

Морфогенез лиственницы сибирской в связи с современным изменением климата в высокогорьях полярного урала
На правах рукописи

Дэви Надежда Михайловна
морфогенез лиственницы сибирской в связи с современным изменением климата в высокогорьях Полярного Урала
Специальность 03.00.16 – экология
Автореферат диссертации на соискание ученой степени
кандидата биологических наук
Пермь – 2008
Работа выполнена в Институте экологии растений и животных
Уральского отделения Российской Академии Наук
Научный руководитель: доктор биологических наук
Мазепа Валерий Семенович
Официальные оппоненты: доктор сельскохозяйственных наук
Нагимов Зуфар Ягфарович
кандидат биологических наук, доцент
Малеев Кирилл Иванович
Ведущая организация: Институт леса им. В.Н. Сукачева
СО РАН
Защита состоится 23 октября 2008 г. в 16.00 часов на заседании Диссертационного совета
Д 212.189.02 при Пермском государственном университете: 614990, г. Пермь, ул. Букирева, 15; факс (342) 2371611;
e-mail: [email protected]
С диссертацией можно ознакомиться в библиотеке Пермского государственного университета.
Автореферат разослан « 15 » сентября 2008 г.
Ученый секретарь
Диссертационного совета
доктор биологических наук, доцент Новоселова Л.В.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. В настоящее время возникла необходимость проведения систематических научных работ, направленных на изучение механизмов адаптации и устойчивости отдельных организмов, их сообществ и экосистем в целом к постоянно меняющимся условиям среды.
В числе важнейших задач, направленных на решение проблемы реакции экосистем на глобальные и региональные изменения климата, является исследование северных регионов. Они в первую очередь реагируют на глобальное потепление, так как климатическая обусловленность реакций экосистем проявляется здесь наиболее четко (Kelly et al., 1982; Vaganov et al., 1999; Шиятов, Мазепа, 2007). Для синэкологов лесные сообщества северных широт с их предельно низким уровнем биоразнообразия и упрощенной структурой являются наиболее простой моделью экосистем в изучении принципиальных особенностей и механизмов экологических явлений (Krner 1999; Influence…, 1999; Kittel et al, 2000; Global change…, 2001).
В настоящее время накоплен большой опыт исследований по экологической морфологии растений, в частности создано несколько учений о жизненных формах (Warming, 1908, Raunkiaer, 1937; Серебряков, 1962). Однако вопросы климатогенного формообразования у растений до настоящего момента изучены недостаточно.
Актуальной остается и оценка углерододепонирующей способности растительных сообществ на фоне современных изменений климата (Усольцев, Залесов, 2005).
Цель исследования – выявление климатогенных стратегий структурной адаптации деревьев лиственницы сибирской, а также оценка их биологической продуктивности в высокогорьях Полярного Урала.
Задачи исследования:
1. Описать многообразие жизненных форм деревьев лиственницы сибирской и оценить спектр экоморф в зависимости от условий произрастания.
2. Определить календарные периоды фаз морфогенеза у деревьев с многоствольной формой роста и сопоставить их с динамикой изменения климата.
3. Проанализировать ростовые характеристики разных жизненных форм лиственницы.
4. Оценить влияние климатических факторов на радиальный прирост различных жизненных форм лиственницы.
5. Произвести расчет накопления углерода и его распределения в надземной и подземной фитомассе деревьев лиственницы сибирской в экотоне верхней границы леса.
Основные положения, выносимые на защиту:
1. На Полярном Урале в экотоне верхней границы леса представлены 4 основные экоморфы лиственницы сибирской: ковровидная, стланиковая, многоствольная и стволовая.
2. В течение последних 100–120 лет произошли существенные изменения в спектре основных экоморф лиственницы сибирской. Эти трансформации календарно сдатированы и являются новым биоиндикатором изменения климата.
3. Скорость радиального и линейного прироста статистически отличаются у различных жизненных форм лиственницы. При смене жизненной формы с стланиковой на многоствольную, обусловленной улучшением условий существования, скорость роста многократно увеличивается.
4. Фракционный состав фитомассы различных жизненных форм лиственницы показывает высокую степень специфичности.
Научная новизна.
- Показано, что на Полярном Урале в экотоне верхней границы леса в XX в. произошел массовый переход стланиковой жизненной формы лиственницы в многоствольную. В современных древостоях наблюдается преобладание стволовой формы даже на сильно ветрообдуваемых местообитаниях. Соотношение различных форм роста деревьев служит надежным индикатором изменения климата.
- Впервые проанализирована изменчивость радиального и линейного прироста различных форм роста лиственницы.
- Показана специфичность влияния основных климатических параметров на прирост лиственницы сибирской в зависимости от жизненной формы дерева.
- Даны оценки накопления биомассы разными жизненными формами лиственницы и произведены расчеты запасов углерода древесного яруса для различных типов лесотундровых сообществ.
Практическая значимость работы.
Полученные результаты могут быть использованы для индикации климатических изменений и для оценки продуктивности, в первую очередь потока и запаса углерода в лесотундровых экосистемах. Материалы диссертационной работы могут быть использованы при чтении лекций по экологии и морфологии растений, дендрологии и дендрохронологии.
Апробация результатов и публикации по теме диссертации.
Результаты исследований выносились на обсуждение на 6-и молодежных конференциях: «Современные проблемы популяционной, исторической и прикладной экологии» (Екатеринбург, 2001), «Биота горных территорий: история и современное состояние» (Екатеринбург, 2002), «Проблемы глобальной и региональной экологии» (Екатеринбург, 2003), «Экологические механизмы динамики и устойчивости биоты» (Екатеринбург, 2004), «Экология: от генов до экосистем» (Екатеринбург, 2005), «Экология от Арктики до Антарктики» (Екатеринбург, 2007), на 5-и Международных конференциях: «Экологические проблемы горных территорий» (Екатеринбург, 2002), «Древесные кольца в археологии, климатологии и экологии» (Фрибур, Швейцария, 2005; Терверен, Бельгия, 2006), «Влияние изменений климата на бореальные и умеренные леса» (Екатеринбург, 2006), «Криогенные ресурсы полярных регионов» (Салехард, 2007), всероссийской конференции «Новые методы в дендроэкологии» (Иркутск, 2007) и Русско-Британском симпозиуме: «Изменение климата, отклик прироста деревьев и реконструкция климата» (Красноярск, 2006).
По теме диссертации было опубликовано 13 работ: 2 статьи, 7 кратких сообщений и 4 тезисов.
Структура и объем диссертации. Диссертация состоит из введения, восьми глав, выводов, списка литературы и приложений. Работа изложена на 152 страницах (из них 139 страниц машинописного текста), содержит 10 таблиц и 31 рисунок. Список литературы включает 154 работы, в числе которых 35 на иностранном языке.
Содержание работы
Введение. Сформулированы актуальность работы, ее значимость, цель, задачи и основные положения, выносимые на защиту.
Глава 1. Внутривидовая изменчивость и жизненная форма растений. Лиственница сибирская
1.1. Внутривидовая изменчивость растений
Исследованием внутривидовой изменчивости растений посвящены труды: М. Адансона (1763), Г. Гордона (1880), Г. Хеги (1908), Г. Хартвига (1834), Е. Цедербауэра (1907), Л. Кляйна (1908), В.В. Станчинского (1927), Ю.А. Филипченко (1927), В.С. Кирпичникова (1935), В.Н. Сукачева (1938), Л.Ф. Правдина (1958), С.А. Мамаева (1973) и многих других ученых.
1.2. Жизненная форма растений
В настоящее время имеется множество подходов к определению понятия “жизненная форма” (Варминг, 1884; Друде, 1913; Раункиер, 1937; Келлер, 1948; Гамс, 1950; Алехин, 1950; Шенников, 1950; Браун-Бланке, 1951; Культиасов, 1953; Григорьев, 1954; Коровин, 1961; Зозулин, 1968; Хохряков, 1981), вызванных различным объемом его трактовки и особенностями задач, которые ставят перед собой те или иные исследователи. Современные авторы (Серебряков, 1962; Жмылев и др., 2005; Кирпотин, 2006; Волков, 2006) подразумевают под жизненной формой - общий облик (габитус растения), обусловленный своеобразием его системы надземных и подземных вегетативных органов, формирующихся в онтогенезе в результате роста и развития растения в определенных условиях среды.
1.3. Эколого-биологические и морфологические особенности рода лиственница
1.4. Лиственница сибирская (Larix sibirica Ledeb.)
Многочисленные исследования показали что лиственница сибирская (Larix sibirica Ledeb.) обладает рядом эколого-биологических и морфологических особенностей (Тимофеев, 1961; Дылис, 1981; Коропачинский, Встовская, 2002), в числе которых – способность образовывать различные жизненные формы, позволяющих ей существовать в экстремальных климатических и почвенно-грунтовых условиях, которые характерны для высокогорьев Полярного Урала (Шиятов, 1966; 1967; Рождественский, 1984; Горчаковский, Шиятов, 1985; Мазепа, Дэви, 2007; Devi et al., 2008).
Глава 2. Природно-климатические условия района исследования
Исследования проводились на территории Ямало-Ненецкого автономного округа Тюменской области на восточном макросклоне Полярного Урала в бассейне реки Соби (6646–6655 с.ш., 6522–6549 в.д.). В подглавах 2.1.–2.7. приведены сведения по геологии, гидрографии, почвам, климату и растительности района исследования.
Глава 3. Объекты и методы исследования
Объектом исследования была лиственница сибирская, произрастающая в виде различных жизненных форм в пределах экотона верхней границы леса – в полосе между верхней границей распространения отдельных деревьев в тундре и верхней границей распространения сомкнутых лесов. Для оценки состава и структуры древесного яруса лиственничных редколесий и лесов использовались 2 стационарных высотных профиля и 3 дополнительные пробные площади (рис. 1). Геоботаническое описание проводилось по общепринятой методике (Полевая…, 1976).

Рис. 1. Расположение профилей и пробных площадей (1-профиль I, 2-профиль II, 3- пробные площади). Масштаб 1:25000
Профиль I имеет восточную экспозицию. В верхней части профиля (298-300м н.у.м.) растительное сообщество представлено тундрой кустарничково-мохово-лишайниковой с ерником, где единично встречаются деревья лиственницы сибирской. Для средней части склона (220 м н.у.м) характерно лиственничное редколесье ерниково-травяно-кустарничково-моховое. На выровненной поверхности в нижней части склона (185 м н.у.м.) представлен лиственничный лес ерниково-травяно-кустарничково-моховой.
Профиль-II заложен на северо-восточном склоне. Первый высотный уровень располагается в верхней части склона морены (265-268 м н.у.м), каменистая поверхность которого покрыта травяно-кустарничковой со мхами и лишайниками тундрой. Встречаются единичные деревья лиственницы одноствольной и многоствольной форм роста. Второй высотный уровень занимает среднюю часть склона (240 м. н.у.м). Сообщество представлено лиственничным редколесьем ерниково-кустарничково-мохово-лишайниковым. Третий уровень расположен в нижней части морены (220 м. н.у.м ), где характерен лиственничник ерниково-травяной с подлеском из ивы и можжевельника.
Пробная площадь 1 заложена на высоте 235 м над уровнем моря в лиственничном редколесье. Пробная площадь 2 заложена на высоте 197 м н.у.м. в лиственничном лесу. Пробная площадь 3 заложена на высоте 240 м н.у.м., где представлена тундровая растительность с единичными деревьями лиственницы.
Объем материала, использованного для решения каждой из поставленных задач.
Задача 1. Учет многообразия жизненных форм деревьев лиственницы сибирской произведен маршрутным методом на территории около 40 км2 у подножия гор Черная и Малая Черная.
Для оценки спектра экоморф в зависимости от условий произрастания использовался полный перечет всех живых деревьев с характеристикой морфометрии на 144 пробных площадках, размером 10х10м, общей площадью 1,44 га. Общее число учтенных деревьев на пробных площадках составило 607 шт. Из них одноствольных 512 шт., многоствольных 95 шт.
Задача 2. Для выявления календарных периодов фаз морфогенеза у деревьев с многоствольной формой роста использовано 23 особи. Даты появления особей определены дендрохронологическим методом (Douglass, 1919; Fritts, 1976; Шиятов, 1973) по годичным кольцам у 37 спилов, взятых у основания гипокотиля. Даты формирования вертикальных стволов у этих особей определены по 132 спилам со стволов, взятых в месте перехода от плагиотропного к ортотропному росту.
Задача 3. Для анализа изменчивости радиального и линейного прироста у особей с одноствольной (21 особь), многоствольной (18 особей) и стланиковой (4 особи) формой роста использовались спилы стволов и ветвей. Общее число спилов составило 677 шт. Общее число измерений годичного радиального прироста составило 21635 шт.
Задача 4. Оценка влияния климатических факторов на радиальный прирост различных жизненных форм производилась на основе 3 обобщенных стандартизированных рядов прироста, для построения которых использовано 66 индивидуальных рядов (15 для одноствольных, 17 для стлаников и 34 для многоствольных деревьев). Продолжительность обобщенных стандартизированных рядов составила: для одноствольной жизненной формы 290 лет, для стланиковой и многоствольной 121 и 108 лет соответственно. В качестве климатических параметров использовался ряд инструментальных метеонаблюдений по метеостанции Салехард (WMO # 2012333000, 66°31' с.ш., 66°36' в.д., 35 м.н.у.м., с 1883 по 2004 гг.).
Задача 5. Расчет накопления углерода и его распределения в надземной и подземной фитомассе древесного яруса произведен с использованием аллометрических зависимостей между диаметром у основания ствола (для одноствольных деревьев и стлаников) или суммой площадей сечения основания всех стволов в куртине (для многоствольных деревьев) и весом абсолютно сухого вещества каждой фракции фитомассы. Аллометрические зависимости построены на основе оценки продуктивности у 21 одноствольной, 10 многоствольных и 3 стланиковых моделей лиственницы сибирской. Расчет абсолютно сухого вещества древесины стволов, ветвей и корней произведен на основе весовых пропорций 576 спилов и навесок. Абсолютно сухое вещество хвои рассчитывалось на основе весовых пропорций 50 навесок. Абсолютно сухая масса всех фракций была переведена в массу углерода по переводному коэффициенту К.И. Кобак (1988), равному 0,5 для древесных частей и 0,45 для хвои.
Статистическая обработка результатов исследований проведена с использованием пакета программ STATISTICA 6.0.
Глава 4. Жизненные формы лиственницы сибирской в экотоне верхней границы леса
4.1. Различия в условиях произрастания
Основными факторами, определяющими морфогенез лиственницы на Полярном Урале, являются тепловой и ветровой режимы, а также мощность снегового покрова (Горчаковский, Шиятов, 1985), поэтому оценка спектра экоморф лиственницы сибирской была проведена на основе материала, собранного маршрутным методом в местообитаниях, различающихся по условиям снегонакопления, типу увлажнения и величине ветровой нагрузки. Основные типы местообитаний были разделены на 3 группы: повышенные гривы и вершины морен, где преобладают постоянно сухие местообитания; средняя часть склонов, где преобладают переменно увлажненные и влажные местообитания; нижняя часть слабо дренированных склонов, характеризующаяся обильным и избыточным увлажнением и слабой ветровой нагрузкой.
4.2. Основные экоморфы лиственницы сибирской
Из всего многообразия наблюдаемых форм роста во время маршрутных исследований было выделено четыре основные экоморфы лиственницы сибирской.
Ковровидная форма (рис. 2а) является приспособлением к самым суровым условиям роста и приурочена к экстремально ветрообдуваемым местообитаниям, в которых деревья могут существовать только у самой поверхности земли, где условия наиболее благоприятны. Встречается единично, обычно за естественными укрытиями: буграми, кочками, камнями. Эта жизненная форма защищена от суровых зимних условий лишь частично, так как снежный покров в таких местообитаниях тонкий и часто пятнистый. Высота особей не превышает 10-15 сантиметров.
При погружении ветвей в верхний слой органического горизонта, обладающего повышенной влажностью по сравнению с влажностью на поверхности почвы, на ветвях формируются придаточные корни и они укореняются. За счет укоренения ветвей и разрастания с подветренной стороны, особи достигают значительного диаметра – до 2 м.
Стланиковая форма (рис. 2б) встречается на ветрообдуваемых местообитаниях (вершинах морен) и склонах, где происходит повреждение верхушек побегов ветром или переносимым снегом. Стланик представляет собой подушку из густо переплетенных, прижатых к земле ветвей. Высота стлаников обычно равна средней высоте снежного покрова и не превышает 0.5-1 м., так как климатические факторы вызывают повреждение терминальных побегов, которые превосходят уровень снега. Проекция кроны таких особей достигает 3-4 метров.
Многоствольное дерево представляет собой куртину, образованную одной особью и состоящую из нескольких вертикальных стволиков (рис. 2в). Количество стволов варьирует от двух до более чем двух десятков. Многоствольные деревья встречаются во всех типах местообитания, но преобладают на частично защищенных от ветра, умеренно увлажненных склонах.
У стволовой формы хорошо выражена одна главная ось первого порядка – прямой, но сильно сбежистый ствол (рис. 2г). Эта форма роста является доминирующей во всех типах местообитания.
А Б В Г



Рис. 2. Основные экоморфы лиственницы сибирской (А – ковровидная, Б - стланиковая,
В – многоствольная, Г – стволовая).
Особи могут совмещать габитуальные признаки, присущие той или иной выделенной форме в зависимости от конкретных условий произрастания, а при продолжительном и направленном изменении условий существования возможен переход одной жизненной формы в другую.
4.3. Морфогенез лиственницы сибирской как индикатор изменения климата
Согласно данным метеостанции Салехард, на Полярном Урале в ХХ веке произошли существенное потепление и увлажнение климата, которые продолжаются до настоящего времени (рис.3.). Средняя температура летних месяцев (июня–августа) в 1883–1920 гг. составляла 10.7C, в 1920–2004 гг. – +11.4C, т.е. возросла на 0.7C, а зимних месяцев (ноябрь–март) возросла на 1.1C (с -20.8 до -19.7С). Количество выпавших осадков в летние месяцы увеличилось на 32 мм (с 147 до 179 мм), а зимних - на 46 мм (с 67 до 113 мм). Так же отмечается более ранний срок начала вегетационного периода, о чем свидетельствует значительное повышение температуры мая. Если в 1883–1920 гг. она составляла -2.4C, то в 1920–2004 гг. повысилась до -1.1C, т.е. возросла на 1.3C.
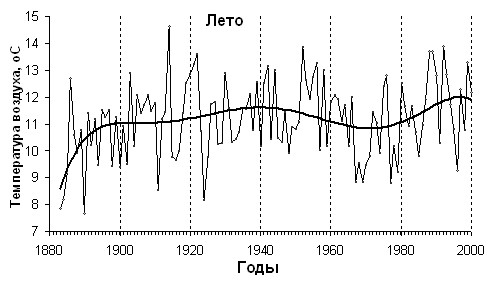



Рис. 3. Температура воздуха и сумма осадков летнего и зимнего периодов по метеостанции Салехард за последние 120 лет.
В исследуемом районе наблюдается большое количество деревьев с многоствольной формой роста, образовавшихся в результате формирования вертикальных стволиков у стланиковой формы. На рис. 4 видно, что время появления особей различно: самые старые деревья появились в начале XV столетия, самые молодые – в конце XIX столетия. Даты появления остальных особей распределились более или менее равномерно.
Даты формирования вертикальных стволов у модельных деревьев распределились иначе. Самые старые вертикальные стволы, живые в настоящее время, единично сформировались в XVII-XVIII столетиях. Некоторое увеличение частоты формирования вертикальных стволов наступило в XIX веке, особенно в его конце. Самая большая частота формирования вертикальных стволов наблюдается в начале ХХ века. После этого частота формирования вертикальных стволов постепенно снижается.

Рис.4. Время появления деревьев лиственницы сибирской и формирования вертикальных стволов.
В это же время началось усыхание части стволов у многоствольных деревьев (рис. 5). Максимум усыхания стволов пришелся на 70-80 годы ХХ века в результате усиления конкуренции за свет и питательные вещества между особями, в связи со значительным увеличением их размеров (Мазепа, Дэви, 2007).
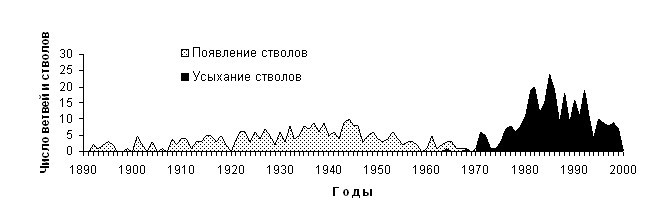
Рис. 5. Время появления и усыхания стволов у деревьев лиственницы с многоствольной формой роста.
Для выявления дендроклиматических связей был проведен анализ влияния термического режима и режима увлажнения летнего и зимнего периодов на частоту дат начала роста деревьев и смены жизненных форм (табл. 1). Анализ не выявил значимой связи между частотой дат начала роста особей всех жизненных форм и средними летними и зимними температурами. Однако выявлена прямая статистическая связь частот появления одноствольных деревьев с динамикой сумм осадков зимних (r= 0.77) и летних месяцев
(r= 0.71). Это согласуется с данными других авторов, что главнейшим лимитирующим фактором для появления проростков лиственницы является влажность субстрата в первые годы жизни растения (Шиятов, 1967; Санников, 1992).
Значимых связей между частотой появления стлаников и динамикой условий увлажнения не выявлено. Однако выявлена прямая статистическая связь между динамикой средних зимних температур, суммой осадков зимнего и летнего периодов и частотой лет, в которые произошло изменение формы роста деревьев с стланиковой на многоствольную.
Таблица 1.
Коэффициенты корреляции (r) между частотой дат появления и смены жизненных форм деревьев лиственницы сибирской и климатическими переменными летних и зимних месяцев Данные сглажены по 10-летиям.
| Появление деревьев (n=12) | Смена жизненной формы с стланиковой на многоствольную (n=4) | ||
| Климатическая переменная | Одноствольные деревья | Многоствольные | |
| Температура ноябрь-март | 0,14 | 0,23 | 0,81* |
| Температура июнь - август | 0,42 | 0,11 | 0,55 |
| Осадки ноябрь-март | 0,77** | 0,49 | 0,87* |
| Осадки июнь - август | 0,71** | 0,48 | 0,99*** |
*P<0.05; **P<0.01; ***P<0.001.
Это свидетельствует о том, что полученное распределение дат формирования вертикальных стволов у стлаников, независимо от времени появления особи, неслучайно и обусловлено улучшением климатических условий в начале ХХ века (Шиятов, Мазепа, 2002; Mazepa, 2005; Мазепа, Дэви, 2007, Devi et al., 2008).
Глава 5. Современное состояние и возрастная структура лиственничных древостоев в экотоне верхней границы леса
На Полярном Урале в экотоне верхней границы леса древостои характеризуются высокой морфологической обособленностью отдельных групп деревьев. Это обусловлено высокой гетерогенностью условий произрастания. Особенно резкие отличия в морфоструктуре древостоев отмечаются при движении вдоль высотного градиента: от тундры с одиночными деревьями к сомкнутому лесу (табл. 2).
5.1. Современное состояние древостоев
При сопоставлении характеристик деревьев на профиле I, отмечается постепенное увеличение основных морфометрических параметров деревьев в среднем в 2-3 раза при движении вниз по склону. Густота древостоя увеличивается в 26 раз. Площадки с большей долей многоствольных деревьев отмечаются в верхней части профиля. В остальных частях профиля эта жизненная форма лиственницы встречается в незначительном количестве (около 7%).
На северо-восточном склоне (профиль II), отмечается увеличение высоты и диаметра стволов в 5-8 раз по мере снижения высоты над уровнем моря. При этом густота древостоя возрастает в 2,5 раза. На профиле отмечается значительная доля многоствольных деревьев на всех высотных уровнях.
Анализ данных, полученных на пробных площадях, показал, что при переходе от тундры к сомкнутому лесу морфометрические параметры деревьев возрастают в 2-3 раза. Количество деревьев на единицу площади возрастает в 22 раза. Площадки с большей долей многоствольных деревьев расположены в тундре с одиночными деревьями и их доля постепенно снижается по мере перехода к сомкнутому лесу.
5.2. Возрастная структура
В полосе экотона верхней границы леса произрастают деревья трех основных поколений: молодое, средневозрастное и перестойное (рис. 6). Они сформировались в 1910-1990, 1830-1890 и 1650-1810 годах соответственно. Разрыв между поколениями составляет 20-50 лет.
К началу XX столетия на всех профилях произрастало лишь треть из ныне живущих деревьев, причем большинство из них в нижней части профилей. Из этого количества деревьев чуть больше трети составляли деревья с многоствольной формой роста. Примерно 20% из них произрастали на участках современных редколесий. Остальные деревья – на участках, где в настоящее время представлены сомкнутые леса.
Таблица 2.
Основные гипсометрические и таксационные характеристики высотных профилей и пробных площадей
| Характеристика | Склон | Тундра с одиночными деревьями | Редколесье | Сомкнутый лес |
| Высота над уровнем моря, м | В | 299 | 220 | 183 |
| С-В | 265 | 240 | 220 | |
| Ю-В | 265 | 235 | 197 | |
| Количество площадок (10х10 м), шт. | В | 12 | 12 | 12 |
| С-В | 12 | 12 | 12 | |
| Ю-В | 20 | 20 | 4 | |
| Густота древостоя, деревьев/га | В | 17 | 475 | 450 |
| С-В | 142 | 875 | 350 | |
| Ю-В | 105 | 565 | 2350 | |
| Процент много-ствольных деревьев, % | В | 100 | 4-10 | 3-11 |
| С-В | 9-29 | 11-17 | 22-36 | |
| Ю-В | 27-49 | 12-18 | 9-15 | |
| **Средний диаметр ствола, см | В | |||
| С-В | ||||
| Ю-В | ||||
| **Средняя высота ствола, м | В | |||
| С-В | ||||
| Ю-В | ||||
| **Средний диаметр кроны, м | В | |||
| С-В | ||||
| Ю-В |
*Численные значения средних морфометрических характеристик деревьев представлены дробью.
В числителе – одноствольные, в знаменателе – многоствольные деревья.
К настоящему времени преобладает молодое поколение, начало появления которого приурочено к 20-м и 70-м годам ХХ века. Это поколение представлено в основном стволовой формой роста.


Рис. 6. Возрастная структура различных древостоев в экотоне верхней границы леса.
Средний возраст одноствольных деревьев уменьшается с увеличением высоты над уровнем моря. В тундре с одиночными деревьями и редколесье 87% одноствольных деревьев появилось после 1970, при том что в сомкнутом лесу их средний возраст составляет 135 лет. Средний возраст многоствольных деревьев на всех высотных уровнях составляет 140 лет.
Глава 6. Изменчивость радиального и линейного прироста деревьев
Анализ индивидуальных рядов прироста показал различия между основными характеристиками древесно-кольцевых хронологий разных жизненных форм лиственницы (табл. 3).
6.1. Радиальный прирост
На рисунке 7 показана статистически значимая разница между средними значениями радиального прироста одноствольной и остальных жизненных форм (F2,63=10.693, p=0.00010). Средние приросты стланиковой и многоствольной форм лиственницы статистически не различаются (F1,49 = 0.43356, p= 0.51333).

Рис. 7. Средние значения радиального прироста и доверительные интервалы 3-х жизненных форм лиственницы сибирской.
Возрастные тренды наиболее выражены в хронологиях, построенных по одноствольным и стланиковым деревьям лиственницы, о чем свидетельствуют наиболее высокие значения автокорреляции, 0,73 и 0, 74 соответственно, по сравнению с хронологией, построенной по многоствольным деревьям (0,64).
Таблица 3.
Основные статистические характеристики индивидуальных древесно-кольцевых хронологий.
| Жизненная форма | Среднее значение радиального прироста, мм | Стандартное отклонение, мм | Максимальное значение радиального прироста, мм | Автокорреляция I-порядка | Среднее значение коэффициента чувствительности |
| Одноствольная | 0,62 | 0,41 | 3,58 | 0,73 | 0,46 |
| Многоствольная | 0,41 | 0,23 | 2,57 | 0,64 | 0,36 |
| Стланиковая | 0,4 | 0,26 | 1,68 | 0,74 | 0,35 |
Различия в значении коэффициента чувствительности (табл. 3) свидетельствует о различной выраженности климатического сигнала в хронологиях.
6.2. Относительный прирост
Для оценки качества дендроклиматических рядов использовались коэффициент чувствительности и стандартное отклонение рядов относительного прироста, в которых исключены различия в величине прироста, обусловленные индивидуальными и возрастными особенностями деревьев. Чувствительность оценивает величину межгодичной изменчивости прироста, а стандартное отклонение – амплитуду его изменчивости. Чем выше коэффициент чувствительности и стандартное отклонение, тем более сильный климатический сигнал содержится в хронологии.
Наибольшей чувствительностью (0,33) и амплитудой колебаний (30%) обладает хронология, построенная по одноствольным деревьям лиственницы. Хронологии, построенные по многоствольным деревьям и стланикам, характеризуются более низкими значениями коэффициента чувствительности и стандартного отклонения. Для многоствольных деревьев эти показатели равны 0,27 и 24%, для стлаников – 0,28 и 25%.
6.3. Линейный прирост
Среднее значение линейного прироста (рис. 8) составляет 3,8 см/год для одноствольных деревьев, 1,99 см/год - для многоствольных и 0,79 см/год для стлаников. Полученные различия значимы статистически (F2,14=331.64, p=0.0001).
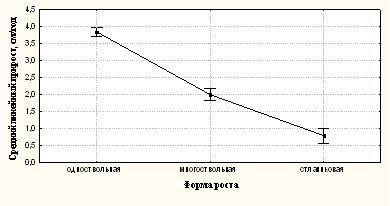
Рис.8. Средние значения линейного прироста и доверительные интервалы у 3-х жизненных форм лиственницы.
Отмечено резкое увеличение радиального и линейного прироста, произошедшее после 1920-х годов (рис.9).Средний линейный прирост после 1920 года увеличился в 1,87 раза у одноствольных деревьев и в 4,3 и 4,8 раза у многоствольных деревьев и стлаников. Значения радиального прироста возросли в 1,5, 2,6 и 2,1 раза соответственно. Это является косвенным доказательством улучшения условий существования для лиственницы в начале ХХ века на Полярном Урале.

Рис. 9. Средние значения радиального и линейного прироста лиственницы до и после 1920 года.
Глава 7. Влияние климатических факторов на динамику радиального прироста деревьев
На основе анализа функций отклика (Fritts, 1976) оценена климатически обусловленная изменчивость радиального прироста одноствольных и многоствольных деревьев и стлаников лиственницы сибирской. Оценка климатических функций отклика производилась за последние 125 лет - период, обеспеченный климатическими данными по метеостанции Салехард.
7.1. Температура воздуха
На изменчивость индексов радиального прироста деревьев положительно и значимо влияет температура летних месяцев (июня и июля). Для многоствольных и стланиковых деревьев статистически значимым оказался вклад температуры апреля. Однако этот вклад незначителен по величине (-0,2). Для одноствольных деревьев процент дисперсии хронологий, объяснимый изменчивостью средних месячных температур воздуха, составил
42 %. Для многоствольных деревьев и стлаников коэффициент составил 37% и 44% соответственно.
7.2 Осадки
Анализ функций отклика показал отсутствие значимого вклада осадков в прирост лиственницы, что свидетельствует о том, что количество осадков во все сезоны года не является значимым экологическим фактором для радиального прироста лиственницы на Полярном Урале.
Глава 8. Накопление и распределение фитомассы древесного яруса в экотоне верхней границы леса на Полярном Урале
8.1. Фракционная структура фитомассы лиственницы сибирской
Во фракционной структуре фитомассы лиственницы доминирует надземная часть, составляя 64% от общей фитомассы у одноствольных деревьев, 66% и 82 % у стлаников и многоствольных деревьев соответственно.
У одноствольных и многоствольных деревьев в общей фитомассе преобладают стволы, на их долю приходится 48% и 63% соответственно. Доля стволов в фитомассе стлаников незначительна и составляет 4%. У стлаников преобладающее значение в фитомассе имеют ветви. На их долю приходится 43% от общей фитомассы.
Доля фотосинтетического аппарата (ветви плюс хвоя) у многоствольных и одноствольных деревьев составляет 16% и 19% от общей фитомассы соответственно. У стлаников на фотосинтетический аппарат приходится в среднем 62% от общей фитомассы.
Корневой индекс (масса подземных органов/масса растения) у одноствольных деревьев наибольший и составляет 36%. Близкое значение фитомассы корней имеют стланики – 34%. У многоствольных деревьев этот показатель значительно ниже – 18%. Это объясняется тем, что многоствольные деревья нуждаются в меньшей корневой массе для стабилизации вертикального роста, так как растущие рядом стволы защищают друг друга от ветра и других неблагоприятных факторов.
8.2. Регрессионные уравнения фитомассы
Для модельных деревьев каждой жизненной формы были вычислены коэффициенты аллометрического уравнения связи массы различных фракций с единым, легко измеряемым морфометрическим показателем. У стлаников и одноствольных деревьев таким показателем является диаметр у основания ствола. Для многоствольных деревьев бралась сумма площадей сечения основания всех стволов в куртине. Полученные уравнения объясняют до 99,7% изменчивости фитомассы отдельных фракций.
8.3. Высотное изменение запасов фитомассы древостоев
Выявлено закономерное увеличение запаса общей фитомассы деревьев вниз по высотному градиенту. Суммарно для многоствольных и одноствольных деревьев увеличение общей фитомассы вниз по склону составило в среднем 733 раза. Общая фитомасса одноствольных деревьев возросла более чем в 1200 раз. Фитомасса многоствольных деревьев увеличилась в 260 раз.
8.4. Оценка запасов углерода в фитомассе лиственничных древостоев вдоль высотного градиента
Запас углерода в фитомассе древесины стволов (Сws), ветвей (Сwb), общей фитомассе корней (Сr) и хвои (Сn) на различных высотных уровнях представлен в табл. 4.
Таблица 4.
Распределение запасов углерода в фитомассе лиственницы сибирской по высотному градиенту.
| Высотный уровень | Сn, (т/га) | Сwb, (т/га) | Сws, (т/га) | Сr, (т/га) | |
| Профиль-I | Тундра с одиночными деревьями | 0,01 | 0,003 | 0,001 | 0,006 |
| Редколесье | 3,845 | 0,903 | 0,23 | 2,911 | |
| Сомкнутый лес | 8,954 | 2,029 | 0,426 | 6,919 | |
| Профиль-II | Тундра с одиночными деревьями | 0,062 | 0,019 | 0,009 | 0,038 |
| Редколесье | 1,438 | 0,431 | 0,142 | 0,783 | |
| Сомкнутый лес | 15,428 | 3,681 | 0,689 | 10,2 | |
| Пробные площади | Тундра с одиночными деревьями | 1,064 | 0,333 | 0,095 | 0,481 |
| Редколесье | 3,605 | 0,994 | 0,253 | 1,966 | |
| Сомкнутый лес | 15,192 | 3,849 | 1,003 | 10,008 | |
Выявлено закономерное повышение запасов углерода во всех фракциях фитомассы вниз по высотному градиенту. Для профилей разной экспозиции показатели возрастания запасов углерода различаются в десятки раз.
Принимая допущение, что отпад и гетеротрофное дыхание взаимно уравновешены (Швиденко и др., 2001), среднее изменение запасов углерода на каждом высотном уровне представлено как чистая экосистемная продукция (т/га в год) за период, равный среднему возрасту деревьев на данном уровне (табл. 5).
Таблица 5.
Чистая экосистемная продукция древостоев.
| Высотный уровень | Средний возраст, лет | Запас углерода в общей фитомассе, т/га | Экосистемная продукция, т/га в год | |
| Профиль-I | Тундра с одиночными деревьями | - * | 0,02 | -* |
| Редколесье | 53 | 7,9 | 0,15 | |
| Сомкнутый лес | 157 | 18,3 | 0,12 | |
| Профиль-II | Тундра с одиночными деревьями | 22 | 0,13 | 0,01 |
| Редколесье | 39 | 2,8 | 0,07 | |
| Сомкнутый лес | 190 | 29,9 | 0,16 | |
| Пробные площади | Тундра с одиночными деревьями | 86 | 1,9 | 0,02 |
| Редколесье | 60 | 6,8 | 0,11 | |
| Сомкнутый лес | 59 | 30,0 | 0,51 | |
*- не определялись
Анализ возрастной структуры древостоев показал, что более 70% всех живых растений на профилях появилось в течение ХХ века. Это говорит о том, что накопление существующего количества биомассы произошло на выбранных профилях, главным образом в ХХ веке. В лесу накопление углерода шло со скоростью 0,1 - 0,5 тонны углерода на гектар в год. В тундре и редине накапливалось в среднем по 0,07 тонны углерода на гектар в год. Эти показатели значительно ниже показателей, приводимых для других горных экотонов (Jansen, 2003), где накопление углерода в древесной фитомассе идет в среднем со скоростью 0,5 тонны углерода на гектар в год даже на верхних уровнях. Тем не менее, поскольку активная экспансия лесной растительности в тундру продолжается до сих пор, это может привести к значительному изменению бюджета углерода в данном регионе.
Выводы
- На Полярном Урале в экотоне верхней границы леса представлены четыре экоморфы лиственницы сибирской: ковровидная, стланиковая, многоствольная и стволовая, которые четко приурочены к определенным условиям произрастания. Резких границ между выделенными формами не существует.
- Улучшение климатических условий в первой половине XX столетия привело к массовому формированию деревьев с многоствольной жизненной формой, путем образования вертикальных стволиков у стланиковых жизненных форм. Интенсивный рост и значительное увеличение морфологических размеров средневозрастных деревьев с многоствольной формой роста в течение XX столетия привели к усыханию и гибели части стволов и ветвей в связи с усилением конкуренции за свет и питательные вещества между особями.
- Жизненные формы лиственницы сибирской характеризуются статистически различной скоростью радиального и линейного прироста. При улучшении условий существования скорость роста многократно увеличивается.
- Фракционный состав фитомассы различных жизненных форм лиственницы показывает высокую степень специфичности.
- При продвижении вниз по высотному градиенту отмечается закономерное увеличение запаса общей фитомассы деревьев и чистой экосистемной продукции древостоев, причем степень возрастания многократно отличается на склонах различной экспозиции.
Список работ, опубликованных по теме диссертации.
- в изданиях, рекомендованных ВАК:
1. Мазепа В.С. Образование многоствольных жизненных форм деревьев лиственницы сибирской в экотоне верхней границы леса на Полярном Урале как индикатор изменения климата / В.С. Мазепа, Н.М. Дэви. // Экология. 2007. № 6. C. 471-475.
2. Devi N. Expanding forests and changing growth forms of Siberian larch at the Polar Urals treeline during the 20th century/ N. Devi et al. // Global Change Biology. 2008. 14, 1-11. doi: 10.1111/j.1365-2486.2008.01583 x.
- в прочих изданиях:
3. Дэви H.М. Анализ изменчивости радиального прироста лиственницы сибирской на Полярном Урале / Н.М. Дэви. //Проблемы глобальной и региональной экологии: материалы конф. молодых ученых. Екатеринбург, 2003. C. 40-45.
4. Дэви Н.М. Оценка фракционного состава фитомассы деревьев лиственницы сибирской в экотоне верхней границы леса на Полярном Урале / Н.М. Дэви. // Экологические механизмы динамики и устойчивости биоты: материалы конф. молодых ученых. Екатеринбург, 2004. C. 65-68.
5. Devi N. Climate driven height growth change of the multistem trees of Larix Sibirica in the Polar Ural Mountains / N. Devi. // Tree Rings in Archaeology, Climatology and Ecology International conference. Department of Geosciences, Geography, University of Fribourg, Switzerland, 2005. P. 31.
6. Devi N.M. Climate driven height growth change of multistem trees of Siberian larch in the Polar Ural Mountains / N. Devi. // Transaction of the TRACE conference. Department of Geosciences, Geography, University of Fribourg, Switzerland, 2005. P. 140-143.
7. Дэви Н.М. Линейный и радиальный прирост многоствольных деревьев лиственницы сибирской под влиянием изменений климата в экотоне верхней границы леса на Полярном Урале / Н.М. Дэви. // Экология: от генов до экосистем: материалы конф. молодых ученых. Екатеринбург, 2005. С. 68-71.
8. Devi N. Climate driven growth form change of Siberian larch in the Polar Ural Mountains / N. Devi.// Abstracts of Workshop on Climate change, the tree growth response, and reconstruction of climate.V.N. Sukachev Institute of Forest SB RAS, Krasnoyarsk, Russia. P.33.
9. Devi N. Phenotypical modifications of Siberian larch in the ecotone of the upper forest bound at Polar Urals, Russia // Abstracts of the TRACE conference. Royal Museum for Central Africa & Ghent University, Belgium. P.36.
10. Devi N.M. Climate driven form genesis of Siberian larch at Polar Ural /N. Devi. // Abstracts of International Conference “Climate changes and their impact on Boreal and Temperate forests. P.23.
11. Дэви Н.М. Особенности радиального прироста основных жизненных форм лиственницы сибирской в районе массива Рай-Из: (Полярный Урал). / Н.М. Дэви. // Экология: от Арктики до Антарктики: материалы конф. молодых ученых. Екатеринбург, 2007. C. 81-86.
12. Дэви Н. Накопление биомассы в различных лесотундровых сообществах на Полярном Урале / Н. Дэви, В. Мазепа, Ф. Хагедорн.// Криогенные ресурсы Полярных регионов: мат. международ. конференции. Т. II. Объединенное науч-техн. изд-во Пущинского научного центра РАН, 2007. C. 72-74.
13. Дэви Н.М. Изменение экотона верхней границы леса в горах Полярного Урала в XX веке. / Н.М. Дэви, В. Мазепа. // Новые методы в дендроэкологии: материалы всерос. научн. конф.с международ. участием. Изд-во инст-та географии им. Сочавы СО РАН, 2007. C.178-180.
Подписано в печать 28.07.2008. Тираж 100 экз. Усл.п.л. 1,0.
Отпечатано в ООО «Таймер»
г. Екатеринбург, ул. Луначарского, 136. т.: (343)350-39-03