

Система и фауногенез коконопрядов мировой фауны ( lepidoptera : lasiocampidae )
На правах рукописи
ЗОЛОТУХИН Вадим Викторович
Система и фауногенез коконопрядов
мировой фауны (Lepidoptera: Lasiocampidae)
03.02.05 – энтомология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
доктора биологических наук
Санкт-Петербург, 2011
Работа выполнена на кафедре зоологии ФГБОУ ВПО
«Ульяновский государственный педагогический университет имени И. Н. Ульянова»
Научный консультант:
доктор биологических наук С.Ю.Синев (ЗИН РАН)
Официальные оппоненты:
доктор биологических наук В.А.Лухтанов (СПбГУ),
доктор биологических наук О.Г.Горбунов (ИПЭЭ РАН),
доктор биологических наук, профессор А.В.Селиховкин (СПбГЛТА)
Ведущая организация:
Саратовский государственный университет им. Н. Г. Чернышевского.
Защита состоится «13» октября 2011 в 16:00 часов на заседании совета Д 212.232.08 по защите докторских и кандидатских диссертаций при Санкт-Петербургском государственном университете по адресу: 199034, Санкт-Петербург, Университетская наб, д. 7/9, ауд. 133.
С диссертацией можно ознакомиться в научной библиотеке им. А. М. Горького Санкт-Петербургского государственного университета.
Автореферат разослан «……....» 2011 г.
Ученый секретарь
диссертационного совета,
кандидат биологических наук Е. Б. Ягунова
e-mail: [email protected]
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследований. Коконопряды (Lasiocampidae) – семейство шелкопрядообразных чешуекрылых (Bombycomorpha), иногда выделяемое в самостоятельное надсемейство Lasiocampoidea. Объем семейства до настоящего времени не выяснен; ежегодно описывается до 100 новых видов, в том числе и из таких хорошо исследованных регионов, как страны Юго-Восточной Азии или острова Тихого океана. В некоторых родах до 60% видов остаются неописанными, а до 20% используемых видовых эпитетов должны быть заменены старейшими валидными названиями. Общепринятой системы группы также не существует; разные авторы значительно расходятся в понимании объема и взаимосвязей между родами и обычно используют слабо обоснованную и уже устаревшую систему 1927 года, предложенную Х. Ауривиллиусом. Начатая ревизия коконопрядов мировой фауны показала, что большинство афро- и неотропических родов представляют собой сложные пара- или полифилетические комплексы, включающие несколько самостоятельных эволюционных ветвей. По-прежнему не разработаны вопросы фауногенеза коконопрядов, изучение которых может способствовать пониманию общих закономерностей эволюции группы.
Изучение коконопрядов актуально и в чисто практическом аспекте, поскольку они являются одними из основных дефолиаторов многих лиственных и хвойных пород. Кроме того, гусеницы Gloveria и Quadrina вызывают серьезные кожные раздражения, а пыль коконов Malacosoma аллергенна (Stehr, 1987). Некоторые виды коконопрядов используют для получения шелка, более прочного и долговечного, чем шелк тутового шелкопряда. Коконы некоторых видов в различных мировых культурах используются также для изготовления украшений и являются своеобразной валютой (Peigler, 1993, 1994), а куколки и гусеницы могут использоваться в пищу.
Цель и задачи исследования. Целью данной работы является изучение таксономической структуры семейства и вопросов его фауногенеза. Для этого были поставлены следующие задачи:
1) провести обзор существующих систем коконопрядов мировой фауны;
- на основе результатов сравнительно-морфологического, функционально-морфоло-гического и биохимического анализов модернизировать систему семейства в объеме мировой фауны;
- проанализировать видовой и родовой состав фаун коконопрядов всех континентов, установить уровень их эндемизма;
- определить вероятные пути фауногенеза группы на каждом из континентов;
- изучить видовой и родовой состав коконопрядов крупных островов Старого Света и выявить особенности становления островных фаун.
Основные положения, выносимые на защиту:
1. На основе анализа признаков внешней морфологии и генитального аппарата создана оригинальная классификация семейства Lasiocampidae, протестированная с помощью молекулярно-генетических методов. В составе семейства выделены пять крупных монофилетических групп, рассматриваемых в ранге подсемейств; самое крупное подсемейство Lasiocampinae Harris, 1841 включает 9 триб.
2. Основные события фауногенеза семейства приходятся на позднемеловой период и связаны с активными рельефообразовательными процессами этого времени. Специализированные эремиальные роды коконопрядов возникли относительно недавно, предположительно в плейстоцене, в связи с крайней аридизацией соответствующих территорий.
3. Фауны коконопрядов отдельных континентов слагаются как за счет видов автохтонного происхождения, так и за счет вселенцев. Неарктическая и неотропическая фауны коконопрядов формируются в результате 3 основных волн миграции предковых форм через Берингию из Евразии. Наиболее богаты эндемиками крупные океанические острова и континенты, на которых развитие местных фаун шло в условиях изоляции.
Научная новизна. Ревизована система семейства Lasiocampidae, ее основные положения опубликованы (Zolotuhin, 1998, 2009). В ходе проделанной работы определен таксономический состав семейства в рамках Старого Света; в частности, выявлен видовой состав ряда региональных фаун (Кавказа, европейской части бывшего СССР, Дальнего Востока России и Российской Федерации в целом, республик Средней Азии, Вьетнама, Филиппин, Таиланда, Сулавези, Флореса, Лаоса, Непала). Установлен видовой состав всех родов Палеарктической и Индо-Малайской фаун и частично ревизована Афротропическая фауна. Описаны в качестве новых для науки 139 видов и 43 подвида, а также 27 родов; подготовлены к описанию 2 новые трибы. Установлено более 100 новых синонимов видового и родового ранга.
Определено положение Lasiocampidae среди родственных семейств, и предложенная гипотеза проверена молекулярными методами. Предложены пути становления фаун различных материков и проанализирован фауногенез некоторых таксономических (Chionopsychinae, Chondrosteginae, Selenepherini и др.) и экологических (хвоегрызущие и эремиальные коконопряды) групп.
Теоретическая и практическая значимость. Результаты проведенной ревизии дают основу для последующих обобщений как в части построения надродовой системы семейства, так и определения его места в пределах бомбикоидного комплекса (Bombycoidea sensu lato). Разработка вопросов становления фаун коконопрядов различных материков вносит вклад в изучение исторического развития наземной биоты. Составленные региональные списки видов могут быть использованы службами лесного и сельского хозяйства различных стран для мониторинга вредных видов, а также природоохранными организациями для определения редких и требующих особого внимания видов. Переданные в Академию Наук Вьетнама списки хвоегрызущих видов (Zolotuhin, Ta, Tran, 2009) уже официально используются для корректировок сроков обработок сосновых посадок. Основные результаты и положения работы включены в курсы отдельных дисциплин, читаемых в УлГПУ, Самарском государственном университете и Саратовском государственном университете.
Апробация работы и публикации. Результаты работы докладывались на ежегодных конференциях преподавателей УлГПУ, Конгрессе по проблемам энтомологии Средней Европы (Жигулевск, 1993), X съезде Всесоюзного (Ленинград, 1989) и XIII съезде Русского энтомологического общества (Краснодар, 2007), XX международном энтомологическом конгрессе (Фиренце, 1996), XV Европейском лепидоптерологическом конгрессе (Эркнер, 2007), Зоологических чтениях Баварии (Мюнхен, 2007) и Конференции по биоресурсам (Ханой, 2009).
По теме диссертации опубликовано 84 работы, из которых 4 – монографии и коллективные монографии, 9 опубликованы в списке изданий, рекомендованных ВАК, из 71 прочих – 11 русскоязычные, 54 в иностранных изданиях и 6 как тезисы.
Структура и объем работы. Диссертация состоит из Введения, 7 глав, Выводов, Списка литературы и Приложения. Список цитируемой литературы включает 262 источника, среди которых 202 на иностранных языках. Основной текст диссертации изложен на 202 страницах, включает 23 таблицы и 34 рисунка. В Приложение вынесены база данных представителей семейства, отобранных для анализа ДНК; матрица нуклеотидных последовательностей фрагмента гена EF-1, использованных в работе; каталог видов коконопрядов мировой фауны и перечень выделов для кластерного анализа распространения коконопрядов.
Благодарности. Выражаю свою глубочайшую благодарность всем родным, друзьям и коллегам, без дружеской поддержки, разнообразной помощи и ценных советов которых данная работа была бы невозможной.
Глава 1. Материал и методика исследований
1.1. Материал. В основе работы лежат собственные сборы коконопрядов и близких групп чешуекрылых, выполненные в ходе полевых исследований в Европейской части России (1984–2008 гг.), Средней Азии (1989–1994), на Дальнем Востоке РФ, включая Сахалин и Кунашир (1991–1993), в Германии (1995), Турции (1995), Таиланде (2005), Вьетнаме (2006, 2008, 2009), ЮАР (2007), Демократической Республике Конго (2008). Обработаны материалы большинства ведущих научных учреждений мира и крупных частных коллекций России и Европы. За время исследований было обработано более 600 000 экземпляров и изготовлено более 5 900 препаратов различных морфологических структур, в том числе преимагинальных стадий – микроскульптуры хориона яиц, хетотаксии гусениц и куколок.
1.2. Методика. Методика сбора и выращивания живого материала была традиционной. Изготовление генитальных препаратов осуществлялось по общепринятой методике, с использованием в качестве уплотнителя Эупарала; инвертирование везики самца проводилось с помощью тонких инсулиновых шприцев. Электронно-микроскопические исследования выполнены на базе Zoologisches Forschungsinstitut und Museum Alexander Koenig, Bonn, с использованием сканирующего электронного микроскопа HITASHI S 450.
Материал для изучения ДНК фиксировался в 96% этаноле; дальнейшая его обработка проводилась на базе специализированных лабораторий (СарГУ и Canadian Centre for DNA Barcoding, Canada). Всего исследовано 2090 образцов.
Глава 2. Морфологический обзор семейства
Дан обзор морфологических признаков, служащих для целей диагностики и построения системы, рассмотренный по следующим разделам: Взрослое насекомое; Генитальный аппарат; Яйцо; Гусеница; Куколка и кокон. Выделено несколько диагностических черт семейства, часть которых являются его аутапоморфиями, а другие должны быть рассмотрены как гомоплазии: боковая или нижняя часть первого членика щупиков несет особую сенсорную структуру, напоминающая хетозему; пилиферы отсутствуют; в переднем крыле дистальная часть радиального ствола тесно сближена с субкостальным стволом; ареола отсутствует; CuA2 переднего крыла отходит от основания радиокубитальной ячейки на значительном удалении от основания М3; соединение крыльев осуществляется по амплексиформному механизму (Minet, 1998) с участием особой мелкошипиковой зоны переднего края заднего крыла, называемой спинареей; мускульный набор генитального аппарата самцов сокращен с исходных 8 до 2-3 пар мышц с полным выпадением m1, m3, m20 у большинства групп; хетотаксия гусениц первого возраста с обильным развитием подпервичных и вторичных щетинок; мандибулы гусениц с несколькими вторичными щетинками; брюшные ноги гусениц изнутри с двумя зонами отчетливой склеротизации, разделенными вертикальной мембраной; футляр хоботка у куколки сильно укорочен или полностью отсутствует.
Вместе с тем, семейство обладает несколькими явными плезиоморфиями: радио-кубитальная ячейка крыльев со следами медиального ствола; на крыльях часто сохраняется рудиментарная А1; гусеницы первого возраста несут полный или почти полный венец крючков на подошве брюшных ног; хетотаксия гусениц исходного типа без значимых модификаций; дендробионтность у подавляющего большинства родов; наличие эпикраниального шва у куколок; кокон (у архаичных родов) тесно связан с почвой и инкрустирован ее частицами.
Глава 3. Биология
В главе рассматриваются образ жизни коконопрядов, пищевые связи гусениц и их региональные особенности. Отсутствие доказанных случаев монофагии и редкость узкой олигофагии делают затруднительным анализ сопряженной эволюции коконопрядов и их кормовых растений. Можно лишь констатировать, что эволюция группы была преимущественно связана с освоением покрытосеменных, и прежде всего двудольных, растений. При проникновении в новые для них биотопы, коконопряды переходили к использованию и новых для них кормовых растений. В условиях аридного климата ими становились ксерофитные злаки и осоки или специализированные формы бобовых. В зависимости от условий обитания и исходных кормовых предпочтений развивалась полифагия на травянистых растениях (все Chondrostegini) или узкая олигофагия/монофагия на бобовых (Selenepherini аридных ландшафтов) или миртовых (австралийские представители). В горных местностях наиболее легко доступным кормовым субстратом для коконопрядов оказались голосеменные растения; приспособление к ним привело к формированию специализированных групп семейства, не способных использовать в пищу лиственные породы.
Глава 4. Система коконопрядов
4.1. Обзор имеющихся систем семейства. Самая ранняя попытка классифицировать коконопрядов была предпринята Дж. Таттом (Tutt, 1902), который рассмотрел представителей Палеарктической фауны в составе 3 отдельных семейств Chondrostegidae, Lachneidae и Eutrichidae, и предложил очень дробный вариант системы, включающей 23 трибы. Позднее Х. Ауривиллиус (Aurivillius, 1927), основываясь на особенностях расположения субкостальной жилки и радиального ствола на заднем крыле, предложил разделение семейства на 7 подсемейств: Chondrosteginae, Malacosomatinae, Archaeopachinae, Gastropachinae, Chionopsychinae, Lasiocampinae и Gonometinae. Все последующие предлагавшиеся варианты системы в той или иной степени опирались на схему, предложенную Ауривиллиусом.
4.2. Филогенетический анализ. Для тестирования выдвинутых ранее гипотез о взаимоотношениях различных родов был проведен филогенетический анализ семейства Lasiocampidae на основе изучения нуклеотидных последовательностей генов EF-1 и СО1 представителей его европейской, индо-малайской и афротропической фаун.
4.2.1. Объекты и методика исследования. Для 49 видов коконопрядов (64 экз.) с территории Евразии и Африки были установлены нуклеотидные последовательности гена EF-1. Также использованы 6 образцов, депонированных в GenBank. В качестве внешней группы выбран Lemonia dumi (семейство Lemoniidae).
Анализ нуклеотидных последовательностей проведен на автоматическом секвенаторе Beckman в лаборатории молекулярной биологии СарГУ. Коррекция полученных последовательностей выполнена в программе BioEdit Sequence Alignment Editor. Для множественного выравнивания использована программа ClustalW. Построение филогенетических деревьев произведено с использованиием методов максимального правдоподобия, ML, минимальной эволюции, ME, максимальной парсимонии, MP и Байеса, BI в пакетах прикладных программ MEGA 4, MEGA 5 и MrBayes 3.1.2.
Матрица нуклеотидных последовательностей составлена из фрагментов длиной от 585 до 1019 п.н. Всего в анализе были использованы 72 нуклеотидные последовательности. На исследуемом участке гена EF-1 с 164 по 1182 п.н. выявлено 345 вариабельных сайтов, что составило 34%. Доля парсимониально значимых сайтов составила 28% (287 сайтов). Среднее отношение транзиций к трансверсиям составило 2,57.
4.2.2. Исследование генетического родства представителей Lasiocampidae.
Топологии полученных кладограмм ML, ME, MP, и BI в целом оказались сходными и в основных узлах имели высокие значения поддержки (более 50% бутстреп-поддержки для методов ME, MP и ML и постериорная вероятность более 0.5 для BI). Метод Байеса дал высокие значения поддержки (0,99-1) для большинства клад (рис. 1), поэтому в качестве оптимального было выбрано именно дерево BI. По результатам проведенного анализа можно выделить несколько четко очерченных групп коконопрядов.
Две ветви, представленные родами Malacosoma и Chionopsyche, во всех вариантах дендрограмм оказываются базальными и обособленными, что позволяет рассматриватьотреть их в качестве отдельных подсемейств Malacosominae и Chionopsychinae. Остальные роды образуют особую кладу с высоким уровнем поддержки (66-98%), для которой существует валидное название Lasiocampinae. Все подчиненные ей крупные клады можно рассматривать в ранге триб Selenepherini, Gastropachini, Pinarini и Argudini tribus nova (Arguda + Syrastrena (+ Hallicarnia).
Род Euthrix представляется на настоящий момент парафилетическим. Вероятно, в дальнейшем будет целесообразно выделить вид E. isocyma в отдельный монотипический род.
Группа родов, объединяемая здесь в составе Argudini (Arguda, Syrastrena, Odontocraspis и Leipoxais), ранее сближалась с Odonestis в рамках трибы Odonestini. Однако все построенные дендрограммы широко разводят эти роды, которые, вероятно,
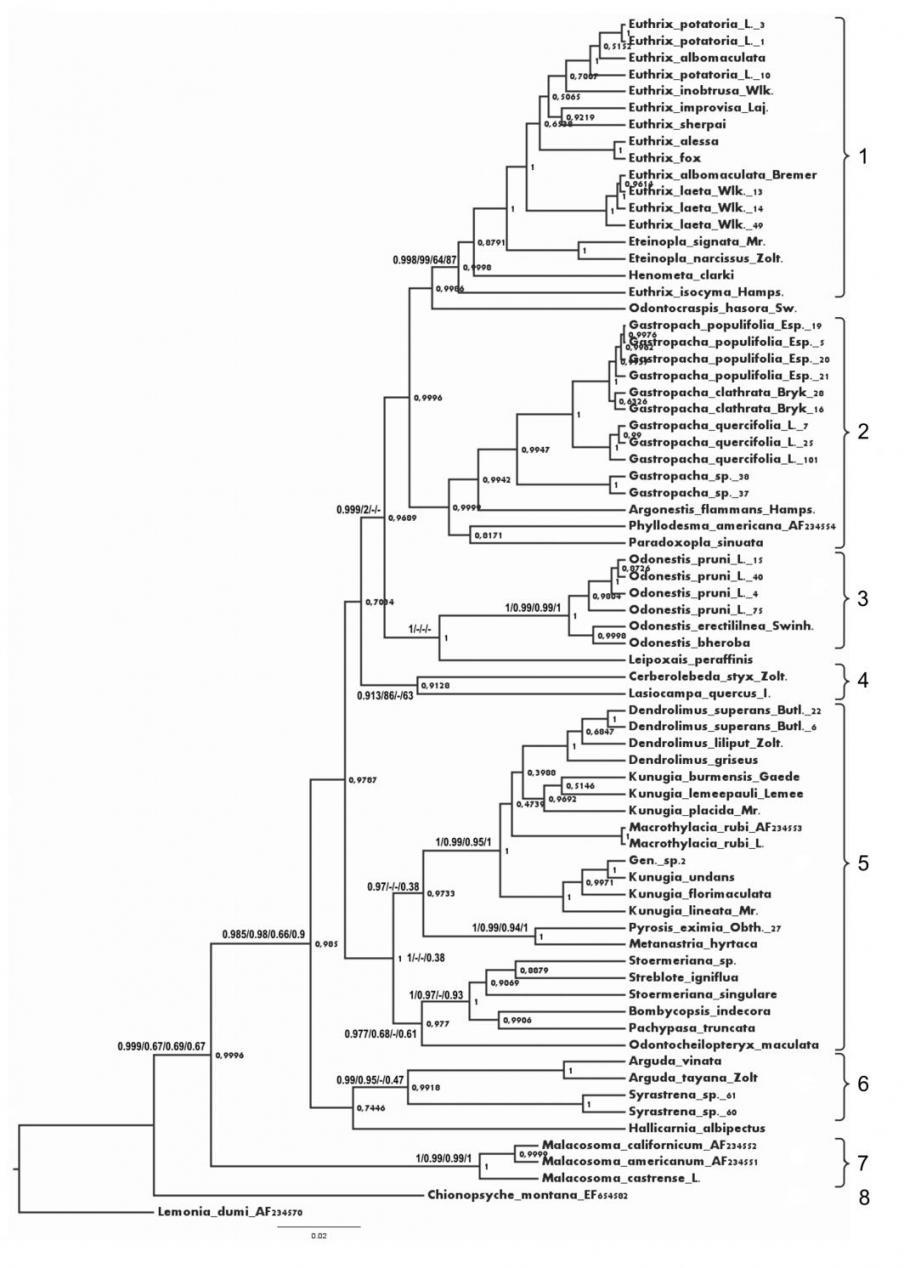
Рис. 1. Схема филогенетических отношений родов коконопрядов на основании изучения полиморфизма нуклеотидных последовательностей гена EF-1 с использованием метода Байеса.
Значения поддержки (постериорная вероятность для метода BI и бутстреп-поддержка для методoв ML, ME и MP) приведены у основания ветвей в последовательности BI/ME/MP/ML; если в анализе клада не выявлялась, то вместо значения поддержки указан прочерк. Цифрами обозначены: 1 – Selenepherini; 2 – Gastropachini; 3 – Odonestini; 4 – Lasiocampini; 5 – Pinarini; 6 – Argudini; 7 – Malacosomini: Malacosominae; 8 – Chionopsychini: Chionopsychinae; Lemonia dumi – внешняя группа.
не являются столь близкими. Роды Odontocraspis и Leipoxais в каждом из 4 вариантов анализа располагаются в различных кладах и не включаются ни в одну трибу. Однако морфологически оба рода близки к представителям трибы Argudini и, вероятно, должны быть отнесены именно к ней.
Обособленную позицию занял род Hallicarnia, ранее провизорно сближавшийся с представителями ветви Argudini; такая точка зрения не поддержана молекулярно-генетическим анализом, и положение этого рода в системе семейства требует уточнения. Неопределенным остается и положение рода Macrothylacia. Признаки строения гениталий и особенности морфологии гусениц сближают этот род с Lasiocampini, но его включение в состав последней трибы пока несколько провизорно.
Представители рода Stoermeriana оказались разнесенными на схеме в удаленные ветви одной клады, что неудивительно, поскольку этот пара- или полифилетический род распадается на несколько морфологически четко очерченных видовых групп.
Несмотря на то, что в BI род Argonestis включается в Gastropachini, морфологически он явно близок к Odonestis. Такое родство с высокой бутстреп-поддержкой отражено также в ML, что позволяет рассматривать его в составе Odonestini.
4.3. Обсуждение.
Полученная кладограмма, в целом, поддерживает идею о синонимизации Pinarinae с Lasiocampinae, рассматриваемых как отдельные трибы Pinarini и Lasiocampini. Вместе с тем, морфологически представители обеих этих групп хорошо отличаются друг от друга, обладая собственными аутапоморфиями. С другой стороны, большинство родов, традиционно рассматривающихся различными авторами в составе Lasiocampini (Anadiasia, Arguda, Euthrix, Pachypasa и др.), в действительности являются типичными членами пинароидной линии. Это подтверждается как признаками строения генитальных структур, так и особенностями развития преимагинальных стадий.
Нет оснований поддержать ранг подсемейства для неотропических Macromphaliinae, которые явно близкородственны Poecilocampinae. Для последних отмечено 28 (у Trichiura crataegi) или 36 (у Poecilocampa populi) хромосом в гаплоидном наборе, против типичных 30-31 хромосом у прочих родов семейства. Недавние исследования показали, что Macromphaliinae также обладают гаплоидным набором, равным 28. Наличие такой важной синапоморфии служит основанием для понижения статуса подсемейства Macromphaliinae до трибы Macromphaliini, stat. nov.
До настоящего времени молекулярно-генетические исследования не затронули виды подсемейства Chondrosteginae, но крайняя специализация его представителей позволяет сохранить за группой статус подсемейства.
На основании сопоставления результатов морфологического и молекулярно-генетического анализа нами предлагается следующий вариант системы коконопрядов мировой фауны:
Lasiocampidae Harris, 1841
1. Chondrosteginae Tutt, 1902
= Chondrostegidae Tutt, 1902
1.1. Chondrostegini Tutt, 1902
2. Poecilocampinae Tutt, 1902
= Trichiurinae Tutt, 1902
= Macromphaliinae Franclemont, 1973
2.1 Poecilocampini Tutt, 1902
= Trichiuridi Tutt, 1902
2.2. Macromphaliini Franclemont, 1973
3. Chionopsychinae Aurivillius, 1927
3.1. Chionopsychini Aurivillius, 1927
4. Malacosominae Tutt, 1902
= Malacosomatinae Aurivillius, 1927
4.1. Malacosomini Tutt, 1902.
5. Lasiocampinae Harris, 1841
= Lachneidae sensu Tutt, 1902
= Eutrichidae sensu Tutt, 1902
= Pachygastriinae Tutt, 1902
= Metanastriinae Tutt, 1902 (partim)
= Pinarinae Kirby, 1892
= Eutrichidae Tutt, 1902 (partim)
= Diplurinae Tutt, 1902
= Dendroliminae Tutt, 1902
= Pachypasinae Tutt, 1902
= Trabaliinae Tutt, 1902
= Chileninae Tutt, 1902
= Cosmotrichinae Tutt, 1902
= Eutrichinae Tutt, 1902
= Nadiasinae Tutt, 1902
= Gastropachinae Newm. & Dyar, 1893
= Gonometinae Aurivillius, 1927
5.1. Lachneini Grote, 1888
5.2. Lasiocampini Harris, 1841
= Pachygastriidi Tutt, 1902
= Lambessidi Tutt, 1902
= Metanastriidi Tutt, 1902 5.3. Macrothylaciini Tutt, 1902
5.4. Selenepherini Tutt, 1902
= Dipluridi Tutt, 1902
= Chilenidi Tutt, 1902
= Cosmotrichidi Tutt, 1902
= Routledgiidi Tutt, 1902
= Nadiasidi Tutt, 1902
5.5. Trabaliini Tutt, 1902.
5.6. Pinarini Kirby, 1892.
= Paralebedidi Tutt, 1902
= Dendrolimidi Tutt, 1902
= Pachypasidi Tutt, 1902
= Bhimidi Tutt, 1902
5.7. Gastropachini Newm. & Dyar, 1893
= Eutrichidi Tutt, 1902
5.8 Odonestini Tutt, 1902
5.9. Argudini trib. nov.
Диагнозы всех надродовых таксонов с указанием типовых родов даны в тексте диссертационной работы.
Глава 5. Особенности распределения и фауногенеза коконопрядов
по материкам
5.1. Видовое разнообразие коконопрядов различных зоогеографических областей рассмотрено на основе схемы, предложенной О.Л. Крыжановским (2002). По фауне коконопрядов, как и медведиц (Дубатолов, 2007), Чилийско-Патагонская область не обнаруживает отличий от Неотропической области ни на видовом, ни на родовом уровне и потому рассматривается в составе Неотропического, а не Нотогейского царства, а Папуасская область, близкая к Австралийской, переносится из Палеотропического царства в Нотогейское.
5.1.1. Голарктическое царство. Известно 56 палеарктических и 17 неарктических родов, а также 389 и 66 видов, соответственно. Общими для Неарктики и Палеарктики являются роды Phyllodesma и Malacosoma, а общие виды для них не известны. Уровень видового эндемизма высок, и около 90% видов не выходят за пределы царства. Для Голарктического царства известен лишь один эндемичный таксон надродового ранга – триба Malacosomini; вероятно, триба Lasiocampini также строго эндемична для хорона.
Бореальная область сильно обеднена родами (28) и видами (75), которые связаны в основном с хвойными и мелколиственными лесами. Доля аркто-альпийских видов ничтожна; единственный вид с аркто-альпийским ареалом (Eriogaster arbusculae) эндемичен для Палеарктики.
Область Древнего Средиземья. Известно только 26 родов с высоким уровнем эндемизма (9); среди эндемиков Psilogaster, Pachypasa, Gufria. Также высок уровень видового эндемизма: 92 вида из 112 известных (82%) – эндемики; многие с точечными ареалами (Malacosoma laurae, M. mixtum, M. squalorum, Macrothylacia digramma, Eriogaster philippsi, Phyllodesma kermesifolia, Ph. suberifolia, Ph. priapus и др.).
Восточноазиатская область. В целом из области известны 41 род и 210 видов с уровнем эндемизма соответственно 24,4% и 52,4%. Среди эндемичных родов Lenodora, Cosmeptera, Eteinopla, Pachypasoides, Similodora, Somadasys, Syrastrenopsis, Notogroma, Amurilla. Условно субэндемичны хозяйственно важные Cosmotriche и Dendrolimus. Своеобразие хорона объясняется его разнообразными ландшафтами и складчатостью в виде горных систем, среди которых особо значимыми являются Гималаи.
Сонорская область. Фауна области насчитывает 17 родов и 66 видов, причем 2 рода общие с фауной Палеарктики (Malacosoma и Phyllodesmа). Эндемичных родов всего 4 (Apotolype, Hypopacha, Heteropacha и Edwardsimemna), но большинство видов эндемичны для хорона, и в Неотропику проникает лишь 6-8 видов.
5.1.2. Палеотропическое царство. Наиболее богатое родами и видами царство, различные области которого имеют специфическую фауну коконопрядов. При этом часть видов в ходе эволюции проникали на территории смежных областей, сформировав современный сложный таксономический состав каждого хорона. Общих видов, распространенных по всей территории царства, нет, общих рода 2 (Streblote и Estigena). Эндемичных надродовых таксонов нет.
Ориентальная (Индо-Малайская) область. Высок уровень видового эндемизма из-за большого количества океанических островов. Известно 32 рода (2 эндемичных – Voracia и Eleganteda) и 208 видов, среди которых более 100 эндемичных. В родах Trabala, Paralebeda, Lebeda, Metanastria, Estigena, Arguda наблюдается целый веер аллопатрических островных видов.
Афротропическая область. Отмечено 77 родов и около 600 видов. Все виды эндемичны. Из надродовых таксонов, помимо эндемичных Chionopsychinae, для области известны автохтонные трибы Chondrostegini, Pinarini и Selenepherini и проникшие из Индо-Малайской области Trabaliini и Gastropachini. Ареал большинства родов жестко ограничен на севере пустыней Сахара, и лишь единичные специализированные виды (из родов Braura, Beralade, Sena, Streblote, Anadiasa) проникают в ее оазисы.
Мадагаскарская область. Для острова отмечено 27 родов и 95 видов коконопрядов; при этом 25 родов эндемичны, и лишь Lechriolepsis и Odontocheilopteryx представлены также в тропической Африке. Впрочем, родство африканских и мадагаскарских представителей рода не бесспорно и отражено лишь в сильном внешнем сходстве. Все виды эндемичны.
5.1.3. Неотропическое царство. Фауна коконопрядов насчитывает 496 видов из 16 родов, причем ее ядро составляют виды 6 близких родов (30 видов Artace, 351 вид Euglyphus, 16 видов Macromphalia, 31 вид Nesara, 29 видов Titya и 98 Tolype), относящихся к трибе Macromphaliini.
Неотропическая область. Отмечено около 450 видов из 16 родов; 1 род (Malacosoma) проникает из Сонорской области. Большинство видов эндемичны, но эндемичных родов нет; родовая система для хорона разработана слабо и нуждается в пересмотре. Все коконопряды Неотропики являются дендробионтами; равнинные виды широко распространены в тропических лесах Амазонии, но горные виды Nesara, Titya, Laruma, Norapidia, Pompeja, Tolitia и Prorifrons узко локальны.
Чилийско-Патагонская область. Фауна коконопрядов слабо изучена, известна только для Чили (Ureta, 1957), откуда указаны 2 рода Macromphalia и Euglyphus с 12 видами. Эндемичных родов нет; степень эндемичности видов не выяснена – считается, что они субэндемичны для хорона.
5.1.4. Нотогейское царство. Несмотря на небольшое число отмеченных здесь родов (17) и видов (94), фауна коконопрядов Нотогеи крайне специфична и полностью эндемична и на родовом и на видовом уровнях.
Австралийская область. Фауна коконопрядов Австралии насчитывает 71 вид из 12 родов из триб Pinarini и Argudini; все они эндемичны, не имеют прямых связей с родами Ориентальной подобласти, и лишь восточноавстралийские Digglesia и Cyclophragma обладают сродством к папуасским Pararguda и Pseudophyllodes. Эндемичных таксонов надродового уровня нет.
Новозеландская область. Коконопрядов нет (Dugdale, 1988).
Папуасская область. Крайне специализированная фауна коконопрядов хорона насчитывает 23 вида из 5 родов (Isostigena, Pseudophyllodes, Pararguda, Sporostigena и новый род для “Taragama” proserpina), относящихся к трибам Pinarini и Argudini; часть видов до сих пор еще не описаны. Все виды и роды являются эндемиками Новой Гвинеи, не обнаруживают родства с видами Индо-Малайской области, но близки к таковым Австралии, что позволяет рассматривать Папуасскую область внутри Нотогейского царства. Несмотря на высокий уровень родового и видового эндемизма, эндемичных таксонов надродового уровня нет.
5.2. Анализ неоднородности фауны коконопрядов. Для выявления неоднородности фауны коконопрядов Земного шара, территории всех континентов и острова были рассмотрены в рамках 149 относительно небольших участков, для которых оказалось возможным выявить достоверный набор обитающих там видов (Приложение 3). При типологическом анализе локальных фаун на уровне родов использован коэффициент сходства Жаккара-Наумова для количественных признаков. Анализ проведен с помощью метода факторной классификации В.А. Трофимова (1976). В результате было получено 23 типа фаун (рис. 2).
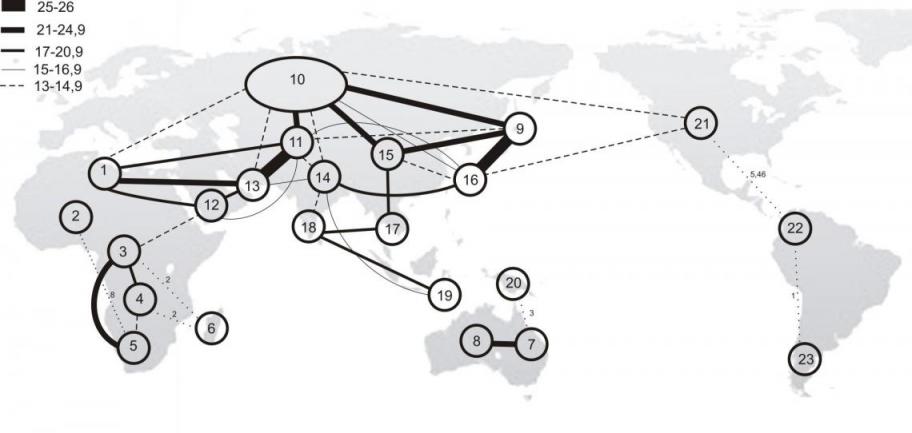
Рис. 2. Граф неоднородности фауны коконопрядов на уровне родов. Сходство фаун между регионами указано линиями пропорциональной толщины (в %%).
На родовом уровне довольно четко выражены 4 группы фаун: Евразиатская, Афротропическая, Новогвинейская и Американская, которые соответствуют крупным зоогеографическим хоронам. При этом фауны Северной Африки и аридных зон Сахеля обладают высоким уровнем сходства с Палеарктической фауной; отдельные фауны Евразии, имея высокий уровень сходства (19-21% и выше) по своему родовому составу, дают четкое разделение на Палеарктическую и Ориентальную области. Фауна Приморья (9), в отличие от Среднеамурской, оказалась в одном кластере с фауной Ориентальной области, и уровень связи по градиенту Китай–Приморье–Индокитай оказался высоким, более 19%. Среднеамурско-Восточноазиатско-Тибетская смешанная фауна (15) имеет высокий уровень сходства как с палеарктической Европейско-Кавказско-Переднеазиатско-Сибирской фауной (10), так и с Южнокитайско-Индийско-Зондской фауной (17) – соответственно 21% и 17%, и является переходной между фаунами Палеарктики и Ориентальной области. Гималайско-Гиндукушская горная фауна (14) обособлена от собственно Ориентальной, имея с ней сходство всего 11%. Крайне обособленными оказались и фауны Малых Зондских островов, что связано с обедненностью их видового состава.
Фауна Афротропической области образовала три тесно связанных между собой кластера (3, 4, 5). Сходство фаун пустынь Намибии и Сахели невелико из-за бедного видового состава. Уровень сходства континентальной фауны с фауной Мадагаскара (6) очень низкий – 2%, всего 2 общих рода. Через Аравию с ее сильно обедненным вариантом аридных фаун Эфиопии и Сомали прослеживается связь фауны Афротропики с фаунами Загроса (13) и Северной Африки (1), которая может отражать историческое расселение афротропических элементов с образованием изолятов для видов, вытесняемых на край ареалов в ходе плейстоценовой аридизации климата.
Отсутствие коконопрядов на Молуккских островах делает возможным провести границу между Ориентальной и Нотогейской фаунами по линии Лайдеккера. Фауна Новой Гвинеи имеет очень слабые связи (3%) с фауной Австралии, но не обнаруживает связей с Патагонией (23). Последняя, хоть и объединяется с фаунами Неарктики и Неотропики (21, 22), но при крайне низком уровне сходства (1%); вероятно, это является следствием ее слабой изученности.
Сходство между фаунами коконопрядов Палеарктики и Неарктики незначительно, причем последняя оказывается значительно ближе к палеарктической (13%), чем к центральноамериканской (5,5%). Предполагается, что в результате комплексной ревизии Неарктической фауны уровень сходства между ними существенно возрастет.
5.3. Эндемизм и его особенности у коконопрядов. Распределение коконопрядов жестко связано с границами материков, и виды-космополиты среди них не известны (табл. 1).
Низкая летная активность, а у некоторых видов (Chondrostega, Mesocelis, отдельные Lasiocampa и Tolype) и бескрылость самок накладывают ограничения на распространение видов, и коконопряды часто образуют узколокальные таксоны. Практически все высокие горные системы населены эндемичными
Табл. 1
Особенности видового и родового эндемизма
коконопрядов различных хоронов
| Хорон | Родов | Видов | ||||
| Всего | Эндемичных | %% | Всего | Эндемичных | %% | |
| Голарктическое царство | 73 | 17 | 23,3 | 455 | 390 | 85,7 |
| Бореальная область | 28 | 2 | 7,1 | 75 | 8 | 10,7 |
| Область Древнего Средиземья | 26 | 9 | 34,6 | 112 | 92 | 82 |
| Восточноазиатская область | 41 | 10 | 24,4 | 210 | 110 | 52,4 |
| Сонорская область | 17 | 4 | 23,5 | 66 | 60 | 90,9 |
| Палеотропическое царство | ||||||
| Ориентальная область | 32 | 2 | 6,3 | 208 | 102 | 48 |
| Афротропическая область | 86 | 80 | 93 | 600 | 600 | 100 |
| Мадагаскарская область | 26 | 24 | 92,3 | 94 | 94 | 100 |
| Неотропическое царство | 16 | 3 | 18,8 | 496 | 470 | 94,8 |
| Неотропическая область | 16 | 0 | 0 | 450 | 398 | 88,9 |
| Чилийско-Патагонская область | 2 | 0 | 0 | 13 | 11 | 84,6 |
| Нотогейское царство | 17 | 17 | 100 | 94 | 94 | 100 |
| Австралийская область | 12 | 12 | 100 | 71 | 71 | 100 |
| Новозеландская область | 0 | 0 | 0 | 0 | 0 | 0 |
| Папуасская область | 5 | 5 | 100 | 23 | 23 | 100 |
видами; к ним же привязаны и центры эндемизма семейства. В Евразии наибольшее число локальных эндемиков приходится на Гималайско-Тибетский регион (60 видов и 3 рода – Baodera, Similodora, Phachebia). Их число уменьшается по направлению к северу, и в горах Центрального и Восточного Китая эндемичными являются хвоегрызущие представители родов Cosmotriche, Dendrolimus, Kunugia, Paradoxopla, Takanea и Pachypasoides (20 видов). Два вида эндемичны для островов Японии, 1 – для Алтая. На Урале и в Альпах эндемичные виды не отмечены.
Количество эндемиков уменьшается от Гималайско-Тибетского региона также к западу и югу; в направлении Передней Азии эндемизм связан с преимущественно с пустынями, а в странах Юго-Восточной Азии отсутствуют мощные горные системы. В Копет-Даге обитают 2 эндемика видового и 1 родового ранга, в Передней Азии отмечены 16 эндемичных видов из родов Sena, Chilena, Eriogaster, Chondrostega и Lasiocampa). Из Западного Средиземноморья известно 12 видовых и 7 родовых/подродовых эндемиков, которые заселяют аридные биотопы Северной Африки (Psilogaster, Chondrostega) и горы Атлас (Malacosoma, Gufria, Phyllodesma, Trichiura).
В Неотропической области центром эндемизма являются Анды (3 рода и около 60 видов), в Сонорской области – Скалистые горы (3 рода и 7 видов). В Афротропической области несомненный центр эндемизма – пустыня Сахара, где эндемичны 1 род (Ergolea) и 22 из 28 известных видов. Расположенная южнее территория практически лишена горных систем и узколокальных эндемиков. Исключением могут считаться кратеры Нгоро-Нгоро и Килиманджаро, где насчитывается 5 условных эндемиков.
Видовых ареалов коконопрядов Африки определяются в большей степени границами климатических зон, чем особенностями рельефа. Крайне специфичен видовой состав Южноафриканской подобласти, откуда известно 130 видов, 85 из которых эндемичны. Большинство из них ксерофильны, обитают в саваннах и не заходят в дождевые тропические леса Гвинейско-Конголезской подобласти, где сосредоточено подавляющее большинство коконопрядов Афротропики (более 420 видов).
5.4. Особенности фауногенеза коконопрядов на различных материках.
5.4.1. Коконопряды Евразии. В Евразии известно 58 родов коконопрядов с 499 видами из подсемейств Chondrosteginae, Poecilocampinae, Malacosominae и Lasiocampinae. Наличие современных центров видового разнообразия и центров эндемизма большинства триб (Poecilocampini, Malacosomini, Macrothylaciini, Lasiocampini и Lachneini) в Средиземноморье позволяет предположить, что их предковые формы имеют Европейское (Средиземноморское) происхождение, в то время как представители трибы Pinarini проникли на территорию Евразии с Индийской материковой плиты. Дальнейший фауногенез этих групп связан с их экспансией, а активному видообразованию способствовала складчатость Гималаев, Тибета, гор Центрального и Восточного Китая и Малой Азии, в меньшей степени – Гиндукуша, Памира, Тянь-Шаня, Копет-Дага. Позже началось интенсивное видообразование на океанических островах.
Представители родов Dendrolimus, Pachypasoides, Cosmotriche, Takanea и Kunugia трофически связаны с хвойными; они являются эндемиками Палеарктической и Ориентальной областей и имеют Южнокитайский и Синотибетско-Восточно-гималайский центры видового обилия. Более 35 центрально- и юго-восточноазиатских видов рода Dendrolimus большей частью эндемичны для отдельных горных систем южного Китая, а максимум видового разнообразия рода Pachypasoides (16 видов) приходится на Тибет, горы Юньнани и Северного Вьетнама. Таким образом, можно считать, что формирование пра-Dendrolimus и пра-Pachypasoides приходится на (поздне)меловой период и рассматривать южный Китай как центр происхождения хвоегрызущих коконопрядов.
Современный облик флора хвойных приобрела в позднем мелу, в период интенсивных горообразовательных процессов (Криштофович, 1941; Тахтаджян, 1987), а их проникновение на север было связано с волнами похолодания в позднем миоцене-плиоцене, в ходе которых они заняли экологические ниши выпадающих теплолюбивых покрытосеменных. Расселение хвоегрызущих коконопрядов на север произошло вслед за экспансией хвойных при общем похолодании климата. При этом «северная» ветвь рода Dendrolimus дала 2 вида: европейский D. pini, трофически связанный с сосной обыкновенной, и восточно-сибирский D. superans, развивающийся на лиственнице. Оба этих вида пространственно и биологически были разобщены еще с миоцена, и лишь совсем недавно (вероятно, в последние 100-150 лет), за счет медленной миграции навстречу друг другу, образовалась зона их симпатрии на участке северо-востока европейской части и смешанных лесов по линии Челябинск – Новосибирск – Иркутск.
5.4.2. Коконопряды Северной Америки. Формирование фауны коконопрядов Северной Америки тесно связано с существованием Берингийского моста. Считается общепризнанным наличие нескольких волн миграций по нему из Азии в Северную Америку. В первую волну миграции палеоцена–эоцена с территории Азии вместе с широколиственными представителями тургайской флоры в Америку проникли представители рода Lebeda или его предковые формы. При общем похолодании климата они вместе со своими кормовыми растениями отступили на современную территорию США, где сформировались несколько родов: Prorifrons, Dicogaster, Edwardsimemna, Eutachyptera, Caloecia. Связь с хвоегрызущими коконопрядами группы Dendrolimus–Pachypasoides предполагается нами для фенотипически сходного сонорского рода Gloveria. Чуть позже в Америку могли проникнуть предки родов Hypopacha и Heteropacha.
Предковые формы трибы Macromphaliini были, вероятно, представителями тропической и субтропической азиатской фауны и проникли с первой или второй волной миграции по Берингийскому мосту из Палеарктики, отступив при похолодании на юг Североамериканского континента. С формированием Панамского перешейка они проникли в Южную Америку, где радиировали в подходящих биотопах, образовав огромное количество (около 500) близких видов, отличающихся прежде всего деталями рисунка крыла – признаком неустоявшихся молодых таксонов.
Смена в позднем эоцене и плейстоцене тропических лесов Берингии мелколиственными умеренной зоны закрыла путь миграции тропических форм коконопрядов, но открыла коридор для таких представителей современной палеарктической фауны, как Malacosoma и Phyllodesma. Поскольку их инвазия произошла относительно недавно (в конце эоцена–плейстоцене, то есть около 20 тысяч лет назад), они хоть и дивергировали экологически, но в большинстве случаев не достигли еще уровня самостоятельных видов.
Таким образом, формирование фауны коконопрядов Северной Америки связано с 3 основными волнами миграций. Первой волной было проникновение азиатских тропических форм в позднем палеоцене – начале эоцена; вторая волна миграций позднего эоцена привела к проникновению в Неарктику видов лиственных лесов умеренного пояса и образованию монотипических родов Hypopacha и Heteropacha. Последняя волна прошла при распространении холодоустойчивых мелколиственных лесов и привела к инвазии представителей родов Malacosoma и Phyllodesma, находящихся сегодня на стадии интенсивного видообразования.
5.4.3. Коконопряды Южной Америки. Фауна коконопрядов Южной Америки пока слабо изучена, включает на сегодняшний день всего 18 родов и дана в предварительных оценках. Центр видового обилия – Андийский, откуда известно около 50% всех описанных видов. Практически полная изоляция Южной Америки на протяжении долгого исторического времени наложила свой отпечаток на формирование ее фауны. Не вызывает сомнения, что широко распространенные на ее территории представители нескольких близких родов (Macromphalia, Tolype, Euglyphus) происходят от общего предка и часто образуют комплексы ‘полувидов’ или молодых видов. Их явное сходство с представителями трибы Poecilocampini позволяет предположить, что они произошли от каких-то анцестральных форм, близких к Trichura и Poecilocampa, которые проникли в Северную Америку из Евразии через Берингов мост в позднем палеоцене – начале эоцена, а затем широко расселились к югу.
5.4.4. Коконопряды Африки. Ревизия коконопрядов Африки только начата, и затронула пока лишь 6 родов. Всего на территории континентальной Африки отмечено 86 родов (число которых, несомненно, возрастет) с 658 эндемичными видами. Наиболее богата фауна Афротропической области – 609 видов (92,4%); совокупная доля эндемиков Сахары и видов Средиземноморского комплекса составляет всего 7,6%. При этом из 9 родов, отмеченных в Сахаре, лишь 2 (Lasiocampa и Ergolea) являются вселенцами из Средиземноморья, а у представителей 7 родов обнаружены связи с видами, известными с более южных территорий.
Во второй половине эоцена – начале олигоцена влажные тропические леса, занимавшие до этого большую часть Африки и мозаично распространенные вдоль побережья Тетиса до субтропиков Голарктики (Мейен, 1987), к северу от экватора стали замещаться разреженными саваннами, открыв мост для проникновения на север африканского континента аридных кафрских коконопрядов. В конце олигоцена этот путь прервался формирующимися экваториальными лесами, определив дальнейшую эволюцию вселенцев в условии изоляции. В результате в аридных регионах севера континента сформировался своеобразный род Chondrostega, проникший позже и на Европейский субконтинент, в Переднюю Азию и Аравию.
Ближе к побережью Тетиса в олигоцене леса саванного типа заменялись лавровыми, образующими своего рода экотонную зону между Европой и Африкой (Мейен, 1987), которая препятствовала обмену фаун. К концу плиоцена с усилением аридизации началось активное внедрение в Северную Африку новых ксерофитов и галофитов (Artemisia, Ephedra, Tamarix, солянок и др.) из Малой Азии, что обеспечило возможность расселения растительноядных представителей родов Sena, Beralade, Chondrostega, Bufoidia из Африки в Азию, a им навстречу – видов из родов Chilena и Pachygastria.
Некоторые коконопряды проникли из Афротропики в Азию в обход Аравии или по влажным торопическим лесам ее южной оконечности, сохранявшимся здесь в миоцене и плейстоцене (Нamilton, 1974). Так, центром происхождения рода Streblote предполагается прибрежная зона Восточной Африки, а время становления приходится на начало эоцена, после которого продвижение на север столкнулось бы с преградами в виде обширных пустынь. Вероятно, предковая его форма, близкая к S. diplozona, проникнув на север и запад Африканского континента, дала группу из нескольких самостоятельных видов, а дальнейшая экспансия на восток привела к активному видообразованию в горах Юго-Восточной Азии и на островах Тихого океана.
Обратный процесс демонстрируют исходно азиатские представители родов Trabala и Estigena, проникшие из Азии в Африку по тропическим лесам северного побережья Индийского океана и Аравийского полуострова. Данный мост прервался уже в раннем палеоцене, и в результате в лесах Афротропики в условиях изоляции сформировались 6 близких видов неописанного рода, близкого к Trabala, а также эндемичный вид рода Estigena.
5.4.5. Коконопряды Австралии. Фауна коконопрядов Австралии включает 71 описанный вид из 12 родов (Turner, 1924; Common, 1990; Edwards, 1996). Все виды и роды эндемичны и не имеют прямых связей с представителями фауны Индо-Малайской области. В течение позднего палеогена и почти всего неогена биота континента развивалась в практически полной изоляции, что придало ей неповторимые черты. Именно в это время произошло, вероятно, окончательное формирование таксономической структуры австралийской фауны коконопрядов. В верхнем плиоцене и нижнем плейстоцене континент соединялся с Новой Гвинеей в районе Торресова пролива и Сахульской отмели, когда и могла произойти экспансия представителей рода Cyclophragma, 3 вида которого известны из восточной части Новой Гвинеи и 2 – из Австралии.
5.5. Эремиальные виды коконопрядов. К пустыням приурочены крайне специализированные роды семейства, такие как Chondrostega, Chondrostegoides, Chilena, Sena, Beralade и Ergolea, а также отдельные представители родов Streblote, Anadiasa, Eriogaster и Lasiocampa, которых можно назвать эремиальными (эремическими) таксонами (Wiltshire, 1982). Они выработали целый комплекс специфических приспособительных признаков: эстивацию, моновольтинность, лет в осенне-весенний период, окукливание в почве в плотном коконе. Доминирующая у них светлая окраска способствует защите от перегрева. Для представителей истинно аридной фауны характерна моновольтинность, а часто и многолетняя диапауза. Экстремальные условия среды определяют жесткий стабилизирующий отбор, нередко идущий в условиях инбридинга, как у Chondrostega с их аптерными самками; у родственных им Mesocelis, самки которых не выходят из коконов, имеет место переход к факультативному партеногенезу.
Для многих видов проникновение вглубь пустынь определено наличием там их кормовых растений. Несколько видов из родов Streblote, Anadiasa и Beralade проникают далеко в пески с акациями, но назвать их истинно эремиальными нельзя. Не покидая фактически своего кормового растения, они находят в его кроне вполне мезофильные условия существования и не выработали особых адаптаций к аридным условиям. Ряд видов коконопрядов (группа Sena–Beralade–Chilena) в условиях пустыни приспособились к питанию ксерофитными бобовыми, проникая с ними еще глубже в пески, где даже акация уже не растет. Поскольку резкая аридизация климата и появление в Северной Африке пустынь датируется плейстоценом (Howarth, 1981; Adams, 1981), становление эремиальных видов должно было идти ранее плейстоценового оледенения, вероятно, уже в четвертичном периоде. Позже и в Сахаре и в Аравии имели место влажные периоды, когда часть региона покрывалась лесами и многочисленными реками, которые и отделили новые эремиальные формы Западной Сахары от анцестральных восточных форм. В наиболее характерных эремиальных группах число видов западной группы значительно меньше, чем восточной.
Переход к обитанию в пустынях связан со сменой кормового субстрата и у австралийских (Digglesia, Symphita, Entometa, Porela, Pinara) и американских (Gloveria arizonensis) коконопрядов. В Австралии такой переход – сравнительно частое явление, связанное с приспособлением к обитанию в аридных условиях их основного кормового растения (эвкалипта, реже акации). Многие австралийские коконопряды могут при этом рассматриваться в качестве скорее мезофильных, чем истинно эремиальных форм, поскольку являются обитателями крон деревьев и, очевидно, приспособились к обитанию в аридных местностях относительно недавно.
Глава 6. Особенности островного эндемизма у коконопрядов
Коконопряды, как группа слабо мигрирующих чешуекрылых, идеально подходит для изучения видообразования на островах (Золотухин, 2009). Нами были проанализированы фауны Мадагаскара, Цейлона, Японского архипелага, Тайваня, Суматры, Борнео, Явы, Филиппинских островов, Сулавеси, Новой Гвинеи и острова Флорес (табл. 2).
Таблица 2
Таксономический состав семейства на островах различных размеров
| Остров | Площадь, в км2 | Родов | Видов | ||
| Всего (в том числе эндемичных) | % эндемичных | Всего (в том числе эндемичных) | % эндемичных | ||
| Новая Гвинея | 829 000 | 6 (6) | 100 | 22 (22) | 100 |
| Борнео | 734 000 | 22 (0) | 0 | 63 (15) | 23.8 |
| Мадагаскар | 590 000 | 26 (24) | 92.3 | 94 (94) | 100 |
| Сулавеси | 453 000 | 10 (0) | 0 | 15 (11) | 73.3 |
| Суматра | 434 000 | 21 (0) | 0 | 69 (9) | 13.0 |
| Ява | 125 000 | 9 (0) | 0 | 12 (7) | 58.3 |
| Лусон | 105 600 | 30 (0) | 0 | 22 (17) | 77.3 |
| Минданао | 94 600 | 33 (0) | 0 | 33 (20) | 60.6 |
| Шри-Ланка | 65 000 | 10 (0) | 0 | 11 (1) | 9.1 |
| Тайвань | 36 000 | 21 (0) | 0 | 24 (12) | 50.0 |
| Флорес | 15 000 | 6 (0) | 0 | 11 (10) | 90.9 |
Островное видообразование происходит в условиях географической изоляции исходного таксона и часто приводит к образованию множества викарных видов. Для коконопрядов это особенно характерно для архипелагов, когда даже недалеко расположенные друг от друга острова оказываются заселенными разными таксонами. Ранг этих таксонов может быть различным – от хороших видов или подвидов до популяций неясного статуса, претендующих на роль «полувидов», находящихся в процессе видообразования. Например, род Trabala, насчитывающий на континенте всего 9 видов, на океанических островах представлен 16 эндемичными видами.
На островах материкового происхождения (Суматра, Борнео) уровень эндемизма коконопрядов ниже, и эндемики приурочены в основном к горным цепям. Острова океанического происхождения при более низком уровне видового обилия имеют более высокий уровень видового эндемизма. Зависимости между площадью острова и количеством эндемичных для него видов коконопрядов не наблюдается, однако эндемичные роды известны лишь для самых крупных островов (Новая Гвинея и Мадагаскар) населены эндемичными родами. Острова, связанные единой геологической историей, а следовательно и общими чертами генезиса фаун, обладают высокими показателями их сходства (Палаван–Борнео, Палаван–Суматра, Ява–Борнео, Ява–Суматра), в то время как фауны Сулавеси и Флорес, напротив, весьма самобытны (табл. 3).
Таблица 3
Коэффициент сходства фаун островов Тихого океана,
рассчитанный по формуле Съеренсена-Чекановского
| Острова | Тайвань | Борнео | Суматра | Ява | Сулавеси | Флорес | Палаван | Лусон | Минданао | Негрос |
| Борнео | 6.2 | |||||||||
| Суматра | 7.8 | 74.2 | ||||||||
| Ява | 13.0 | 29.3 | 32.9 | |||||||
| Сулавеси | 0 | 5.1 | 0 | 0 | ||||||
| Флорес | 0 | 2.7 | 2.5 | 8.7 | 0 | |||||
| Палаван | 3.1 | 45.2 | 32.0 | 20.0 | 0 | 4.9 | ||||
| Лусон | 0 | 16.5 | 15.4 | 9.5 | 5.4 | 6.0 | 34.6 | |||
| Минданао | 3.0 | 20.8 | 19.6 | 11.3 | 2.5 | 4.5 | 31.7 | 61.8 | ||
| Негрос | 3.6 | 21.4 | 19.8 | 14.6 | 5.6 | 6.0 | 35.3 | 55.8 | 55.6 | |
| Лейте | 0 | 12.8 | 11.9 | 11.4 | 6.7 | 7.7 | 26.7 | 64.9 | 58.3 | 50.0 |
Можно было бы ожидать, что схожие климатические и геологические условия близлежащих островов сделают границы распространения приуроченных к ним таксонов нечеткими, однако этого не происходит, и ареалы даже близких видов коконопрядов ограничены отдельными островами. Нет никаких подтверждений и рецентной колонизации островов выходцами с соседних континентов или архипелагов, если не брать во внимание немногочисленные факты завоза. Очевидно существование множества ограничений на распространение, обусловленных чисто биологическими механизмами – межвидовой конкуренцией или наличием жестких коэволюционных связей с представителями аборигенной биоты. Особенно актуально это для близких видов одного рода, занимающих идентичные экологические ниши на соседних островах.
Глава 7. Особенности происхождения надродовых таксонов коконопрядов
и формирования их ареалов
Отсутствие палеонтологического материала не позволяет достоверно определить время, место и особенности формирования различных таксонов семейства коконопрядов. Поэтому попытка восстановить происхождение и пути фауногенеза группы делается здесь на основе сопряженного анализа морфологии различных стадий развития, характера трофических связей и распространения.
7.1. Подсемейство Chionopsychinae. Монотипическое афротропическое подсемейство с 3 видами и дизъюнктивным ареалом, разорванным на 2 небольших участка, располагающихся в Западноафриканской (Камерун) и Восточноафриканской (Судано-Замбезийской) подобластях. Подсемейство обладает комплексом чрезвычайно специфических и специализированных морфологических признаков (гениталии самца резко асимметричны из-за редукции левой вальвы и крюковидной трансформации юксты, а гениталии самки сильно модифицированы за счет сильного разрастания оснований апофиз), так что его родственные связи не очевидны. Однако в целом генерализованное строение вальв и наличие ункуса и гнатоса свидетельствуют об архаичности Chionopsychinae, а крайняя стенобионтность видов и их узкие ареалы указывают на его реликтовый статус. Происхождение подсемейства связано с Африканским континентом, и видообразование шло аллопатрически, при разбиении исходного ареала формирующимися дождевыми лесами.
7.2. Подсемейство Chondrosteginae. Небольшое архаичное подсемейство с широкодизъюнктивным средиземноморско-южноафриканским ареалом, насчитывающее в своем составе 4 рода и около 20 стенобионтных эремиальных видов. Его представители также сочетают целый набор признаков в плезиоморфном состоянии (генерализованный план строения вальв, наличие двураздельного ункуса и гнатоса, практически полный набор генитальных мышц и др.) с чертами крайней специализации. Последние проявляются не только в развитии комплекса признаков, облегчающих выход из подземного кокона (лобные шипы, когтевидные придатки голени), но и в четко выраженной тенденции к брахиптеризации самок, отмеченной в 3 из 4 родов подсемейства. Считается (Стекольников, Золотухин, 1994), что Chondrosteginae филогенетически близки к Lasiocampini.
Происхождение подсемейства мы связываем с Южной Африкой, где отмечается центр его родового обилия и где встречаются виды с крылатыми самками (Chondrostegoides, Trichiurana). Дальнейшая его эволюция, вероятно, шла в пределах аридных зон восточного побережья Африки, откуда оно проникло в Средиземноморье, дав там вторичный центр видообразования, в котором распространен комплекс молодых видов группы Chondrostega tingitana (в Северной Африке) и Ch. pastrana (в Передней Азии). Архаичность группы, ее крайняя стенобионтность и узкие ареалы видов подчеркивают реликтовый статус таксона.
7.3. Подсемейство Poecilocampinae. Подсемейство с 2 трибами и широкодизъюнктивным голарктическо-южноафриканско-неотропическим ареалом. Центр происхождения связан с Древним Средиземьем.
7.3.1. Триба Poecilocampini. Триба с 6 родами и широкодизъюнктивным голарктическо-южноафриканским типом ареала, представляющая собой достаточно архаичную группу в силу генерализованного строения генитального аппарата самцов, простого крылового рисунка, а также максимально полного набора мышц в гениталиях самцов. Однако кариотип ее представителей сильно модифицирован из-за разделения или слияния отдельных хромосом (28 и 36 хромосом в гаплоидном наборе вместо типичных для семейства 30-31). На основании топографии мышц генитального аппарата считается, что Poecilocampini родственны Gastropachini и Pinarini. Центром видового обилия и происхождения трибы является Средиземноморье; вторичный центр видообразования – Восточнокитайский, где обитают 4 вида рода Poecilocampa. Все виды дендробионтные, преимущественно горные, некоторые с очень узкими ареалами. Из аридных частей Южной Африки известны 3 вида рода Bombycomorpha, сохранившие и здесь связь с древесной растительностью (Rhus spp.). Представители трибы, распространенные в зоне мелколиственных лесов Восточной Палеарктики, в палеогене-неогене по Берингийскому мосту проникли в Северную Америку, где дали начало особому роду Hypopacha.
7.3.2. Триба Macromphaliini. Триба включает 6 родов и 450 дендробионтных видов, распространенных в Новом Свете. Предполагается, что она берет начало от предковых форм тропической и субтропической фаун Восточной Азии, близких к Тrichiura или Poecilocampa, которые проникли на территорию Северной Америки через Берингию в палеогене, а при последующем похолодании отступили далеко на юг и радиировали в Южной Америке с образованием огромного количества близких, зачастую находящихся в процессе становления видов. Центром происхождения и центром видового обилия трибы следует признать Неотропическую область, откуда шло распространение ее представителей как на юг, до Чилийско-Патагонской области, так и на север, до Бореальной области.
7.4. Подсемейство Malacosominae. Небольшое монотипическое подсемейство с голарктическим типом ареала и единственным родом, включающим 2 подрода. Подрод Malacosoma s. str. объединяет хортобионтные формы и имеет, вероятно, средиземноморский центр происхождения и видового обилия; вторичным центром видообразования является горный центральноазиатский, откуда известно 3 близких вида. Подрод Clisiocampa включает преимущественно дендробионтные виды и имеет голарктическое распространение. Центр его происхождения центральноазиатский, а вторичные центры видообразования приурочены к горам Восточного Китая и Сонорской области. В Неарктике насчитывается 5 близких молодых видов, сформировавшихся в неогене после проникновении предковой формы из восточной Палеарктики через Берингию.
7.5. Подсемейство Lasiocampinae. Самое крупное, практически всесветно представленное подсемейство, монофилия которого пока не доказана.
7.5.1. Триба Lachneini. Небольшая триба с 2 родами, имеющими средиземноморские ареалы. Cтановление и дифференциация видов происходила в горных системах, прежде всего Малой Азии (Eriogaster s. str.), или в условиях крайней ксерофитизации в аридных биотопах Малой и Центральной Азии (подрод Autosphyla рода Eriogaster) и Африки (Ergolea).
7.5.2. Триба Lasiocampini. Триба включает 5 родов и имеет транспалеарктический ареал. Центром родового разнообразия являются горы Восточного и Южного Китая, а центр видового обилия расположен в аридных биотопах южного Средиземноморья. В некоторых родах видообразование шло в горных районах Центральной Азии (Amurilla, Cerberolebeda, Baodera) или в условиях крайней ксерофитизации аридных биотопов Малой Азии (Pachygastria).
7.5.3. Триба Macrothylacini. Монотипическая триба с 2 средиземноморскими видами, один из которых проникает в Восточную Палеарктику.
7.5.4. Триба Trabaliini. Небольшая триба с 4 палеотропическими родами, из которых 2 распространены в Афротропике, имея центр видового обилия в Западноафриканской подобласти, а 2 – в Ориентальной области, где богато представлены на островах Тихого океана. Происхождение трибы связано с Южной Азией, откуда одна из ветвей линии Trabala в позднем палеогене – неогене проникла в Афротропическую область, сформировав там несколько близких видов (см. главу 5).
7.5.5. Триба Selenepherini. Триба насчитывает 7 евразиатских и 7-10 африканских родов. Большинство родов сконцентрированы в Мадагаскарском, Гималайско-Тибетском и Центрально-азиатском центрах, а центрами видового разнообразия являются Южнокитайский и Мадагаскарский. Представители рода Cosmotriche тесно связаны с голосеменными растениями, и интенсивное видообразование здесь шло в Южном и Восточном Китае. Большинство видов трибы – дендрофаги, но Lenodora и многие Euthrix перешли к хортофагии. Переход к питанию на однодольных (злаки и осоки) привел к активному видообразованию в роде Euthrix, насчитывающем в настоящее время около 40 видов. Приспособление представителей рода Sena к обитанию в аридных биотопах привел к смене кормового субстрата с исходных бобовых на более доступные в условиях пустынь злаки и гречишные и также вызвал интенсивное видообразование (известно 36 видов).
7.5.6. Триба Gastropachini. Довольно крупная и широко распространенная триба с 15 родами, не представленная только в Неотропике и Австралийской области. Все ее виды являются дендрофагами. В Палеарктике известно 5 родов, в Неарктике отмечен единственный монотипический сонорский род Heteropacha. Из Афротропики известно 9 родов, распространенных в основном в Западноафриканской подобласти и преимущественно эндемичных родов (Opisthodontia, Theophasida, Marmonna, Morongea, Weberolegra и др.). Активное видообразование происходило в Западном Средиземноморье в условиях возникающей складчатости Атласа (подрод Epicnaptera рода Phyllodesma), в горах Центрального Китая (Stenophylloides, Gastropacha, Paradoxopla, Phyllodesma s. str.), а также на островах Тихого океана (Estigena, Paradoxopla).
7.5.7. Триба Pinarini. Самая крупная триба всесветного распространения, объединяющая около 100 родов. Все относящиеся к ней виды являются дендрофагами, а некоторые (dendrolimus, Pachypasoides, Notogroma, часть Kunugia) специализируются на питании хвойными. Предполагается, что группа сформировалась на Азиатском субконтиненте; наблюдается сходство на уровне родов между Ориентальной областью и каждой из других крупных зоогеографических областей: Афротропической, Австралийской, Сонорской и Неотропической. Это сходство проявляется, прежде всего, в наличии близкородственных викарных родов (рис. 3). Кроме того, представители некоторых родов проникали из одного хорона в другой, образуя на вновь заселенных территориях хорошо обособленные группы видов (внутри Streblote, Stoermeriana, Dicogaster), которые иногда рассматриваются в ранге самостоятельных подродов или даже родов. Эволюция евразиатских представителей трибы связана с освоением ими горных систем Центральной Азии и дальнейшим проникновением на север и запад, а также с интенсивным видообразованием на островах Зондского архипелага. Все роды Сонорской фауны, по-видимому, произошли от проникших в миоцене предковых форм, близких к Lebeda.

Рис. 3. Викарирующие таксоны трибы Pinarini в фауне различных зоогеографических областей.
7.5.8. Триба Odonestini. Небольшая триба с 2 родами. Центр видообразования расположен в Юго-Восточной Азии, причем лишь 5 видов имеют материковое происхождение, а остальные 15 – островное; наибольшее их обилие наблюдается на Филиппинах и Зондском архипелаге. Некоторые виды рода Odonestis могли проникнуть на континент в ходе колебаний уровня моря в плейстоцене. На севере ареала (Китай, Россия) триба представлена единственным видом Odonestis pruni с транспалеарктическим, фрагментированным в горных частях ареалом.
7.5.9. Триба Argudini. Небольшая триба с 7 родами, имеющими дизъюнктивный афротропическо-ориентально-австралийский ареал. Центром ее происхождения является Гималайско-Тибетский, а наибольшее видовое обилие приходится на острова Тихого океана. На севере ареала (Центральный Китай, о. Тайвань и юг Дальнего Востока России) встречаются лишь отдельные виды родов Arguda, Syrastrenopsis, Takanea, Syrastrena, причем видовые ареалы здесь сильно фрагментированы. В Австралии триба представлена двумя родами (Porela и Digglesia), а из Афротропики известен род Leipoxais.
ВЫВОДЫ
1. В результате ревизии коконопрядов мировой фауны на основе анализа широкого спектра признаков предложена оригинальная система надродовых таксонов семейства, включающая 5 подсемейств с 14 трибами, одна из которых (Argudini) описана как новая. Существенно пересмотрен состав многих родов, а 66 родов полностью ревизованы, описано 27 новых родов, 139 видов и 43 подвида, 20 родовых названий синонимизированы.
2. Изучение морфологии имагинальных и преимагинальных стадий, а также проведение молекулярно-генетического анализа позволило выделить в семействе Lasiocampidae три четко обособленные монофилетические группы родов: Malacosoma, Chionopsyche и Poecilocampa-Macromphalia, которые рассматриваются в ранге подсемейств Malacosominae, Chionopsychinae и Poecilocampinae. Подсемейство Macromphaliinae признается лишь трибой в составе Poecilocampinae. Обоснована необходимость отнесения евразиатских родов семейства к 6 трибам Gastropachini, Odonestini, Selenepherini, Lasiocampini, Pinarini и Argudini; эти трибы, однако, не обладают явственными синапоморфиями, и их объединение в составе подсемейства Lasiocampinае представляется провизорным. Архаичные роды Chondrostega, Chondrostegoides, Mesocelis и Trichiurana объединяются в подсемейство Chondrosteginae на основе признаков морфологической и экологической специализации.
3. Фауны коконопрядов, выделенные на родовом уровне, соответствуют основным крупным зоогеографическим хоронам. Четко выражены 4 группы фаун: Евразиатская, Афротропическая, Новогвинейская и Американская. При этом фауны Северной Африки и аридных зон Сахеля обладают высоким уровнем сходства с Палеарктической фауной, а граница между Палеарктической и Ориентальной фаунами оказывается сильно размытой. Уровень сходства между фаунами коконопрядов Палеарктики и Неарктики незначителен.
4. Центры эндемизма семейства привязаны преимущественно к высоким горным системам; в Евразии это Гималайско-Тибетский регион с Синотибетско-Восточногималайским и Южнокитайским центрами видового обилия. На Африканском континенте несомненным центром эндемизма является пустыня Сахара; отсутствие узколокальных эндемиков в Афротропике объясняется отсутствием здесь высокогорий.
5. Фауна коконопрядов Евразии имеет корни на пра-Индийской материковой плите и на территории пра-Европы, где сформировались представители подсемейств Poecilocampinae, Malacosominae и Lasiocampinae. Практически все виды и большинство родов Афротропической фауны эндемичны и произошли от общих с Индо-Австралийскими предковых форм. Все виды и роды коконопрядов Австралии эндемичны и не имеют прямых связей в фауне Ориентальной области. Американские виды коконопрядов также эндемичны, но обнаруживают тесное родство с видами азиатской фауны, а роды могут быть как общими с Палеарктическими, так и близкими к Индо-Малайским. При формировании фауны Северной Америки можно предположить наличие 3 основных волн миграции через Берингию, в ходе которых сюда последовательно проникли тропические элементы, обитатели лиственных лесов субтропического или умеренного пояса и виды холодоустойчивых мелколиственных лесов.
6. Формирование комплекса хвоегрызущих коконопрядов шло вместе с интенсивным видообразованием хвойных в позднемеловом периоде в ходе поднятия горной складчатости и общего похолодания климата. Оно хорошо увязывается с флорогенезом хвойных на территории современного Китая, считающегося центром их происхождения, что дает возможность рассматривать этот регион и в качестве центра происхождения большинства родов коконопрядов хвоегрызущей группы.
7. Комплекс эремиальных коконопрядов Старого Света имеет двойственное происхождение и раннеплейстоценовый возраст. Обитатели оазисов и тугайных лесов представлены дериватами фауны тропических лесов и связаны с древесной растительностью. Собственно эремиальные виды возникли на окраинах ареалов своих тропических предков, перешедших к обитанию в саванновых или степных биотопах. Они освоили кустарниковую и кустарничковую растительность, а также приспособились к питанию широким спектром травянистой растительности, продвигаясь вслед за ней даже в экстремально аридные зоны и претерпевая значительные морфологические, экологические и этологические изменения.
8. Современное распространение отдельных подсемейств и триб коконопрядов сильно отличается от исходного. Европейское происхождение имеют небольшое примитивное подсемейство Poecilocampinae и Malacosominae, давшие вторичные центры видообразования в горах Китая и в Сонорской области. Африканскими по происхождению являются своеобразные реликтовые подсемейства Chondrosteginae и Chionopsychinae. Наиболее многочисленны и широко распространены трибы Pinarini и Selenepherini, сформировавшиеся, вероятно, на Азиатском субконтиненте, но впоследствии проникшие в Австралию.
9. Коконопряды, как группа с низкой летной активностью, характеризуются высоким уровнем островного эндемизма и множеством викарных видов. Этот уровень заметно ниже для островов материкового происхождения, где эндемики приурочены в основном к горным цепям. Для островов смешанного происхождения показатель эндемизма выше, а острова океанического происхождения имеют самый высокий уровень видового эндемизма при более низком уровне видового обилия.
Материалы диссертации опубликованы в следующих работах изданий из списка перечня ВАК
- Стекольников А.А., Золотухин В.В. Филогенетическое положение подсемейств палеарктических коконопрядов (Lepidoptera, Lasiocampidae) с учетом строения скелета и мускулатуры гениталий самцов // Вестник Санкт-Петербургского ун-та. – 1994. – Сер. 3, вып. 1. – С. 26-32.
- Стекольников А.А., Золотухин В.В. Филогенетическое положение подсемейств палеарктических лемониид (Lemoniidae, Lepidoptera) с учетом строения скелета и мускулатуры гениталий самцов // Энтомол. обозр. – 2002. – T. 83, вып. 3. – С. 693-671.
- Золотухин В.В. Особенности островного эндемизма коконопрядов (Lepidoptera, Lasiocampidae) // Зоол. ж. – 2009. – T. 88, № 1. – С. 35-46.
- Chistjakov Yu.A., Zolotuhin V.V. Species of Lasiocampidae (Lepidoptera) of Russia and Adjacent Countries // Entomological Review – 1995. – T. 74. – P. 69-78.
- Stekolnikov A.A., Zolotuhin V.V. Phylogenetic relationships and Systematic Position of the Family Lemoniidae (Lepidoptera) with Account of Skeleton Structure and Muscles of Male Genitalia // Entomological Review – 2002. – T. 82, № 6. – P. 680-685.
- Zolotuhin V.V. New and Little-known Species of the Genus Phyllodesma Hbn. (Lepidoptera Lasiocampidae) // Entomological Review – 1995. – T. 74, № 1. – P. 33-40.
- Zolotuhin V.V. New and Little-known Lepidoptera Heterocera (Lasiocampidae, Nolidae) from Russia and Adjacent Territories // Entomological Review –1995. – T. 74, № 8. – P. 84-93.
- Zolotuhin V.V. Peculiarities of Island Endemism in Lasiocampidae (Lepidoptera) // Entomological Review – 2009. – T. 89, № 1. – Р. 34-45.
- Zolotuhin V.V., Zahiri R. The Lasiocampidae of Iran (Lepidoptera) // Zootaxa – 2008. – № 1791. – P. 1-52.
Монографии и статьи в коллективных монографиях
- Золотухин В.В. Разноусые чешуекрылые Вьетнама. Сем. Коконопряды (Lepidoptera: Lasiocampidae). – 2009. – Ульяновск: Изд-во КТП. – 128 с.
- Zolotuhin V.V., Pinratana A. The Moths of Thailand. Vol. 4. Lasiocampidae. – 2005. – Bangkok, Publ. Bros St. Gabriel. – 205 p., 28 pls.
- Золотухин В.В. Сем. Lasiocampidae. B: Синев С.Ю. (ред.). Каталог чешуекрылых (Lepidoptera) России. – 2008. – СПб.; М.: Тов-во научн. изд. КМК.– 424 с.
- Zolotuhin V.V. Lasiocampidae (Lepidoptera: Lasiocampoidea) // Esperiana. – 2007. – Memoir 4. – P. 205-214.
Прочие статьи
- Золотухин В.В. К систематическому положению коконопрядов рода Phyllodesma Hbn. (Lepidoptera, Lasiocampidae) Сибири // Вестник Ленинград. ун-та – 1991. – Сер. 3, № 17. – С. 126-129.
- Золотухин В.В. Коконопряды группы franconicum (Malacosoma; Lasiocampidae, Lepidoptera) фауны СССР. Сообщение 1. // Вестник Санкт-Петербургского ун-та. – 1992. – Сер. 3, № 1. – C. 25-33.
- Золотухин В.В. O Diplura argentifera Hering (Lepidoptera, Lasiocampidae) // Вестник Санкт-Петербургского ун-та. – 1993. – Сер. 3, вып. 2. – C. 108-110.
- Золотухин В.В. Новые и малоизвестные коконопряды рода Phyllodesma Hbn. (Lepidoptera, Lasiocampidae) // Энтомол. обозр. – 1994. – T. 73, вып. 1. – C. 136-143.
- Золотухин В.В. Желтые шелкопряды Кавказа (Lemoniidae, Lepidoptera) // Зоол. ж. – 1994. – T. 73, № 2. – С. 94-101.
- Золотухин В.В. Новые и малоизвестные разноусые чешуекрылые (Lepidoptera: Heterocera, Lasiocampidae, Nolidae) фауны России и сопредельных территорий. // Зоол. ж. – 1994. – T. 73, № 10. – С. 62-71.
- Чистяков Ю.А., Золотухин В.В. Коконопряды рода Poecilocampa Steph. (Lepidoptera, Lasiocampidae) России и сопредельных территорий // Энтомол. обозр. – 1994. – T. 73, вып. 2. – C. 321-330.
- Гуркович А.В., Золотухин В.В. Обзор коконопрядов рода Pehria Strand (Lepidoptera, Lasiocampidae) // Труды Русского энтомологического общества, С. Петербург. –2010. – Т. 80. №2. – С. 16–22.
- Ефимов Р.В., Золотухин В.В., Дёмин А.Г., Кнушевицкая М.В. Филогенетические взаимоотношения внутри рода Euthrix Meigen, 1830 (Lepidoptera: Lasiocampidae: Pinarinae: Cosmotrichini) на основе анализа нуклеотидного полиморфизма митохондриального гена цитохромоксидазы 1 (COI) // Энтомологические и паразитологические исследования в Поволжье, Саратов. – 2007. – Вып. 6. – С. 22–32.
- Прозоров А.М., Золотухин В.В. Выделение неотипа Opisthodontia dentata Aurivillius, 1899 (Lepidoptera, Lasiocampidae) фауны Африки с описаниями нового рода и нового вида // Труды Русского энтомологического общества, С. Петербург. –2010. – Т. 80. №2. – С. 143–48.
- Сергеев А.И., Золотухин В.В. О таксономическом объеме вида Lasiocampa quercus L. (Lepidoptera: Lasiocampidae) // Эверсманния. – 2008. – Вып. 13-14. – С. 5–17.
- Dubatolov V.V., Zolotuhin V.V. A list of the Lasiocampidae (Lepidoptera) from the territory of the former USSR // Atalanta. – 1992. – Vol. 23. – P. 531–548.
- Gurkovich A.V., Zolotuhin V.V. Mariaeia Dufrane, 1945, a lasiocampid genus misplaced in the Chrysopolomidae (Insecta: Lepidoptera) // Entomofauna. – 2009a. - Vol. 30. – P. 289–300.
- Gurkovich A.V., Zolotuhin V.V. Revision of Odontocheilopteryx Wallngr in Africa (Lepidoptera, Lasiocampidae) // Neue entomol. Nachr. – 2009b. – Vol. 63. – P. 77–111.
- Kostjuk I.Yu., Zolotuhin V.V. To a fauna of the mongolian Phyllodesma Hbn. (Lepidoptera, Lasiocampidae) // Atalanta. – 1994. – Vol. 25. – P. 297–305.
- Sergeev A.I., Zolotuhin V.V. A review of the genus Odonestis Germar, 1812 with descriptions of two new species and a new subspecies (Lepidoptera, Lasiocampidae) // Atalanta. – 2010. – Vol. 41. – P. 375–396.
- Zolotuhin V.V. On new and little-known Lasiocampidae (Lepidoptera) from Armenia, USSR // Atalanta. – 1991. – Vol. 22. – P. 117–123.
- Zolotuhin V.V. An annotated checklist of the Lasiocampidae (Lepidoptera) of Caucasus // Atalanta. – 1992. – Vol. 23. – P. 225–243.
- Zolotuhin V.V. An annotated checklist of the Lasiocampidae of the Russian Far East // Atalanta. – 1992. – 23. – P. 499–517.
- Zolotuhin V.V. An annotated checklist of the Lasiocampidae of “European Russia” // Atalanta. – 1992. – 23. – P. 519–529.
- Zolotuhin V.V. Baodera gen. nov. for Trichiura khasiana Moore (Lepidoptera, Lasiocampidae) // Atalanta. – 1992. – 23. – P. 491–493.
- Zolotuhin V.V. On Bhima potanini Alpheraky, 1895, as one of the underservedly forgotten species of Lasiocampidae (Lepidoptera) // Atalanta. – 1992. – 23. – P. 219–224.
- Zolotuhin V.V. On the types of Lasiocampidae (Lepidoptera) described by F. Bryk // Atalanta. – 1992. – 23. – Vol. 495–498.
- Zolotuhin V.V. An annotated checklist of the Lasiocampidae of Kazakhstan and Middle Asia (Lepidoptera) // Atalanta. – 1995. – Vol. 26. – Vol. 273–290.
- Zolotuhin V.V. To a study of asiatic Lasiocampidae (Lepidoptera). 1. The Lasiocampidae of Thailand // Japan Heterocerists` Journal. – 1995. – Vol. 14. – P. 157–170.
- Zolotuhin V.V. Notes on Chinese Lepidoptera (Lasiocampidae, Endromididae, Bombycidae) with a description of a new species // Nachr. entomol. Ver. Apollo. – 1996. – Vol. 16. – P. 373–386.
- Zolotuhin V.V. To a study of asiatic Lasiocampidae (Lepidoptera). 2. On the status of Stenophylloides javanus Draeseke, 1941 // Atalanta. – 1996. – Vol. 27. – P. 335–338.
- Zolotuhin V.V. To a study of asiatic Lasiocampidae. 3. Short taxonomic notes on Genus Paralebeda Aurivillius, 1894 (Lepidoptera) // Entomofauna. – 1996. – Vol. 17. – P. 245–256.
- Zolotuhin V.V. Further synonymic notes in the Lasiocampidae (Lepidoptera) with description of a new Euthrix-species // Entomofauna. – 1998. – Vol. 19. – P. 53–76.
- Zolotuhin V.V. Bhima Moore, 1888 is a junior subjective synonym of Pyrosis Oberthuer, 1880 (Lepidoptera, Lasiocampidae) // Atalanta. – 1999. – Vol. 29. – P. 283–284.
- Zolotuhin V.V. To a study of Asiatic Lasiocampidae (Lepidoptera). 4. Genus Micropacha Roepke, 1953 // Tinea. – 2000. – Vol. 16. – P. 151–160.
- Zolotuhin V.V. Contributions to the study of Asiatic Lasiocampidae. 5. Descriptions of new species of Euthrix Meigen, 1830 and of related genera, with a synonymic note (Lepidoptera, Lasiocampidae) // Atalanta. – 2001. – Vol. 32. – P. 453–471.
- Zolotuhin V.V. Studies on Asiatic Lasiocampidae (Lepidoptera). 6. Descriptions of new species from India, China and Thailand with further synonimic notes on some Chinese taxa // Ent. Z. – 2002. – Vol. 112. – P. 135–140.
- Zolotuhin V.V. Contributions to the study of the Asiatic Lasiocampidae (Lepidoptera). 7. // Atalanta. – 2005. – Vol. 36. – P. 551–558.
- Zolotuhin V.V. To a knowledge of the Gastropacha Ochs., 1810 species of the Stenophylloides Hmps. [1893] 1892 group (Lepidoptera, Lasiocampidae) // Tinea. – 2005. – Vol. 18. – P. 291–306.
- Zolotuhin V.V. An annotated distributional check-list of the genus Trichiura Stephens, 1828 with description of six new taxa, especially from Asia Minor (Lepidoptera: Lasiocampidae) // Nachr. ent. Ver. Apollo. – 2007. – Suppl. 19. – P. 37–48.
- Zolotuhin V.V. Lasiocampidae (Lepidoptera: Lasiocampoidea) // Esperiana, Memoir. – 2007. – Band 4. – P. 205–214.
- Zolotuhin V.V. On the identity of the Turkish species of the Eriogaster rimicola-group (Lasiocampidae) // Nota lepidopterologica. – 2007. – Vol. 30. – P. 45–50.
- Zolotuhin V.V. A review of the genus Chionopsyche Aurivillius, 1909 (Lepidoptera, Lasiocampidae: Chionopsychinae) with description of a new species // Atalanta. – 2010. – Vol. 41. – P. 361–366.
- Zolotuhin V.V., Dubatolov V.V. A new species of Lasiocampidae (Lepidoptera) from Tajikistan // Atalanta. – 1992. – Vol. 23. – P. 215–217.
- Zolotuhin V.V., Gurkovich A.V. A review of the genus Pachypasa Walker, 1865, sensu lato in Africa (Lepidoptera, Lasiocampidae) // Neue entomologische Nachrichten. – 2009. – Vol. 63. – P. 1–72.
- Zolotuhin V.V., Gurkovich A.V. Synonymic notes on African Lasiocampidae (Insecta: Lepidoptera) // Entomofauna. – 2009. – Vol. 30. – P. 273–288.
- Zolotuhin V.V., Gurkovich A.V. Unusual case of gynandromorphism in Mimopacha knoblauchi Dewitz, 1881 with preliminary remarks on sex determination in the Lasiocampidae (Lepidoptera) // Tinea. – 2008. – Vol. 20. – P. 148–151.
- Zolotuhin V.V., Hauenstein A. A new species of Euthrix Meigen, 1820 from China (Lepidoptera: Lasiocampidae) // Nachr. ent. Ver. Apollo. – 2007, Suppl. 19. – P. 35–36.
- Zolotuhin V.V., Holloway J.D. The Lasiocampidae of Sulawesi // Tinea. – 2006. – Vol. 19. – P. 244–259.
- Zolotuhin V.V., Ihle S., Tannert R., Sinjaev V.V. A first case of gynandromorphism in Trabala Walker (Lepidoptera, Lasiocampidae) // Tinea. – 2008. – Vol. 20. – P. 146–147.
- Zolotuhin V.V., Ihle S. The Lasiocampidae (Lepidoptera) of Laos // Tinea. – 2008. – Vol. 20(4). – P. 217–235.
- Zolotuhin V.V., Kostjuk I.Yu. Phantosoma witti gen. et. sp. nov., a new autumn lasiocampid moth from Turkmenistan (Lasiocampidae) // Nota lepid. –2000. – Vol. 23(2). – P. 141–146.
- Zolotuhin V.V., Kurshakov P.A. Fine sculpture and phylogenetic implications of egg shell morphology in the Lasiocampidae (Lepidoptera) // Neue entomologische Nachrichten. – 2011. – Vol. 67. – P. 3-21.
- Zolotuhin V.V., Perekrasnov A.A. A review of the Euthrix laeta (Walker, 1855) complex with a description of a new species and two new subspecies (Lepidoptera, Lasiocampidae) // Atalanta. – 2010. – Vol. 41. – P. 367–374.
- Zolotuhin V.V., Prozorov A.M. A review of the genera Opisthodontia Aurivillius, 1895, and Stenophatna Aurivillius, 1909, with erection of 8 new genera and descriptions of 37 new species and 2 new subspecies (Lepidoptera: Lasiocampidae) // Atalanta. – 2010. – Vol. 41. – P. 397–460.
- Zolotuhin V.V., Saldaitis A., Reza Zahiri. Two new mountain species of the genus Poecilocampa Stephens 1828 (Lasiocampidae) // Tinea. – 2010. – Vol. 21. – P. 88–94.
- Zolotuhin V.V., Saldaitis A. A new species of Trichiura Stephens, 1828 (Lepidoptera, Lasiocampidae) from Lebanon // Entomofauna. – 2008. – Vol. 30. – P. 53–60.
- Zolotuhin V.V., Sinyaev V.V. A new genus and 2 new species of the Lasiocampidae from Vietnam (Lepidoptera) // Entomofauna. – 2009. – Suppl. 16. – P. 273–280.
- Zolotuhin V.V., Sinyaev V.V. The Lasiocampidae (Lepidoptera) of Laos. Part 2 // Neue entomologische Nachrichten. – 2011. – Vol. 67. – P. 37-39.
- Zolotuhin V.V., Solovyev A.V. Bombyx flavomarginata Poujade, 1886, – a limacodid species misplaced in the Lasiocampidae (Lepidoptera: Limacodidae) // Tinea. – 2008. – Vol. 20. – P. 98–101.
- Zolotuhin V.V., Tran Thieu Du. A new species of Dendrolimus Germar, 1812, from Vietnam (Lepidoptera, Lasiocampidae) // Tinea. – 2009. – Vol. 21. – P. 1–5.
- Zolotuhin V.V., Treadaway C.J., Witt Th.J. The Lasiocampidae (Lepidoptera) of the Philippines // Nachr. ent. Ver. Apollo. – 1998. – Suppl. 17. – P. 133–222.
- Zolotuhin V.V., Witt Th. Lasiocampidae of Nepal. Adenda and corrigenda to “Moths of Nepal”, Parts 1-5. // Moths of Nepal. Tinea. – 2000. – Suppl. Part 6. – P. 153–163.
- Zolotuhin V.V., Witt Th. The Lasiocampidae of Vietnam. // Entomofauna. – 2000. – Suppl. 11. – P. 25–104.
- Zolotuhin V.V., Witt Th. New and little-known species of the Lasiocampidae (Lepidoptera) from China. // Tinea. – 2004. – Vol. 18. – P. 36–42.
- Zolotuhin V.V., Witt Th. Contribution to the knowledge of Indonesian Lasiocampidae (Lepidoptera) // Tinea. – 2005. – Vol. 19. – P. 59–68.
- Zolotuhin V.V., Witt Th. A new genus and a new species of the Lasiocampidae from Pakistan (Lepidoptera: Lasiocampidae) // Nachr. ent. Ver. Apollo. – 2007. – Suppl. 19. – P. 32–34.
- Zolotuhin V.V., Witt Th. Revision of the genus Pyrosis Oberthr, 1880 (=Bhima Moore, 1888) (Lepidoptera: Lasiocampidae) // Nachr. ent. Ver. Apollo. – 2007. – Suppl. 19. – P. 1–31.
- Zolotuhin V.V., Wu Chunsheng, 2008. Three new species of the Lasiocampidae (Lepidoptera) from China // Tinea. – 2008. – Vol. 20. – P. 264–268,
а также 6 работ в сборниках тезисов и материалах международных и российских конференций. Общий список опубликованных по теме диссертации работ включает 84 источника и приведен в тексте диссертации полностью.