастной и половой структуре, сезонной динамике численности и биомассы.
4. Размерно-возрастная структура популяций низших ракообразных зависит от температурного режима и морфометрических параметров водоема. Для массовых видов веслоногих рачков установлена количественная зависимость средней длины тела от средней температуры воды, при которой происходит развитие науплиусов.
5. В ранневесенний период зоопланктон имеет наименьшее число видов, численность, биомассу и видовое разнообразие. Численность определяют циклопоиды, а биомассу – каланоиды и голые жаброноги. В весенний период наблюдается наибольшее количество видов, числа структурообразующих видов и биомассы. Основу численности составляют циклопоиды, а биомассы – голые жаброноги, каланоиды и циклопоиды. В летний период количество видов уменьшается, но увеличивается видовое разнообразие; численность и биомассу сообщества формируют каланоиды и кладоцеры.
6. Продолжительность существования водоема оказывает наибольшее влияние на среднее за сезон количество видов Cladocera, число структурообразующих видов (по биомассе), среднее за сезон число видов и соотношение биомасс Cyclopoida/Calanoida. Биомасса зоопланктона наиболее велика в лиманах. Отношение Bmin/Bmax вариабельно в водоемах с искусственно регулируемым уровнем воды, более стабильно в лиманах.
7. По способу добывания пищи выделены основные трофические группы низших ракообразных и установлена принадлежность к ним отдельных видов. У голых жаброногов выделено пять групп: фильтраторы; переходная группа, сочетающая соскабливание, фильтрацию, захват; фильтраторы, способные к пассивному захвату; хвататели-фильтраторы; хвататели-соскабливатели. У Calanoida – три группы: фильтраторы; фильтраторы-хвататели; активные хвататели, способные к фильтрации.
8. Активные хвататели составляют основу сообщества в ранневесенний период. Фильтраторы-хвататели и хвататели-соскабливатели присутствуют в сообществе только в ранневесенний и весенний периоды. Собиратели-хвататели составляют основу трофической структуры (по численности) в ранневесенний и весенний периоды, фильтраторы – в летний период. В ранневесенний и весенний периоды биомассу зоопланктона в разных группах водоемов формируют различные трофические группировки, а в летний период – фильтраторы. Трофическое разнообразие (по биомассе) убывает в ходе сезонного развития зоопланктона. При увеличении продолжительности существования водоемов ведущая трофическая группа меняется в последовательности: собиратели-хвататели – фильтраторы-хвататели – фильтраторы. Доля фильтраторов по численности и биомассе наиболее значительна в лиманах, связанных с оросительными системами. Биомасса фильтраторов-хватателей велика в водоемах степных водоразделов и лиманах, собирателей-хватателей – в лужах.
СПИСОК ОСНОВНЫХ РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
1. Евдокимов Н.А. Жизненные циклы двух видов Hemidiaptomus Kiefer, 1933 (Copepoda, Calanoida) из временных водоемов Саратовского Заволжья // Биология внутренних вод: проблемы экологии и биоразнообразия. Тез. докл. XII Межд. конф. молодых ученых. Борок: ИБВВ РАН, 2002. С. 67–68
2. Евдокимов Н.А. Жизненные циклы двух видов Hemidiaptomus Kiefer, 1933 (Copepoda, Calanoida) из временных водоемов Саратовского Заволжья // Биология внутренних вод: проблемы экологии и биоразнообразия. Мат. XII Межд. конф. молодых ученых. Борок: ИБВВ РАН, 2002. С. 31–39.
3. Евдокимов Н.А. Особенности жизненного цикла Diaptomus mirus Lilljeborg, 1989 (Copepoda, Calanoida) во временных водоемах Саратовского Заволжья // Вопросы биологии, экологии, химии и методики обучения: Сб. научн. статей. Вып. 5. Саратов: Научная книга, 2002. С. 51–54.
4. Евдокимов Н.А., Ермохин М.В. Фауна голых жаброногов (Crustacea, Anostraca) Саратовской области // Охрана растительного и животного мира Поволжья и сопредельных территорий: Мат. Всерос. науч. конф. Пенза: Изд-во ПГПУ им. В.Г. Белинского, 2003 С. 200–203.
5. Евдокимов Н.А. Трофический статус каланоидных ракообразных (Copepoda, Calanoida) во временных водоемах Саратовской области // Трофические связи в водных сообществах и экосистемах: Мат. Межд. конф. Борок: ИБВВ РАН, 2003. С. 36.
6. Евдокимов Н.А. Функциональная морфология и трофический статус голых жаброногов (Crustacea, Anostraca) во временных водоемах Саратовской области // Трофические связи в водных сообществах и экосистемах: Мат. Межд. конф. Борок: ИБВВ РАН, 2003. С. 37.
7. Евдокимов Н.А., Ермохин М.В. Видовой состав личинок хирономид (Chironomidae, Diptera) в меропланктоне временных водоемов Саратовской области. // Энтомологические и паразитологические исследования в Поволжье. Вып. 4. Саратов: Изд-во Сарат. ун-та, 2005. С. 72–80.
8. Евдокимов Н.А. Фауна Calanoida временных водоемов Саратовской области // Биоресурсы и биоразнообразие экосистем Поволжья: прошлое, настоящее, будущее: Матер. межд. совещ. Саратов: Изд-во Сарат. ун-та, 2005. С. 276–279.
9. Ермохин М.В., Евдокимов Н.А. Редкие и исчезающие виды водных беспозвоночных на страницах Красной книги Саратовской области // Поволжск. экол. журн. 2006. Спец. вып. С. 14–23.
Подписано в печать 1.08.2006. Формат 60х84 1/16.
Бумага офсетная. Гарнитура Таймс New Roman Cyr. Печать офсетная.
Усл. печ. л. 1. Тираж 100 экз. Заказ 320.
Отпечатано в Центре полиграфических и копировальных услуг. Предприниматель Серман Ю.Б. Свидетельство №3117.4100600, Саратов, ул. Московская, 157, офис.