

Эволюция формы и функции раковины головоногих моллюсков подкласса coleoidea
На правах рукописи
БИЗИКОВ Вячеслав Александрович
ЭВОЛЮЦИЯ ФОРМЫ И ФУНКЦИИ РАКОВИНЫ
ГОЛОВОНОГИХ МОЛЛЮСКОВ ПОДКЛАССА COLEOIDEA
03.00.08 – зоология
А В Т О Р Е Ф Е Р А Т
диссертации на соискание ученой степени
доктора биологических наук
Москва-2008
Работа выполнена во Всероссийском научно-исследовательском институте рыбного хозяйства и океанографии
Официальные оппоненты:
доктор биологических наук, профессор; член-корреспондент РАН
Малахов Владимир Васильевич,
Московский государственный университет имени М.В. Ломоносова, Биологический факультет
доктор геологических наук, профессор
Барсков Игорь Сергеевич,
Московский государственный университет имени М.В. Ломоносова, Геологический факультет
доктор биологических наук
Кантор Юрий Израилевич,
Институт проблем экологии и эволюции имени А.Н. Северцова РАН
Ведущая организация: Зоологический институт РАН
Защита состоится 24 ноября 2008 г. в 15.30 на заседании диссертационного совета Д 501.001.20 при Биологическом факультете МГУ имени М.В. Ломоносова по адресу: 119991, Москва, ГСП-1, Ленинские горы, МГУ имени М.В. Ломоносова, Биологический факультет, ауд. М1
С диссертацией можно ознакомиться в библиотеке Биологического факультета МГУ имени М.В.Ломоносова.
Автореферат разослан 20 сентября 2008 г.
Ученый секретарь
диссертационного совета,
кандидат биологических наук Л.И. Барсова
I. ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Внутрираковинные головоногие моллюски, объединяемые в подкласс Coleoidea, представляют собой многочисленную, разнообразную и процветающую группу. Их современная фауна насчитывает около 800 видов, которые группируются, согласно принятой в настоящее время системе, в 6 отрядов: кальмаров, каракатиц, спирул, сепиолид, вампиров и осьминогов. Учитывая, что наружнораковинные головоногие ныне – это единственный род Nautilus с 8-10 видами, современный этап эволюционной истории Cephalopoda по праву можно назвать эпохой Coleoidea. Колеоидеи широко освоили Мировой океан, встречаясь от уреза воды до открытой океанской пелагиали, от поверхности до ультраабиссали, от экватора до Арктики и Антарктики. Несомненным признаком биологического успеха колеоидей служит тот факт, что многие из них занимают высшие трофические уровни в океанических экосистемах, достигают исключительно высокой численности и биомассы. Coleoidea – весьма древняя группа с богатой эволюционной историей. Первые внутрираковинные головоногие известны с конца девона, а в целом их ископаемая фауна насчитывает 7-9 отрядов, наиболее многочисленными и известными из которых являются белемниты.
Как часто случается в малакологии, исследователи ископаемых и современных головоногих «говорят на разных языках». Вся систематика и филогения вымерших цефалопод строится на признаках строения их раковины, поскольку только она обычно и сохраняется в ископаемом состоянии [Jeletzky, 1966]. В свою очередь, неонтологи строят систему рецентных форм в основном по признакам мягкого тела: строению глаз, ловчего аппарата, мантии, плавников и пр. [Young, Vecchione, 1996]. В результате палеонтологи, зная в деталях строение ископаемых раковин, имеют самое смутное представление об их функциональной роли и взаимоотношении с мягким телом, а неонтологи расходятся во мнениях относительно примитивного–продвинутого состояния признаков мягкого тела и ставят под сомнение саму возможность построения непротиворечивой системы и филогении современных головоногих [Несис, 1996; Young et al., 1998]. Очевидно, что создание единой филогенетической системы ископаемых и рецентных колеоидей невозможно без глубокого изучения морфологии и функции раковины современных форм. Лишь конкретное понимание роли раковины и ее отдельных частей позволит отличить гомологическое сходство от часто встречающегося у головоногих конвергентного сходства. Однако морфология и функциональная роль раковины современных головоногих до сих пор остаются весьма слабо изученными.
Первая и наиболее последовательная попытка построения единой филогенетической системы Coleoidea на основе строения раковины ископаемых и рецентных форм была предпринята более 80 лет назад Адольфом Нэфом [Naef, 1921/1923]. За десятилетия, прошедшие со времени ее появления, система Нэфа неизбежно устарела. Один из существенных недостатков работ Нэфа – отсутствие в них функционального анализа морфологии раковины рецентных видов. Недостаток этот вполне объясним: во времена Нэфа об образе жизни большинства головоногих моллюсков не было известно практически ничего.
Это определило цель исследования: всестороннее изучение функциональной морфологии раковины современных головоногих моллюсков подкласса Coleoidea и ее взаимоотношения с мышечной системой этих животных.
Для достижения цели решали следующие задачи:
1) Исследование морфологии раковины и ее взаимоотношений с мягким телом у представителей всех основных групп современных головоногих.
2) Исследование функциональной роли раковины и её отдельных структурных элементов у современных представителей подкласса Coleoidea.
3) Анализ гомологий раковин различных групп современных и ископаемых Coleoidea.
4) Исследование филогенетических связей современных и ископаемых колеоидей на основании строения их раковины.
5) Анализ общих закономерностей взаимоотношения мягкого тела и раковины при переходе последней из наружного состояния во внутреннее.
Научная новизна. Разработаны и применены новые методы резки и приготовления препаратов срезов раковин и тотальных срезов тела головоногих моллюсков. Разработана единая терминология раковины и с её помощью описана морфология раковины и её взаимоотношение с мягким телом у представителей всех подотрядов и надсемейств современных головоногих моллюсков, как у взрослых, так и у личинок. Впервые проведен функциональный анализ раковины и способов прикрепления к ней мускулатуры. Составлена морфологическая классификация раковин современных колеоидей и выделено 12 морфологических типов раковины. Показано, что раковины некоторых морфологических типов (спирулидный, сепиидный, сепиолидный, цирратный, инцирратный, хиротеутидный, тизанотеутидный, оммастрефидный, кранхиидный) хорошо различаются уже на личиночных стадиях, а раковины других типов (вампиротеутидный, лолигинидный и онихотеутидный) на личиночных стадиях сходны между собой). Проведен анализ морфологического разнообразия плавников современных колеоидей и предложена новая их классификация на основе новых морфо-функциональных критериев. Предложены новые, морфо-функциональные критерии анализа структуры раковины и на их основе впервые проведено гомологическое сопоставление структурных элементов раковин всех основных групп рецентных Coleoidea. Проведен филогенетический анализ основных групп рецентных и ископаемых внутрираковинных головоногих моллюсков на основе строения их раковины и предложена новая система подкласса Coleoidea.
Теоретическое и практическое значение работы. На основании анализа раковин современных и ископаемых колеоидей сделаны общие заключения о перестройке плана строения головоногих моллюсков при погружении их раковины внутрь тела, о смене функций и структуры раковины в ходе её трансформации из наружной во внутреннюю. Выявлено два типа контактов мышц с раковиной у колеоидных головоногих моллюсков – первичные и вторичные контакты – и показана ключевая роль хрящевой ткани в осуществлении контактов вторичного типа (прикрепление мышц к внешней поверхности раковины). Предложена новая гипотеза о происхождении плавников колеоидей, учитывающая роль хряща в формировании плавников и их сочленении с раковиной. Выявлены общие тенденции изменения морфологии раковины в эволюции колеоидей, показана функциональная обусловленность утраты раковины в отдельных ветвях. Показана функциональная связь между декальцинацией раковины и утратой ею гидростатической функции у колеоидных головоногих. Установлено, что гладиус – внутренняя декальцинированная раковина без фрагмокона – появлялся неоднократно и независимо в различных эволюционных ветвях колеоидей: спирулоидной, сепиоидной, теутоидной и вампироподной. Проведен филогенетический анализ основных групп рецентных и ископаемых Coleoidea на основе строения их раковины. Показано, что кальмары в их современном понимании являются не систематической, а экологической категорией, не таксоном, а жизненной формой. Предложена новая система подкласса Coleoidea. Показано, что эволюция локомоторного аппарата Coleoidea в основных чертах повторяла принципиальные морфо-функциональные решения, реализованные в эволюции локомоторного аппарата примитивных бесчелюстных панцирных хордовых, предков рыб, однако у головоногих она проходила на основе иного – моллюскового – плана строения.
Основные положения, выносимые на защиту:
- Морфология мускульной системы головоногих моллюсков в эволюционном отношении более консервативна, чем морфология их раковины. В связи с этим план мускулатура может использоваться в качестве системы координат при гомологическом анализе раковины Coleoidea.
- Кальмары – головоногие с декальцинированной раковиной (гладиусом) – являются не систематической категорией, а определенным этапом параллельной эволюции различных ветвей колеоидей, наступающим после редукции в раковине газового фрагмокона.
- Обызвествление раковины Coleoidea функционально связано с ее гидростатической функцией. Декальцинация раковины свидетельствует об утере ею гидростатической функции (и, соответственно, фрагмокона).
Апробация работы. Материалы диссертации доложены на международной конференции американского малакалогического общества в Санта-Барбаре (США) в 1996 г., на международных симпозиумах Совета по изучения головоногих моллюсков (CIAC) в Мадзара-дель-Вало (Италия) в 1989 г. и в Абердине (Великобритания) в 2000 г, на симпозиуме ИКЕС «Биология популяций промысловых беспозвоночных» (Канада, 1990), на всероссийских конференциях в Зоологическом Институте РАН в Санкт-Петербурге: в 2003г. и в 2006г., на научных семинарах Кафедры сравнительной анатомии и зоологии беспозвоночных Биологического факультета МГУ, Института зоологии им. И.И. Шмальгаузена НАН Украины, Лаборатории промысловых беспозвоночных ВНИРО, на заседаниях Ученого Совета ВНИРО.
Благодарности. Я глубоко благодарен своим друзьям и коллегам, в творческом общении с которыми постепенно складывалась эта работа: К.Н. Несису, Я.И. Старобогатову, Ч.М. Нигматуллину, А.И. Архипкину, Б.Г. Иванову, Р.М. Сабирову, Л.А. Догужаевой, Ю.А. Филипповой, А.А. Нейман, Д.Н. Хромову, Д.О. Алексееву, О.Н. Катугину, М.А. Рогову, R.E. Young (Гонолулу, США), S.von Boletzky (Париж, Франция), D.T. Donovan (Лондон, Великобритания), F.G. Hochberg (Санта-Барбара, США), T. Kubodera (Токио, Япония). Выражаю искреннюю признательность Т. Линьковскому (Гдыня; Польша) за предоставленную возможность работы с коллекциями личинок головоногих моллюсков из планктонных сборов Мексиканского залива.
Публикации. По теме диссертации опубликовано 46 работ, в том числе четыре монографии (одна – в соавторстве), 28 статей (из них 9 – в рецензируемых журналах, поименованных в списках ВАК), 13 тезисов и материалов конференций, одно авторское свидетельство.
Структура работы. Работа состоит из двух томов. 1-й том – текст диссертации (408 стр.) – содержит введение, 10 глав, заключение и выводы. 2-й том - иллюстрации (186 стр.) – содержит 212 рисунков и 1 таблицу. Список цитируемой литературы включает 252 наименования, в том числе 48 работ на русском и 204 – на иностранных языках.
II. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
В основу работы положены исследования, проводимые автором с 1984 г. Материалы для работы собраны в экспедициях НПС «Одиссей» (1984/85), РПС «Гидробиолог» (1987), НИС «Пр. Водяницкий» (1990), а также в многочисленных рейсах на промысловых судах в Северной Пацифике и Юго-Западной Атлантике. Также использованы материалы коллекций головоногих моллюсков, собранные во ВНИРО, АтлантНИРО, ИО РАН и в Зоологическом Музее МГУ. В общей сложности обработано 272 особи головоногих и их раковин, в том числе представителей 22 семейств кальмаров, 2 семейств плавниковых осьминогов (п/отр. Cirrata) и 7 семейств бесплавниковых (п/отр. Incirrata), вампиротеутиса (Vampyroteuthis), каракатицы (Sepia), сепиолиды (Rossia), спирулы (Spirula) и наутилуса (Nautilus). Для изучения внутренней структуры гладиусов выполняли их срезы различной ориентации по оригинальной методике: гладиус зажимали между двух кусочков пенопласта и резали вручную микротомным ножом [Бизиков, 1990; 1996]. Взаимоотношение раковины и мягкого тела исследовано по тотальным срезам предварительно замороженных животных. Как правило, выполняли один тотальный сагиттальный срез и 5-6 поперечных срезов на следующих уровнях: воронка; средняя часть вороночных ретракторов; задние окончания вороночных ретракторов; передняя часть плавников; максимальная ширина плавников; задняя часть плавников. Тотальные срезы фотографировали или зарисовывали под бинокулярным микроскопом; полученные изображения сканировали и монтировали с использованием графических редакторов.
III. СТРОЕНИЕ РАКОВИНЫ NAUTILUS И ЕЕ ВЗАИМООТНОШЕНИЕ
С МЯГКИМ ТЕЛОМ
9-10 видов рода Nautilus, встречающихся в морях Индо-Вестпацифики, являются единственными выжившими до наших дней представителями наружнораковинных головоногих моллюсков (подкласс Ectocochlea) - древней группы, история которой насчитывает свыше 500 млн. лет. Наутилус сохранил удивительный набор архаичных признаков строения раковины и мягкого тела, из-за чего это животное часто называют «живым ископаемым». Организация наутилуса служит отправной точкой отсчета в филогенетических и сравнительно-морфологических исследованиях головоногих моллюсков. Если бы наутилус не дожил до наших дней, филогения Coleoidea в гораздо меньшей степени поддавалась бы расшифровке методами сравнительной анатомии.
Функциональная морфология, гистология и работа мускулатуры наутилуса исследована мной на примере 4 экземпляров Nautilus pompilius Linnaeus, 1758, пойманных у о. Минданао (Филиппины). В ходе гистологических исследований обнаружены ранее неизвестные парные мускулы – затылочные ретракторы – отходящие от задних окончаний воротниковых складок к дорсальной стенке жилой камеры и по-видимому участвующие в респираторном цикле. Проанализирована работа мускулатуры во время дыхания и гидрореактивного движения наутилуса и показано наличие функционального конфликта между этими процессами. Тотальные срезы наутилуса, выполненные на уровне затылочных клапанов показали, что жабры наутилуса, поддерживаемые навесу мягкими осевыми хрящами, могут находиться в рабочем положении (напротив отверстий воротниковых карманов) лишь при слабых респираторных токах воды. Сильные токи и высокое давление, создаваемые при гидрореактивном толчке, прижимают жабры к вентральной стенке мантийной полости и выключают их из работы. Неспособность жабр наутилуса работать при высоких давлениях стала слабым местом его респираторной и локомоторной систем, сделала невозможным интенсификацию дыхания и движения. По-видимому, эволюционным решением проблемы «плыть или дышать» стало развитие у наутилуса особого вентиляционного режима, сочетающего дыхание с медленным, но продолжительным реактивным плаванием. Режим быстрого реактивного плавания остался для него резервом, используемым лишь в экстренных ситуациях.
IV. СТРОЕНИЕ РАКОВИНЫ И ЕЕ ВЗАИМООТНОШЕНИЕ С МЯГКИМ
ТЕЛОМ У РАЗНЫХ ГРУПП ПОДКЛАССА COLEOIDEA
Даны подробные описания морфологии, микроструктуры раковины и взаимоотношения с мягким телом у представителей 19 семейств современных колеоидей. По результатам описания для каждой группы составлялась схема раковины, на которой указывались зоны прикрепления к ней различных мышц и органов. При этом выявлен ряд новых фактов, касающихся строения раковины и мягкого тела ряда групп современных колеоидей, в том числе:
Спирула (Spirula spirula Linnaeus, 1758). Раковинный мешок спирулы усилен слоем хряща, окружающего раковину плотным футляром. Именно к этому хрящу, а не к самой раковине, прирастают плавники, мантия и ретракторы головы. Все мышцы локомоторного аппарата спирулы прикрепляются либо к ее наружной поверхности (мантия, плавники), либо к краям устья (ретракторы головы). К внутренней дорсальной стенке жилой камеры раковины прикрепляется лишь лигамент, поддерживающий пищеварительную железу. В мантийной полости спирулы, на вентральной стороне от ретракторов головы обнаружено выпячивание висцерального мешка, вмещающее нижнюю полую вену, прямую кишку и проток чернильного мешка.
Каракатица (Sepia officinalis Linnaeus, 1758). Исследование декальцинированных раковин каракатиц показало, что основу их дорсального щита составляет тонкий хитиновый слой, видимый по краям раковины в виде каймы и гомологичный проостракуму других колеоидей. Он состоит из трех продольных элементов: медиальной пластинки (рахиса) и пары латеральных пластинок, разделенных нечеткими асимптотами. По обе стороны от хитинового слоя расположены два обызвествленных слоя: наружный бугорчатый и внутренний, формирующий видоизмененный фрагмокон. Анализ расположения зон крепления мышц к сепиону показывает, что сепион в качестве опорной структуры представляет собой типичный проостракум, только спрятанный под модифицированным фрагмоконом.
Тихоокеанская рссия (Rossia pacifica Berry, 1911). Гладиус Rossia сложен двумя раковинными слоями: средним (хитиновым) и внутренним (хрящеподобным). Проостракум не доходит до заднего края гладиуса. Задний конец проостракума, отмеченный схождением асимптотических линий, заканчивается на расстоянии примерно 1/5 длины гладиуса от его заднего конца. Это место соответствует апикальной вершине конуса других колеоидей. Конус развернут в плоскость и редуцирован до плоской узкой и тонкой пластинки в задней части гладиуса. Задний край вентральной стенки конуса имеет форму вилки с узкими лучами, расставленными под углом около 90. Гладиус Rossia на всем протяжении заключен в толстый хрящевой футляр, образованный стенками раковинного мешка и служащий для крепления мышц локомоторного аппарата.
Вампиротеутис (Vampyroteuthis infernalis Chun, 1903). Впервые показано трехслойное строение гладиуса вампира, трехмерная форма гладиуса, наличие вентрального выреза в стенке конуса. Выяснено, что проостракум вампира состоит из пяти продольных элементов: медиальной пластинки (рахиса), парных латеральных пластинок и крыльев. Показано, что сочленение плавников с телом у вампиротеутиса сходно с таковым у большинства колеоидей: основания плавников сочленены с телом подвижно, через базальные эпителиальные мешки и мускулы-регуляторы.
Калифорнийский опистотеутис (Opisthoteuthis californiana Leach, 1818). Подробно описано строение гладиуса опистотеутиса и крепление к нему мышц. Гладиус представляет собой толстую подковообразную структуру, лежащую поперек мантии. Выяснено, что гладиус опистотеутиса и других цирратных осьминогов образован одним раковинным слоем, сложенным многочисленными концентрическими слоями хрящеподобного вещества, по микроструктуре весьма сходного с внутренним раковинным слоем гладиуса вампиротеутиса и кальмаров. Крупные крыловидные плавники цирратных осьминогов плотно прирастают основаниями к внешней поверхности расширенных боковых участков гладиуса. Базальные эпителиальные мешки отсутствуют. Плотное прирастание плавников к раковинному мешку, вероятно, обусловлено развитием нового способа плавания: подводного полета с помощью взмахов крыловидных плавников, при одновременной редукции мантийно-вороночного реактивного аппарата.
Осьминог-цирротеутис Cirroteuthis muelleri Eschricht, 1836. Исследование тотальных поперечных срезов тела цирротеутиса показало, что висцеральный мешок у этого вида срастается с боковыми стенками мантии, что приводит к значительному сокращению объема мантийной полости. Базальные хрящи плавников, как и у опистотеутиса, прирастают к раковинному мешку. Воронка слабо мускулистая. Вороночные ретракторы слабые и крепятся к мантии, что указывает на неспособность этого животного к реактивному движению.
Тихоокеанский осьминог (Enteroctopus dofleini (Wulker, 1910)). Описана морфология и микроструктура рудиментов раковины тихоокеанского осьминога, представляющих собой пару хрящеподобных палочек, расположенных на дорсолатеральной стороне мантии и известных в литературе под названием стилеты. Стилеты образованы концентрическими слоями хрящеподобного раковинного вещества, по-видимому гомологичного материалу гладиусов цирратных осьминогов. Выявлена значительная случайная ассиметрия стилетов одной пары. Показано, что стилеты служат якорьками, удерживающими основания вороночных ретракторов. Мантийные мышцы прирастают к стилетам по их периферии, но не обрастают сверху.
Аргонавт (Argonauta nodosa Solander, 1786). Раковина аргонавтов редуцирована полностью. Пара косых рубчиков на дорсо-латеральных стенках в задней трети мантии указывает прежнее положение стилетов. В отсутствие стилетов крепление вороночных ретракторов к мантии стало слабее. Это ослабление компенсировано развитием сложного мантийно-вороночного замыкательного аппарата, фиксирующего положение воронки относительно тела. Важно отметить, что исчезновение раковины у аргонавтов не привело к существенному изменению их плана строения, который остался тем же, что и у других инцирратных осьминогов, имеющих стилеты.
Патагонский кальмар (Doryteuthis gahi d’Orbigny, 1835). Раковина представляет собой гладиус: хитиновую пластинку перовидной формы. Выяснено, что гладиус лолигинид в типичном случае образован тремя раковинными слоями: средним хитиновым и хрящеподобными наружным и внутренним. Проостракум лолигинид состоит из трех продольных элементов: медиальной пластинки (рахиса) и пары латеральных пластинок. Передний свободный отдел рахиса короткий (15-20% длины гладиуса). Конус и рострум редуцированы (из кальмаров-лолигинид рострум имеется только у рода Alloteuthis [Naef, 1921/1923]). Показано, что в местах прикрепления к гладиусу латеральные сегменты ретракторов головы у лолинид слиты с ретракторами воронки.
Лучеперый кальмар (Chtenopteryx sicula Verany, 1851). Гладиус лучеперого кальмара весьма сходен с гладиусами кальмаров-лолигинид внешней перовидной формой, трехчастным проостракумом, редуцированными конусом и рострумом. Отличием Chtenopteryx является более длинный передний свободный отдел рахиса (около 40% длины гладиуса). В задней части ретракторы воронки срастаются с ретракторами головы и имеют общее место прикрепления к вентральной стороне латеральных пластинок. Необычная дифференцировка плавников Chtenopteryx на мускульные лучи и кожистую перепонку по-видимому является адаптацией к жизни на больших глубинах: формирование мускульных лучей приводит к сокращению мышечной массы плавников, а следовательно, к сокращению потребности в кислороде и питании. Крепление плавников к мантии у Chtenopteryx сходно с таковым у лолигинид.
Глубоководный красно-бурый кальмар (Bathyteuthis abyssicola Hoyle, 1885). Гладиус сходен по строению с Chtenopteryx и кальмарами-лолигинидами. Основным отличием Bathyteuthis является сильное удлинение переднего свободного отдела рахиса (до 60% от длины гладиуса). Это удлинение увеличивает объем мантийной полости и позволяет кальмару делать быстрые короткие броски, но ослабляет способность к продолжительному скоростному плаванию. Гладиус сформирован средним (хитиновым) раковинным слоем; внутренний и наружный слои неразличимы. Проостракум состоит из трех элементов. Латеральные сегменты ретракторов головы в местах прикрепления к гладиусу слиты с ретракторами воронки. Плавники сочленены с мантией подвижно, посредством эпителиальных базальных мешков.
Кальмар-ромб (Thysanoteuthis rhombus Troschel, 1857). Гладиус имеет строение, уникальное для современных кальмаров. Его широкий копьевидный проостракум состоит из 7 продольных элементов: непарной медиальной (рахиса) и трех парных – гиперболических пластинок, параболических пластинок и крыльев. Конус мелкий, чашевидный. Рострум отсутствует. Из раковинных слоев наиболее развит средний (хитиновый) слой, образующий проостракум и конус. Внутренний слой ограничен задней третью гладиуса. Наружный слой отсутствует. Проостракум выступает внутрь мантийной полости и частично окружает висцеральный мешок. Жабры оказываются снаружи от гладиуса, в щели между параболическими пластинками и стенкой мантии. Зоны прикрепления мускулов-ретракторов к гладиусу далеко разнесены между собой: ретракторы головы крепятся к выступающим вперед параболическим пластинкам, а ретракторы воронки – к крыльям.
Тихоокеанский кальмар (Todarodes pacificus Troschel, 1857). Гладиус имеет характерную для оммастрефид мечевидную форму. В задней части имеется узкий «перехват» - стебель. Проостракум трехчастный, состоит в основном из рахиса, укрепленного тремя продольными ребрами жесткости. Латеральные пластинки трансформированы в утолщенные валики по бокам от рахиса. Конус небольшой, покрыт снаружи хрящеподобным колпачком, представляющим собой остаток альвеолы рострума. В местах прикрепления к гладиусу ретракторы головы не срастаются и даже не соприкасаются с ретракторами воронки. Крепление мантии к гладиусу повсеместно осуществляется через хрящевую ткань, которая в области стебля образует вокруг гладиуса замкнутый хрящевой чехол, плотно сцепленный с гладиусом благодаря глубоким пазам, идущим вдоль стебля. Основания плавников срастаются и крепятся к гладиусу подвижно, посредством общего базального плавникового хряща, профиль которого напоминает рыбий позвонок.
Гигантский тихоокеанский крючьеносный кальмар (Moroteuthis robusta (Verril, 1876)). Главная отличительная особенность гладиуса M. robusta – длинный хрящеподобный рострум, занимающий заднюю четверть раковины и по форме напоминающий морковку. Проостракум состоит из 5 продольных элементов: рахиса, парных латеральных пластинок и крыльев. Конус мелкий, вставленный в расширенное основание рострума (альвеолу). Гладиус образован 3 раковинными слоями: средним, наружным и внутренним. Рострум длинный, цилиндрический, прозрачный. Апикальная линия хорошо видна на просвет. Мантия и плавники крепятся к гладиусу посредством хорошо развитых хрящей. Латеральные сегменты ретракторов головы в местах прикрепления к гладиусу не срастаются с ретракторами воронки. «Хвост» на заднем конце плавников M. robusta поддерживается рострумом, окруженным толстым и плотным соединительнотканным чехлом.
Командорский кальмар (Berryteuthis magister (Berry, 1913)). Гладиус образован в основном средним (хитиновым) раковинным слоем; внутренний слой имеется в задней половине гладиуса, а наружный редуцирован до тонкого колпачка вокруг конуса. Проостракум состоит из пяти продольных элементов: рахиса, пары латеральных пластинок и пары крыльев. Рострум отсутствует. Латеральные сегменты ретракторов головы на всем протяжении не срастаются с ретракторами воронки. В области плавников мантия крепится к гладиусу посредством сложно устроенного мантийного хряща. Базальные хрящи плавников сращены в единый осевой плавниковый хрящ. Задний конец плавников поддерживается небольшим апикальным хрящом, являющимся задним продолжением плавникового хряща.
Кальмар-ликотеутис (Lycoteuthis diadema Chun, 1900). Проостракум состоит из трех продольных элементов: рахиса и пары латеральных пластинок. Конус мелкий и широкий, с едва заметным рострумом на апикальной вершине. В области прикрепления к гладиусу ретракторы воронки и латеральные сегменты ретракторов головы срастаются между собой, но не срастаются с медиальным сегментом ретракторов головы и с висцеральным мешком: между ними имеется узкая глубокая щель, почти доходящая до гладиуса. Задние окончания вороночных ретракторов закреплены обоими краями: внутренний край прирастает к латеральным пластинкам гладиуса, а внешний край – к мантийной стенке. Необычен способ прикрепления мантии к гладиусу: в задней части мантия входит в глубокие узкие щели, идущие вдоль краев гладиуса и играющие роль зажимов. Крепление плавников жесткое, без участия эпителиальных базальных мешков. Базальные хрящи плавников образуют плотный футляр, окружающий высокий медиальный киль гладиуса с дорсальной стороны. Задний конец плавников поддерживается апикальным плавниковым хрящом.
Большой биченосный кальмар (Mastigoteuthis magna Joubin, 1913). Гладиус длинный, веретеновидный. Проостракум состоит из трех продольных элементов: медиальной пластинки (рахиса) и пары латеральных пластинок. Конус очень длинный (34-35% длины гладиуса), игловидный, с невысоким дорсальным килем. Рострум отсутствует. Хрящевая ткань в местах прикрепления мантии к гладиусу развита слабо. Выяснено, что мантия не распространяется до заднего конца тела, а заканчивается на уровне переднего края конуса. Сам конус окружен студенистым соединительнотканным валиком вакуолизированной аммонийной ткани, играющим роль поплавка, функционального аналога фрагмокона. В местах прикрепления к гладиусу ретракторы головы и воронки не срастаются между собой. Висцеральный мешок с вентральной стороны от печени образует выпячивание, в котором помещаются прямая кишка, нижняя полая вена, чернильный мешок и фотофоры. Плавники прикрепляются к гладиусу жестко; базальные эпителиальные мешки сильно редуцированы.
Кальмар-хиротеутис (Chiroteuthis veranyi Chun, 1900). Проостракум гладиуса состоит из единственного элемента – рахиса. Конус узкий и длинный, игловидный, занимает примерно 50% длины гладиуса. Рострум отсутствует. В местах прикрепления мантии к гладиусу хрящевая ткань хорошо развита. Как и у мастиготеутид, мантия у хиротеутид не распространяется назад далее переднего края конуса, а сам конус окружен студенистым валиком вакуолизированной аммонийной ткани. Ретракторы воронки и головы имеют раздельные зоны прикрепления к гладиусу. Висцеральный мешок с вентральной стороны от печени образует выпячивание, в котором помещаются прямая кишка, нижняя полая вена, чернильный мешок и фотофоры. Плавники прикрепляются к гладиусу жестко, базальные эпителиальные мешки отсутствуют.
Кальмар - галитеутис (Galiteuthis Joubin, 1898). Гладиус представляет собой узкую и тонкую игловидную пластинку, слабо расширенную в средней части. Проостракум узкий, состоит из медиальной пластинки (рахиса) и пары латеральных пластинок. Конус и рострум отсутствуют. Вместо конуса развивается псевдоконус, образованный свернутыми в трубку половинками гладиуса. Гладиус сложен двумя раковинными слоями: средним (хитиновым) и внутренним (хрящеподобным). Передняя часть проостракума тонкая, пленковидная, лишена опорной роли вследствие срастания переднего края мантии с головой и воронкой. Хрящевая ткань в местах прикрепления мантии к гладиусу выражена только в задней половине мантии. Ретракторы головы полностью редуцированы. Стенки воронки и воротниковые складки превратились в тонкие слабо мускулистые перегородки-септы. Ретракторы воронки на всем протяжении срастаются внешними краями со стенками мантии и превращаются в тонкие косые септы, идущие косо вверх и назад по обе стороны от висцерального мешка. Задние окончания ретракторов прикрепляются внутренними краями к расширенной части оторочки гладиуса, а внешними краями – к стенкам мантии. Плавники не сращены основаниями, крепятся к гладиусу жестко, без базальных эпителиальных мешков.
V. СПОСОБЫ КОНТАКТОВ МЫШЦ С РАКОВИНОЙ:
ПЕРВИЧНЫЕ И ВТОРИЧНЫЕ КОНТАКТЫ
Внутрираковинные головоногие подкласса Coleoidea являются единственной группой в типе Mollusca, в которой мышцы крепятся не только к внутренней, но и к внешней поверхности раковины. Решение этой сложной задачи стало возможным благодаря двум взаимозависимым эволюционным приобретениям: появлению переднего дорсального выроста стенки жилой камеры (проостракума) и обрастанию этого выроста мантией с образованием вокруг него эпителиального раковинного мешка, замкнутого на дорсальной (внешней) стороне. Появление эпителиального мешка, плотно облегающего раковину, сделало возможным прикрепление к нему мышц, причем с функциональной точки зрения этот способ оказался равноценным прикреплению мышц непосредственно к самой раковине.
Таким образом, с эволюционной точки зрения разнообразие способов взаимодействия мышц с раковиной в подклассе Coleoidea, можно разделить на две группы: первичные и вторичные контакты. Первичные контакты унаследованы Coleoidea от наружнораковинных предков. Они образованы мышцами-ректракторами головы и воронки и в типичном случае представляют собой непосредственное прикрепление мышц к внутренней поверхности раковины. Вторичные контакты появились в результате обрастания мантией раковины в процессе погружения последней внутрь тела. Они образованы мантией и ее производными (плавниками) и характеризуются опосредованным взаимодействием мышц и раковины через раковинный мешок. В местах вторичного контакта в стенках раковинного мешка всегда образуется более или менее выраженный хрящ. В наибольшей степени зоны первичного контакта сохранились у представителей семейств: Sepiidae, Vampyroteuthidae, Loliginidae, Thysanoteuthidae, а также у некоторых микронектонных кальмаров с широким гладиусом - семейств Cthenopterygidae, Enoploteuthidae и др. Зоны вторичного контакта наиболее развиты у нектонных кальмаров семейств Ommastrephidae, Onychoteuthidae, Gonatidae, у сепиолид (Rossiinae), а из форм, имеющих фрагмокон, – у Spirulida.
Характер прикрепления мышц к раковине в зонах первичного контакта весьма консервативен. Как правило, это непосредственное прикрепление мышц к внутренней поверхности раковины. Напротив, в зонах вторичного контакта способы прикрепления мышц к раковине могут быть самыми разнообразными. В общем виде их можно разделить на три основные группы: неподвижное прикрепелние путем прирастания мышц к раковинному мешку; подвижное прикрепление скользящего типа (плавники); подвижное прикрепление замкового типа (затылочный замыкательный хрящ).
1) Неподвижные соединения путем прирастания мышц к раковинному мешку. Исходным является, по-видимому, простое прикрепление мышц к раковинному мешку (рис. 1А). В более сложных случаях в месте контакта развивается хрящевая ткань, а на поверхности раковины появляются структуры, повышающие прочность контакта. Это могут быть: бугорки на дорсальной поверхности раковины (сепион каракатиц сем. Sepiidae; рис. 1B); продольные ребрышки и бороздки (внешняя поверхность конуса Ommastrephidae; рис. 1C); ребра жесткости различного профиля (Lycoteuthis; рис. 1D); глубокие щели вдоль утолщенных краев раковины (Moroteuthis; рис. 1E); продольные пазы различного профиля и глубины, образованные складками проостакума (стебель Ommastrephidae; рис. 1F); крыловидные выросты наружного раковинного слоя в задней части гладиуса (сем. Gonatidae; рис. 1G); спиральное эндогастрическое закручивание заднего отдела гладиуса («улитка» Histioteuthis; рис. 1H); спиральное продольное закручивание краев гладиуса (Gonatidae; рис. 1I).
2) Подвижные прикрепления скользящего типа (плавники). При этом виде вторичных контактов в основании плавников, в месте их контакта с раковиной, развиваются базальные хрящи с широким плоским основанием. В типичном случае между основаниями плавниковых хрящей и раковинным мешком имеются замкнутые эпителиальные базальные мешки, стенки которых образуют поверхности скольжения и обеспечивают вертикальную подвижность плавников при маневрировании (рис. 2А). Вертикальные перемещения оснований плавников регулируются специальными мускулами: элеваторами и депрессорами. В онтогенетическом аспекте базальные эпителиальные мешки являются производными раковинного мешка [Naef, 1928].
3) Подвижное прикрепление замкового типа (затылочный хрящ). Затылочный замыкательный хрящ – это единственная зона вторичного контакта, расположенная на внутренней стороне раковины, в области переднего свободного отдела рахиса. Он образован двумя контактными хрящами, комплементарными друг другу: мантийным и затылочным (рис. 2В). Крайний консерватизм в строении затылочного хряща свидетельствует о появлении этой структуры на самых ранних этапах эволюции Coleoidea и жестких функциональных ограничениях на её изменения.

Рис. 1. Неподвижные вторичные контакты мышц с раковиной у Coleoidea. Условные обозначения: mn – мантия; sh – раковина; dsh – дорсальный щит (у каракатиц); radr – радиальные ребра альвеолы; mrib – щель в маргинальном ребре гладиуса; сc – хрящевой чехол; wpl - крыловидные выросты альвеолы; helix – спиральное закручивание заднего отдела гладиуса; coil – спиральное закручивание краёв гладиуса.

Рис. 2. Подвижные вторичные контакты у Coleoidea. А. Подвижное прикрепление скользящего типа (прикрепление плавников к дорсальной поверхности раковинного мешка у каракатицы Sepia). В. Подвижное прикрепление замкового типа (затылочный хрящ у кальмара Doryteuthis). Условные обозначения: sh – раковина ; rm – хрящевая стенка раковинного мешка; bp – базальный эпителиальный мешок; bcf – базальный хрящ плавника; ncv – висцеральный компонент затылочного хряща; ncm – мантийный компонент затылочного хряща.
VI. РАЗНООБРАЗИЕ ПЛАВНИКОВ СОВРЕМЕННЫХ КОЛЕОИДЕЙ
И ИХ ПРОИСХОЖДЕНИЕ
Наличие мускулистых плавников является одним из диагностических признаков подкласса Coleoidea. Среди современных колеоидей плавники отсутствуют лишь у инцирратных осьминогов (отр. Incirrata), однако у древних представителей этой группы (Palaeoctopus) они имелись. При всем разнообразии плавников колеоидей, все они имеют общий план строения и выполняют локомоторную функцию, участвуя в движении и в активном поддержании ориентации тела в пространстве. Плавники являются наиболее распространенным видов вторичных контактов у колеоидей, и их исследование позволяет глубже понять основные закономерности трансформации раковины из наружной во внутреннюю.
Классификация плавников. Предложена новая классификация плавников колеоидей, основанная на следующих морфо-функциональных критериях: относительная длина плавника (соотношение длины основания плавника к его ширине); форма; размеры относительно мантии; положение плавника (субтерминальное или терминальное); характер прикрепления к телу (подвижное/неподвижное); крепление оснований плавников (раздельное или совместное); локализация крепления (к раковине, либо к мантии). На основании этих критериев плавники колеоидей классифицированы на следующие группы: весловидные, языковидные, почковидные, ромбические, сердцевидные, округлые, эллиптические и плавниковая кайма (рис. 3). Весловидные плавники (рис. 3А) встречаются у вампиров и цирратных осьминогов. Основания весловидных плавников всегда раздельные. Такие плавники работают, подобно крыльям птиц, а плавание вампиров и цирратных осьминогов напоминает полет. У всех головоногих с весловидными плавниками они являются основными органами локомоции (способность к реактивному движению ослаблена, либо вовсе утрачена). Почковидные плавники (рис. 3В) встречаются у многих современных колеоидей: спирул (отр. Spirulida), сепиолид (отр. Sepiolida), кальмаров семейств Pickfordiateuthidae, Australiteuthidae, Bathyteuthidae, Pyroteuthidae, Alluroteuthidae, Histioteuthidae и некоторые Cranchiidae. Основания почковидных плавников всегда раздельные. Такие плавники характерны для мелких планктонных и микронектонных головоногих, «порхающих» в толще воды. При этом само животное часто висит головой вниз (Spirula, Bathyteuthidae, Cranchiidae). Колеоидеи с почковидными плавниками как правило сочетают плавниковое плавание с реактивным. Ромбические и сердцевидные плавники (рис. 3С,D) наиболее распространены среди нектонных, микронектонных и некто-бентосных кальмаров сем. Loliginidae, Lycoteuthidae, Enoploteuthidae, Gonatidae, Ommastrephidae, Onychoteuthidae и др. В пределах этого типа выделяются две большие группы: плавники с раздельными основаниями и плавники со сросшимися основаниями. Раздельные основания характерны для плавников шельфовых кальмаров сем. Loliginidae (рис. 3С); плавники со сросшимися основаниями встречаются у многих семейств океанических кальмаров (рис. 3D). Прослеживается параллелизм форм между плавниками с раздельными и сросшимися основаниями, что обусловлено, по-видимому, сходством экологии видов. У быстро плавающих нектонных кальмаров (Loligo, Ommastrephidae, Gonatidae) плавники имеют меньшие относительные размеры, чем плавники микронектонных и нектобентосных форм (Loliolus, Lolliguncula, Enoploteuthidae, Onychoteuthidae). Все кальмары с ромбическими плавниками сочетают плавниковое плавание с реактивным.
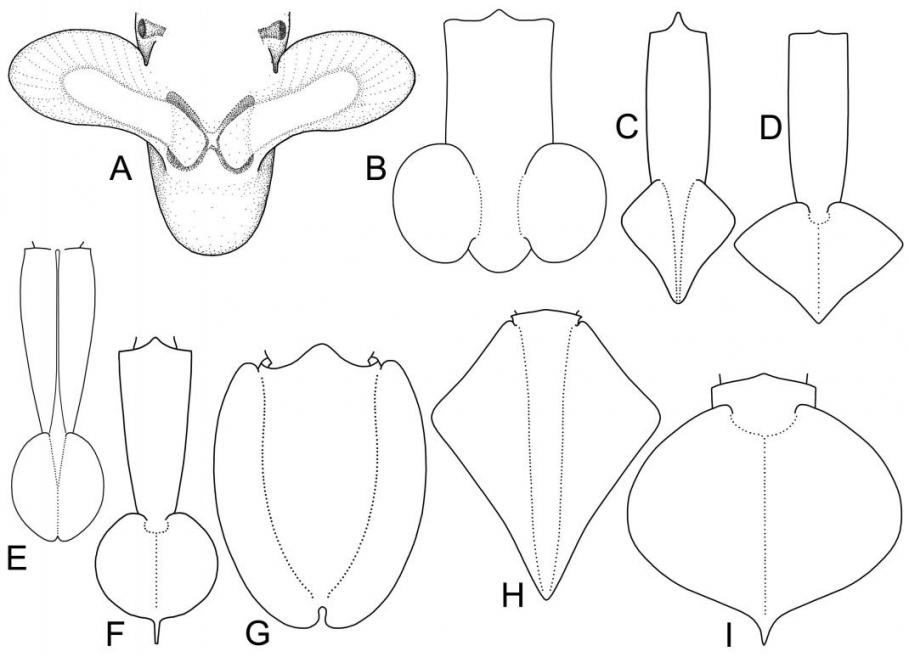
Рис. 3. Морфологическое разнообразие плавников современных Coleoidea. А. Весловидные плавники (Cirroteuthis). В. Почковидные плавники (Pickfordiateuthis). С. Ромбические плавники с раздельными основаниями (Loligo). D. Ромбические плавники со сросшимися основаниями (Todarodes). E. Округлые плавники с раздельными основаниями (Liocranchia). F. Округлые плавники со сросшимися основаниями (Chiroteuthis). G. Плавниковая кайма (Sepia). Н. Большекрылые плавники с раздельными основаниями (Thysanoteuthis). I. Большекрылые плавники со сросшимися основаниями (Octopoteuthis).
Округлые или эллиптические плавники (рис. 3E,F) свойственны малоподвижным полупланктонным и планктонным кальмарам. Основания таких плавников могут быть как раздельными (сем. Cranchiidae; рис. 3E), так и сросшимися (сем. Chiroteuthidae, Joubiniteuthidae, Grimalditeuthidae и др.; рис. 3F). Опорой таким плавникам служит удлиненный задний отдел гладиуса, который иногда выступает позади плавников в виде длинного и тонкого хвоста. Плавники этого типа – наименее мускулистые. Мышечный слой в них столь тонок, что у многих видов в прижизненном состоянии плавники прозрачны. Это обусловлено тем, что плавники данного типа используются не для активного движения, а для поддержания положения тела в пространстве. Многие кальмары с округлыми или эллиптическими плавниками утеряли способность к реактивному плаванию. Плавниковая кайма (рис. 3G) - плавники в виде узкой каймы, протягивающейся по бокам мантии, встречаются у каракатиц-сепиид и кальмаров сем. Loliginidae, Chtenopterigidae и Neoteuthidae. Основания плавников всегда раздельные. Животные с такими плавниками отличаются наименьшей скоростью и наибольшей маневренностью. Колеоидеи с плавниками в виде каймы ведут придонный образ жизни на литорали, шельфе и склоне, в биотопах со сложной пространственной структурой. Как правило они имеют хорошо развитый мантийно-вороночный комплекс и сочетают медленное плавниковое плавание с быстрым гидрореактивным. Большекрылые плавники имеют разнообразную форму (сердцевидную, ромбическую, округлую) и занимают свыше 90% длины мантии или покрывают её целиком (рис. 3Н,I). Плавники такого вида встречаются у пелагических кальмаров сем. Thysanoteuthidae, Octopoteuthidae, Ancistrocheiridae, Cycloteuthidae, Magnapinnidae и Mastigoteuthidae. Большекрылые плавники являются основным органом движения, а при редукции реактивного аппарата (сем. Magnapinnidae) – единственным движителем [Vecchione et al., 2001]. У кальмара-ромба плавники имеют раздельные основания (рис. 3Н), а в остальных семействах большекрылых кальмаров основания плавников срастаются и неподвижно прикрепляются к гладиусу с дорсальной стороны (рис. 3I).
Наиболее примитивное состояние из описанных выше типов имеют субтерминальные плавники почковидной формы, поскольку именно такие плавники закладываются в онтогенезе большинства современных колеоидей. Сравнительный анализ морфологии плавников наглядно иллюстрирует распространенность параллелизмов в их эволюции у современных колеоидей. Виды головоногих со сходной экологией как правило имеют плавники сходной формы, однако далеко не всегда это дополняется сходством внутреннего строения. Можно сделать вывод, что форма и функциональные характеристики плавников обусловлены экологией вида, тогда как детали их внутреннего строения дают достаточно сравнительно-анатомического материала для филогенетического анализа на уровне родов, семейств и отрядов.
Происхождение плавников. Проведенный анализ морфологии плавников современных колеоидей позволил конкретизировать представления о происхождении и эволюции плавников головоногих моллюсков, разработанные Нэфом в начале ХХ века [Naef, 1921/23]. Согласно Нэфу, плавники первоначально возникли как продольные складки мантии, обволакивавшей раковину, и располагались вдоль дорсо-латеральных стенок конотеки (рис. 4А). Как показано в предыдущей главе, плавники являются зонами вторичного контакта мышц с раковиной, и для таких контактов характерно развитие хряща. Это даёт основание предположить, что на этапе формирования плавников в области контакта плавниковой складки с поверхностью раковины развился хрящ, ставший в последствии базальным хрящом плавников.

Первоначально этот хрящ мог быть частью раковинного мешка. Непосредственный контакт базальных хрящей с поверхностью раковины обеспечивал плавникам прочную опору, но затруднял регулировку их положения при маневрировании. При смещении плавника его базальный хрящ скользил по поверхности раковины, вызывая растягивание раковинного мешка и появление на нем складок. Следующим этапом формирования плавников было появление устойчивой складки раковинного эпителия, располагавшейся между раковиной и базальными хрящами плавников (рис. 4B).
Рис. 4. Происхождение плавников в подклассе Coleoidea (по Naef, 1921/1923, с изменениями).
А. Образование мантийной складки на дорсо-латеральных сторонах раковины.
В. Появление складки под основаниями формирующихся плавников.
С. Обособление базальных мешков от раковинного мешка.
D. Смещение оснований плавников с раковины на внешнюю поверхность мантии.
Условные обозначения: sh – раковина ; bcf – базальный хрящ плавника; mn – мантия; bp – базальный эпителиальный мешок; pore – канал, между раковинным мешком и базальными эпителиальными мешками. em – мускул-элеватор; dm – мускул-депрессор. find – дорсальный мускульный слой плавника; finv – вентральный мускульный слой плавников.
Складка отделила от раковинного мешка боковые карманы, что позволило разделить эпителий, секретировавший раковину, от эпителия, обеспечивавшего скольжение плавников. По-видимому, одновременно с боковыми эпителиальными карманами из прилегающих участков мантии дифференцировались мускулы-регуляторы (элеваторы и депрессоры), настраивавшие положение оснований плавников. На следующем этапе прикрепление плавников приобрело современный вид: боковые эпителиальные карманы изолировались от раковинного мешка и превратились в замкнутые базальные мешки, обеспечивавшие скольжение оснований плавников при маневрировании (рис. 4С). Наконец, у некоторых видов кальмаров мантия распространилась на дорсальную поверхность гладиуса и полностью поглотила его, вследствие чего крепление плавников утеряло контакт с гладиусом, сместившись на поверхность мантии (рис. 4D). Единообразный для всех Coleoidea план строения плавников и их сочленения с телом указывает на то, что плавники появились на самой заре становления этой группы, до дифференциации всех филогенетических стволов.
VII. ГОМОЛОГИИ СТРУКТУРНЫХ ЭЛЕМЕНТОВ РАКОВИНЫ В ПОДКЛАССЕ COLEOIDEA
Морфология раковин современных представителей подкласса Coleoidea столь разнообразна, что гомологическое сопоставление их структурных элементов у основных групп – каракатиц, кальмаров и осьминогов – до сих пор сталкивалась с непреодолимыми трудностями [Donovan, Toll, 1988; Хромов, 1990]. Эти трудности, однако, можно преодолеть, если мы будем рассматривать раковину не изолированно, а в окружении прикрепленной к ней мускулатуры и мягкого тела.
Сравнительно-морфологический анализ современных головоногих моллюсков показывает, что морфология их мягкого тела, прежде всего мускульной системы, гораздо более консервативна, чем морфология их раковины. В самом деле, в отличие от раковины, мышечная система головоногих имеет общий для всех Coleoidea план строения, подробно описанный А. Нэфом [Naef, 1921/1923]. Этот план строения существенно не меняется у форм с сильно редуцированной раковиной (Octopodidae; Sepiolidae) и даже при полном исчезновении раковины (Argonautoidea; Idiosepiidae). Образно говоря, эволюция раковины Coleoidea проходила в постоянной системе координат, задаваемых мышечной системой этих моллюсков. Поскольку все мышцы локомоторной системы прямо или косвенно контактируют с раковиной, гомологизацию структурных элементов последней можно проводить по расположению и зонам прикрепления соответствующих мышц: мантии, плавников, ретракторов головы и воронки. Из немышечных элементов для анализа гомологий большое значение имеет положение звездчатых ганглиев.
В большинстве эволюционных ветвей Coleoidea гидростатическая и защитная функции раковины снижалась, а опорная, наоборот, возрастала [Зуев, 1965]. При редукции фрагмокона и превращении раковины в декальцинированный гладиус опорная функция становится основной и начинает определяеть план строения раковины. Происходит это следующим образом. Каждая группа мышц имеет собственное специфическое место (зону) прикрепления к раковине. По мере роста животного размеры мышц увеличиваются, и зоны их прикрепления постепенно расширяются и смещаются, вследствие чего на раковине образуются расширяющиеся сектора. Именно этим – смещением зон мышечного крепления в процессе роста – обусловлен радиальный план строения раковин всех колеоидей. Все структурные элементы раковины колеоидей – рахис, латеральные пластинки, крылья, флаги конуса – всегда имеют вид секторов, расходящихся от заднего апикального конца раковины.
Таким образом, в настоящей работе для анализа гомологий раковин в подклассе Coleoidea применялись следующие морфологические критерии:
- Положение затылочного хряща. В тех случаях, когда он имеется, этот хрящ всегда приурочен к переднему свободному отделу медиальной пластинки (рахиса) проостракума и соответствует ему по ширине, а часто и по длине.
- Положение зон прикрепления медиальных сегментов ретракторов головы. В тех случаях, когда медиальные сегменты ретракторов головы не объединены с латеральными, зоны их прикрепления располагаются в области медиальной пластинки (рахиса) позади затылочного хряща.
- Положение зон прикрепления латеральных сегментов ретракторов головы. В тех случаях, когда латеральные сегменты ретракторов головы не слиты с медиальными, зоны их прикрепления располагаются вдоль передних краев латеральных пластинок, по бокам от рахиса, на одном уровне или несколько позади прикрепления медиальных сегментов.
- Положение зон прикрепления ретракторов воронки. Ретракторы воронки прикрепляются к передним краям латеральных пластинок проостракума, позади от мест прикрепления латеральных сегментов ретракторов головы. У некоторых групп ретракторы головы и латеральные сегменты ретракторов воронки срастаются в задней части; в этом случае зоны их прикрепления на раковине не разграничены. В группах, где ретракторы воронки не срастаются с ретракторами головы, зоны их прикрепления как правило морфологически дифференцированы в виде отдельной пары пластинок – крыльев – расположенных с внешней стороны от латеральных пластинок.
- Положение звездчатых ганглиев. У подавляющего большинства Coleoidea они располагаются у краев раковины и приурочены к уровню появления латеральных пластинок проостракума.
- Крепление мантии. Крепление мантии к проостракуму весьма разнообразно: мантия может крепиться по краям проостракума, либо к его дорсальной части. Для гомологического анализа важно крепление вентральной стенки мантии в задней части раковины: здесь мантия всегда прирастает вдоль переднего края вентральной стенки конуса и вдоль краев флагов конуса, с внутренней либо наружной стороны.
- Положение оснований плавников. В тех случаях, когда плавники не срастаются основаниями, их положение, как уже отмечалось выше, приурочено к флагам конуса.
Применение этих критериев впервые позволило провести гомологическое сопоставление структурных элементов раковин всех групп рецентных Cephalopoda. Так, сопоставление мускулатуры Nautilus и представителей подкласса Coleoidea показало, что ретракторы воронки наутилуса гомологичны таковым колеоидей, ретракторы головы наутилуса гомологичны латеральным сегментам ретракторов головы колеоидей, а затылочные ретракторы наутилуса, по-видимому, гомологичны медиальным сегментам ретракторов головы колеоидей. Зоны прикрепления этих мышц – вся дорсальная, дорсолатеральные и латеральные стенки жилой камеры наутилуса – топологически соответствуют медиальной и латеральным пластинкам проостракума колеоидей, что свидетельствует о кардинальном сужении раковины при её переходе из наружного во внутреннее состояние.
Анализ раковины Spirula выявил, с одной стороны, примитивное архаичное строение раковины, а с другой – далеко не примитивный характер прикрепления к раковине мышц. Все мышцы локомоторного аппарата спирулы прикрепляются не к внутренней поверхности жилой камеры, как у наутилуса, а либо к внешней поверхности раковины, либо к ее краям, либо вообще крепятся не к раковине, а к стенке мантии. Во всех случаях прикрепления мышц к раковине спирулы представляют собой вторичные контакты, т.е. осуществляется не прямым прирастанием к поверхности раковины, а опосредованно, через хрящевую стенку раковинного мешка. Зоны крепления мышц по мере роста спиральной раковины постоянно смещаются по ее поверхности: раковина как бы проворачивается внутри тела моллюска.
 Рис. 5. Гомологии структурных элементов сепиона каракатиц (A) и гладиуса кальмаров-лолигинид (B). Оттенками серого обозначены зоны прикрепления: ncm – затылочный хрящ; rcm – медиальный сегмент ретракторов головы; rcl – латеральные сегменты ретрактов головы; rf – ретракторы воронки; gst – звёздчатые ганглии; mn – мантия. Пунктиром указано положение асимптот в сепионе. Рис. 5. Гомологии структурных элементов сепиона каракатиц (A) и гладиуса кальмаров-лолигинид (B). Оттенками серого обозначены зоны прикрепления: ncm – затылочный хрящ; rcm – медиальный сегмент ретракторов головы; rcl – латеральные сегменты ретрактов головы; rf – ретракторы воронки; gst – звёздчатые ганглии; mn – мантия. Пунктиром указано положение асимптот в сепионе. |
Гомологическое сравнение сепиона каракатиц и гладиусов кальмаров показало, что дорсальный щит раковины каракатиц гомологичен проостракуму, а по расположению зон мышечного прикрепления он более всего сходен с гладиусами кальмаров-лолигинид (рис. 5). Элементы проостракума каракатиц разделены асимптотическими линиями, образованными изгибами линий нарастания. Медиальная пластинка является зоной прикрепления медиальных сегментов ретракторов головы, которые у Sepia имеют вид двух мускулов (как и у Nautilus), срастающихся в средней части, а также служит опорой мантийного компонента затылочного хряща (рис. 5А). И то, и другое указывает на гомологичность медиальной пластинки проостракума каракатиц медиальной пластинке (рахису) гладиуса кальмаров-лолигинид (рис. 5В). Латеральные пластинки в раковине каракатиц представляют собой зоны прикрепления латеральных сегментов ретракторов головы и ретракторов воронки (рис. 5А). В этом отношении они гомологичны латеральным пластинкам гладиусов лолигинид. Плавники каракатиц не сращены основаниями. С раковиной они контактируют лишь в задней части, проходя над флагами внешнего конуса, самим конусом и достигая апикального конца рострума.
Наиболее ярким примером эффективности новых критериев стало то, что с их помощью удалось определить гомологию раковин осьминогов, в которых из-за сильной редукции никакие узнаваемые структурные элементы уже не прослеживаются (рис. 6). Сделать это помогло сравнение зон прикрепления мантии, плавников и мускулов-ретракторов к гладиусу вампира и к раковинам осьминогов.
 Рис. 6. Прикрепление мышц к раковине Vampyropoda. A. Vampyroteuthis. B. Opisthoteuthis. C. Octopus. Левая колонка: вид с вентральной стороны. Правая колонка: вид сбоку с левой стороны (дорсальная сторона справа. Условные обозначения: mn – мантия; rc – ретракторы головы; rf – ретракторы воронки; Рис. 6. Прикрепление мышц к раковине Vampyropoda. A. Vampyroteuthis. B. Opisthoteuthis. C. Octopus. Левая колонка: вид с вентральной стороны. Правая колонка: вид сбоку с левой стороны (дорсальная сторона справа. Условные обозначения: mn – мантия; rc – ретракторы головы; rf – ретракторы воронки; fin – плавники. |
Так, плавники у колеоидей приурочены к флагам конуса и внешним участкам латеральных пластинок проостракума (рис. 6А). У цирратных осьминогов (рис. 6В) плавники прикрепляются к внешней поверхности расширенных латеральных крыльев, что указывает на гомологию этих участков флагам конуса и прилегающими участками проостракума гладиусов вампиров и кальмаров. Тогда, учитывая общий план строения раковины колеоидей, медиальная часть гладиуса циррат (седло) представляет собой остаток медиальной пластинки (рахиса) проостракума. Далее, у вампиров латеральные сегменты ретракторов головы прикрепляются к латеральным пластинкам, а ретракторы воронки – к крыльям гладиуса. У примитивных цирратных осьминогов вороночные ретракторы прирастают к краям латеральных рогов гладиуса, а ретракторы головы – к периферии крыльев, вдоль их вентральной и дорсальной сторон. Это позволяет говорить о гомологии рогов и внутренней поверхности латеральных крыльев гладиуса циррат латеральным пластинкам и крыльям гладиуса вампиров.
К стилетам бесплавниковых (инцирратных) осьминогов прикрепляется только две группы мышц: мантия и ретракторы воронки (рис. 6С). Мантия обрастает стилеты по всему периметру, но не покрывает их снаружи. Ретракторы воронки прикрепляются к внутренней стороне ближе к переднему концу стилетов. В местах прикрепления стилеты образуют изгиб (угол) с шероховатой поверхностью, обращенный навстречу ретракторам. Прикрепление ретракторов воронки указывает на гомологичность области изгиба стилетов латеральным рогам гладиуса цирратных осьминогов и, соответственно, внешним частям латеральных пластинок проостракума гладиуса вампиров и кальмаров. Заднее плечо стилета, заякоренное в стенке мантии, по-видимому, является остатком латеральных крыльев гладиуса циррат и соответствует флагам конуса. С точки зрения гомологии раковинных слоев, гладиусы цирратных осьминогов и стилеты инциррат вероятно формируются внутренним раковинным слоем, поскольку толщина, консистенция и микруструктура раковины осьминогов весьма сходны с таковыми внутреннего раковинного слоя гладиусов некоторых групп кальмаров.
VIII. МОРФОЛОГИЧЕСКАЯ КЛАССИФИКАЦИЯ РАКОВИНЫ
В ПОДКЛАССЕ COLEOIDEA
В этой главе на основании сравнительно-морфологического анализа раковин современных Coleoidea выделены 12 морфологических типов раковин, различающихся формой, относительными размерами и степенью развития элементов раковины: спирулидный, сепиидный, сепиолидный, лолигинидный, тизанотеутидный, вампиротеутидный, цирратный, инцирратный, хиротеутидный, оммастрефидный, онихотеутидный и кранхиидный. Выделенные типы раковин прослеживаются как на взрослых стадиях, так и на стадиях мальков (рис. 7). Все типы охарактеризованы по единому плану, включающему признаки самой раковины, ее взаимоотношения с мягким телом и характерные признаки мягкого тела. Наиболее характерные черты строения раковины и её взаимоотношения с мягким телом для каждого типа перечислены в таблице 1.
При сравнении выделенных морфологических типов раковины с существующей классификацией колеоидей видно, что группы, выделенные по строению раковины, в целом соответствуют отрядам и подотрядам традиционной системы: Spirulida, Sepiida, Sepiolida, Vampyromorphida, Cirrina, Incirrina. Исключение составляют кальмары: традиционная система делит их на два подотряда – Myopsina и Oegopsina. Все миопсиды имеют гладиус лолигинидного типа, а эгопсиды по строению гладиуса распадаются на шесть различных типов, причем два семейства – Chtenopterygidae и Bathyteuthidae – оказываются в лолигинидном типе вместе с кальмарами-миопсидами.
Сравнение морфологии раковин взрослых стадий и личинок показывает, что у фрагмоконных колеоидей (Spirulida, Sepiida) диагностические признаки раковины проявляются уже на эмбриональных стадиях (рис. 7А,В). В обеих группах фрагмокон закладывается на ранних этапах формирования эмбриональной раковины, и одновременно с его закладкой раковина начинает кальцинироваться. Мальки спирулид и сепиид на стадии выклева имеют раковину, в которой выражено большинство специфических признаков соответствующего типа. В раковинах бесфрагмоконных колеоидей фрагмокон отсутствует уже на стадии выклева и, по-видимому, вообще не закладывается в эмбриогенезе. Все бесфрагмоконные колеоидеи имеют декальцинированную раковину (гладиус).
 |  |
Рис. 7. Морфологические типы раковины в подклассе Coleoidea (слева – раковины мальков; справа – раковины взрослых стадий). А. Спирулидный тип. B. Сепиидный тип. C. Сепиолидный тип. D. Лолигинидный тип. E. Вампиротеутидный тип. F. Цирратный тип. G. Инцирратный тип. H. Тизанотеутидный тип. I. Хиротеутидный тип. J. Оммастрефидный тип. K. Онихотеутидный (ростроморфный) тип. L. Онихотеутидный (безростровый) тип. M. Кранхиидный тип (бесхвостая форма). N. Кранхиидный тип (форма с хвостом).
Гладиусы ряда морфологических типов – цирратного, инцирратного, сепиолидного, тизанотеутидного, оммастрефидного и хиротеутидного – имеют специфические черты строения уже на стадии выклева. Гладиусы мальков этих групп выглядят как маленькое подобие гладиусов взрослых особей и отличаются от них лишь меньшей дифференциацией структурных отделов и слабым развитием опорных элементов. У цирратных осьминогов раковина закладывается в виде небольшой поперечной пластинки (рис. 7F), а у инцирратных осьминогов раковинный мешок в эмбриогенезе делится надвое, формируя палочковидные стилеты [Naef, 1928]. Гладиус мальков сепиолид значительно короче мантии, а рахис не достигает заднего конца гладиуса. Гладиус мальков тизанотеутид имеет широкий стреловидный проостракум с выступающими вперед передними краями и мелким чашевидным конусом на заднем конце (рис. 7H). Гладиус личинок оммастрефид имеет типичную мечевидную форму и состоит из рахиса, укрепленного тремя продольными ребрами жесткости (рис. 7J). Наконец, гладиус личинок хиротеутид на стадии выклева имеет характерную игловидную форму с узким проостракумом и игловидным конусом, занимающим 40-50% длины гладиуса (рис. 7I).
Таблица 1. Характеристика морфологических типов раковин колеоидей: строение раковины
и её взаимоотношение с мягким телом
| Морфологи-ческий тип | Вид рако-вины | Количество раковин-ных слоёв | Рост-рум | Прост-ракум | Число элемен-тов про-остракума | Передний край мантии | Зоны крепления ретракторов головы и воронки |
| Спирулидный | фрагмокон | 3 | нет | нет | --- | свободный | раздельные |
| Сепиидный | сепион | 3 | есть | есть | 3 | свободный | срастаются |
| Сепиолидный | гладиус | 2 (нет наружного) | нет | есть | 3 | свободный | раздельные |
| Лолигинидный | гладиус | 3 | есть | есть | 3 | свободный | срастаются |
| Тизанотеутидный | гладиус | 2 (нет наружного) | нет | есть | 5 | свободный | раздельные |
| Вампиротеутидный | гладиус | 3 | есть | есть | 5 | сращен | раздельные |
| Цирратный | гладиус | 1 (нет среднего и наружного) | нет | нет | --- | сращен | раздельные |
| Инцирратный | стилеты | 1 (нет среднего и наружного) | нет | нет | --- | сращен | раздельные |
| Хиротеутидный | гладиус | 2 (нет наружного) | нет | есть | 3 | свободный | раздельные |
| Оммастрефидный | гладиус | 3 | есть | есть | 3 | свободный | раздельные |
| Онихотеутидный | гладиус | 3 | есть | есть | 5 | свободный | раздельные |
| Кранхиидный | гладиус | 2 (нет наружного) | нет | есть | 3 | сращен | раздельные |
С другой стороны, гладиусы вампиротеутидного, лолигинидного и онихотеутидного типов на стадии выклева мальков имеют строение, сходное между собой (рис. 7D,E,K,L). Их проостракум состоит в основном из рахиса с длинным передним свободным отделом. Боковые элементы проостракума представлены широкой оторочкой, расположенной в задней части гладиуса и недифференцированной на латеральные пластинки и крылья. На заднем конца гладиуса расположен широкий чашевидный конус, на апикальной вершине которого имеется едва заметный колпачковидный рострум. Гладиусы всех групп, имеющих во взрослом состоянии две пары боковых элементов проостракума (латеральные пластинки и крылья), на стадии выклева имеют одну пару боковых элементов, не дифференцированную на латеральные пластинки и крылья. Дифференциация крыльев от латеральных пластинок происходит на более поздних, постличиночных этапах онтогенеза.
Проостракум (в тех случаях, когда он имеется) на ранних онтогенетических стадиях состоит из трех радиальных элементов: медиальной пластинки (рахиса) и пары латеральных пластинок. У сепиид рахис почти не выступает за передние края латеральных пластинок (передний свободный отдел рахиса отсутствует). У сепиолидного, вампиротеутидного, лолигинидного, тизанотеутидного, онихотеутидного и хиротеутидного типов рахис заметно выступает вперед за края латеральных пластинок, а у оммастрефидного и кранхиидного типов рахис является основной частью гладиуса.
Наличие единого исходного плана строения проостракума всех современных колеоидей свидетельствует о монофилии Coleoidea. С другой стороны, различия в строении раковин, проявляющиеся в разных группах на разных этапах онтогенеза, указывают на разное время формирования отдельных групп в ходе эволюции колеоидей. Глубокие различия гладиусов отдельных групп эгопсид – онихотеутидного, оммастрефидного, тизанотеутидного, хиротеутидного типов – свидетельствуют о полифилии подотряда Oegopsina в его нынешнем составе.
IX. РАКОВИНЫ ИСКОПАЕМЫХ COLEOIDEA
Данная глава содержит обзор строения раковин основных групп ископаемых колеоидей преимущественно по литературным данным. На основании зависимостей, выявленных между раковиной и мягким телом у рецентных колеоидей, сделаны реконструкции внешнего облика и образа жизни основных ископаемых групп.
X. ИЗМЕНЕНИЯ ФОРМЫ И ФУНКЦИИ РАКОВИНЫ И ЛОКОМОТОРНОГО АППАРАТА ГОЛОВОНОГИХ МОЛЛЮСКОВ ПРИ ПОГРУЖЕНИИ
РАКОВИНЫ ВНУТРЬ ТЕЛА
Обзор палеонтологических данных (глава IX) показывает, что первые колеоидеи (Aulacocerida) не имели проостракума и, вероятно, еще не имели и мускульной мантии. Скорее всего, у древнейших колеоидей первичная немускулистая мантия просто обволакивала раковину снаружи. Однако уже это давало внутрираковинным головоногим преимущество перед наружнораковинными: мантия могла образовывать складки, которые стали органами движения нового типа – появились первые плавники. В эволюции колеоидей появление плавников, вероятно, предшествовало формированию мантийно-вороночного реактивного аппарата. Кроме того, обволакивание раковины мантией сделало возможным горизонтальную балансировку тела за счет развития рострума.
Для использования мантии в качестве органа движения необходимо появление выступа на краю жилой камеры – проостракума, – который мог бы служить для мантии опорой. Появление проостракума стало вторым, после раковинного мешка, эволюционным приобретением Coleoidea, определившим весь комплекс характерных признаков этой группы. С появлением проостракума первичная немускулистая мантия по обе стороны от него трансформировалась во вторичную (мускулистую) мантию колеоидного типа. Тем самым завершилась дифференциация первичной мантии на плавники и мускулистую мантию. Поскольку мантийная полость после появления проостракума уже не ограничивалась жилой камерой, появилась возможность существенно увеличить мощность реактивного двигателя за счет увеличения объема мантийной полости и наращивания мантийной мускулатуры. Поддержка нового органа движения – мускульной мантии – была не единственной функцией проостракума. Одновременно с ней он обеспечивал опору мускулов-ретракторов, висцерального мешка, удлиняющегося вследствие формирования мускульной мантии, и фиксацию положения головы и воронки при реактивном плавании. До формирования раковинного мешка, замкнутого на дорсальной стороне проостракума, эффективное прикрепление мантии и плавников к раковине и, следовательно, их использование в качестве органов локомоции, с функциональной точки зрения невозможно.
Дифференциация структурных элементов проостракума Coleoidea происходила, вероятно, одновременно с формированием самого проостракума. Парные латеральные элементы проостракума формировались из боковых стенок жилой камеры как зоны первичного контакта ретракторов головы и воронки. Медиальная пластинка (рахис), вероятно, формировалась из первоначального дорсального выступа жилой камеры, обеспечивавшего опору мантии и стабилизацию головы. Наличие у большинства современных колеоидей хрящевого футляра (затылочного хряща) вокруг переднего отдела медиальной пластинки указывает на то, что эта часть раковины образует зону вторичного контакта с мышцами и, следовательно, является новообразованием. В то же время позади затылочного хряща на медиальной пластинке находится зона прикрепления ретракторов головы, представляющая собой зону первичного контакта. Можно предположить, что ретракторы головы у древнейших колеоидей первоначально прикреплялись к внутренней стенке жилой камеры, но затем, по мере формирования проостракума и удлинения тела, сместились вперед на проостракум, сохранив характер прикрепления к раковине.
Трансформация раковины Cephalopoda из наружной во внутреннюю сопровождалась сменой ее основных функций. У Nautilus главными функциями раковины являются: защитная, гидростатическая, опорная и структурная (раковина определяет объем и форму жилой камеры и всего мягкого тела моллюска). У Coleoidea раковина превращается во внутренний скелет, дающий опору мантии, воронке и плавникам. Опорная функция раковины становится главной, гидростатическая сохраняется лишь у немногих представителей, структурная функция меняет свое содержание (раковина становится основной несущей конструкцией), а защитная утрачивается полностью.
Превращение раковины из наружной во внутреннюю сопровождалось радикальной сменой способов движения и дыхания (рис. 10). У Nautilus имеется только один способ движения – гидрореактивный – с двумя режимами: медленного и быстрого плавания. В режиме медленного плавания реактивная тяга возникает как побочный эффект респираторного цикла, за счет дыхательных движений воротниковых складок. В режиме быстрого плавания реактивная тяга создается резким втягиванием головы внутрь жилой камеры в результате сокращения ретракторов головы. Из-за отсутствия фиксации положения жабр в мантийной полости дыхательный цикл в режиме быстрого плавания у Nautilus блокируется (Глава III).

Рис. 10. Различия в локомоторной и респираторной системе Nautilus (А) и кальмаров (В). Стрелками показаны токи воды при дыхании и плавании. А: из [Wells, 1988]; В: из [Naef, 1922].
У Coleoidea режим медленного реактивного плавания за счет дыхательных движений воротниковых складок исчезает, а в режиме быстрого реактивного плавания кардинально меняется механизм создания реактивной тяги: она создается не поршнеподобным втягиванием головы, а сокращением мускульной мантии. Кроме того, появляется принципиально новый более экономичный способ плавания посредством плавников. Важным приобретением Coleoidea стала фиксация жабр на внутренней стенке мантии с помощью специального лигамента, что позволило не прерывать дыхательный цикл в режиме реактивного плавания и сделать этот способ движения постоянным.
Появление новых способов движения у внутрираковинных головоногих привело к глубокой перестройке их мышечной системы. У Nautilus основными локомоторными мышцами являются ретракторы головы, прикрепляющиеся к дорсолатеральным стенкам жилой камеры (рис. 10). Ретракторы воронки развиты слабо. У Coleoidea ретракторы головы утрачивают локомоторную функцию и сильно уменьшаются. У большинства современных форм они редуцированы до тонкой мускульной оболочки, окружающей висцеральный мешок. Ретракторы воронки, напротив, усиливаются, обеспечивая удержание и управление воронкой в условиях возросших нагрузок при быстром реактивном плавании. Главными локомоторными мышцами Coleoidea становятся мускульная мантия и плавники.
До тех пор, пока раковина Coleoidea сохраняла фрагмокон, она оставалась обызвествленной. Напротив, исчезновение газового фрагмокона неизбежно вело к полной декальцинации раковины. В самом деле, газово-жидкостный гидростатический аппарат головоногих может функционировать лишь при наличии жестких и прочных стенок раковины, фиксирующих его объем и способных выдерживать колебания внешнего давления. Следовательно, обызвествление раковины головоногих является необходимым условием выполнения ею гидростатической функции; оно всегда связано с наличием фрагмокона. Напротив, внутренний скелет, если он не сочлененный, как у хордовых, должен быть гибким, эластичным и по возможности легким. Отсутствие обызвествления (декальцинация) раковины свидетельствует о ее превращении в исключительно опорную структуру, что предполагает утрату фрагмокона. По-видимому, такая декальцинация происходила достаточно быстро в эволюционном отношении. Среди современных головоногих нет ни одного примера обызвествленной раковины без фрагмокона. Среди ископаемых форм частично обызвествленными были раковины Paleoteuthomorpha и Teudopseinae. В первом случае это связано с частичной редукцией фрагмокона, а во втором – с относительно недавней в эволюционном отношении потерей фрагмокона предками теудопсид. Ярким примером функционально обусловленного обызвествления раковины являются белемниты: их гидростатический аппарат (фрагмокон и его противовес-рострум) обызвествлены, а опорный аппарат – проостракум – декальцинирован [Jeletzky, 1966].
XI. ЭВОЛЮЦИЯ РАКОВИНЫ В ПОДКЛАССЕ COLEOIDEA
Анализ строения раковин древнейших Coleoidea дает основания предполагать, что эта группа произошла от наружнораковинных головоногих средних или мелких размеров с прямой узкой раковиной, длинной трубковидной жилой камерой, фрагмоконом с узким вентральным сифоном, небольшим сферическим или овальным протоконхом с цекумом и просифоном. Эти предположения вполне согласуются с палеонтологическими данными: среди древнейших колеоидей нет ни одной спирально закрученной формы. Невозможность происхождения колеоидей от спирально закрученных форм можно подкрепить и логическими доводами. Проостракум – прямая опорная структура, соизмеримая по длине с телом – не мог возникнуть у животных со спирально закрученной раковиной.
Положение ретракторов наутилуса (затылочных, головы и воронки) свидетельствует о гомологии этих мускулов дорсо-вентральным мускулам других классов моллюсков, а наличие у современных головоногих нескольких пар ретракторов указывает на то, что цефалоподы произошли от форм с метамерными дорсо-вентральными мускулами, причем их было не менее трёх пар.
Наиболее вероятными предками Coleoidea являются бактриты (отр. Bactritida Shimansky, 1951). Примитивные представители этого отряда имели прямые узкие раковины [Teichert, 1988], вполне соответствовавшие всем перечисленным критериям. Механизмы балансировки раковины у прямораковинных бактритов отсутствовали, а значит, эти животные висели в воде вниз головой и, очевидно, вели планктонный образ жизни [Holland, 2003]. Гипотеза об исходной для колеоидей планктонной жизненной форме, плавашей вниз головой, на первый взгляд, кажется странной: до сих пор было принято считать, что первичной для колеоидей была прибрежная горизонтально сбалансированная некто-бентосная форма [Несис, 1985; 1990]. Однако эта гипотеза позволяет объяснить некоторые "странные" особенности строения колеоидей. Так, у колеоидей статолиты в органах равновесия (статоцистах) расположены на морфологически передней (вертикальной) стенке камеры, тогда как у остальных моллюсков статолиты/статоконии всегда располагаются на дне статоцистов [Budelmann, 1988]. Трудно понять, как могли появиться статоконии на вертикальной стенке статоциста горизонтально плававших форм, однако все становится очевидным, если представить, что исходный предок висел вниз головой: в этом случае статоконии располагались на передней (нижней) стенке статоцистов – там, где они и должны находиться у моллюсков.
Известные детали строения мягкого тела древних колеоидей позволяют сделать некоторые предположения о возможном строении тела их предков. По-видимому, они имели десять рук примерно одинаковой длины, поскольку именно такое строение рук является исходным для Coleoidea. Широкое распространение у современных колеоидей перепонки-умбреллы между руками, особенно развитой у примитивных и планктонных форм, позволяет предполагать наличие такой перепонки и у их предков. Вооружение рук могло состоять из усиков-цирр и присосок без роговых колец, расположенных в один ряд. Жабры прикреплялись по всей их длине к мантийной стенке специальным лигаментом.
Первые внутрираковинные головоногие, по строению и, вероятно, образу жизни почти не отличались от наружнораковинных предков-бактритов. Однако наблюдаемое среди аулакоцерид разнообразие в развитии рострума, фрагмокона и обызвествления раковины свидетельствует о том, что в пределах этой группы уже происходила дивергенция с формированием различных жизненных форм. От аулакоцерид, по-видимому, происходят, по крайней мере, три крупные группы колеоидей: белемниты, фрагмотеутиды и спирулиды. В линии, ведущей к белемнитам, впервые в эволюции колеоидей произошла практически полная редукция жилой камеры: от неё осталась лишь дорсальная стенка – проостракум. Эти изменения были следствием появления мускульной мантии, ставшей новым органом движения колеоидей. Судя по морфологии раковины, генеральным направлением эволюции белемнитид было формирование горизонтально сбалансированной некто-бентосной прибрежно-неритической формы, сходной с современными кальмарами-лолигинидами.
Эволюция спирулидной ветви. Спирулиды произошли, по-видимому, от аулакоцерид (рис. 11). Об этом свидетельствует наличие в обеих группах слабо обызвествленного рострума с неразвитой радиальной структурой и слабо выраженной апикальной линией, фрагмокона с вентральным сифоном, а также узкого удлиненного тела, соответствующего форме жилой камеры (у аулакоцерид) или проостракума (у древнейших спирулид). Происхождение и ранние этапы эволюции спирулидной ветви не документированы в палеонтологической летописи. Раковины древнейших известных спирулид (Naefia, Adygeya) внешне сходны с раковинами «безростровых» аулакоцерид, однако уже не имеют перламутрового слоя. Жилая камера у них редуцировалась до узкого палочковидного проостракума. На этом этапе у спирулид сохранился прямой фрагмокон и небольшой рострум. По-видимому, сохранился и исходный – планктонный – образ жизни. Крайне узкая раковина меловых спирулид дает основание предполагать, что у этих форм уже произошло смещение основной массы висцеральных органов из редуцированной жилой камеры фрагмокона в мантийную полость, характерное для современных спирулид. В жилой камере осталась только печень.
Следующий этап эволюции спирулид связан с появлением в начале кайнозоя горизонтально сбалансированных раковин с массивным рострумом (Vasseuridae, Belopteridae, Spirulirostridae). Исходными в этом ряду являются Vasseuridae, раковина которых сохраняет прямую форму, лишь слегка изогнутую эндогастрически на ранних онтогенетических этапах. По-видимому, вассеуриды были прибрежными некто-бентосными формами, экологически сходные с современными кальмарами-лолигинидами. Однако, судя по нарастанию рострума, у личинок он отсутствовал, и они, по-видимому, сохраняли предковый планктонный образ жизни с ориентацией головой вниз.
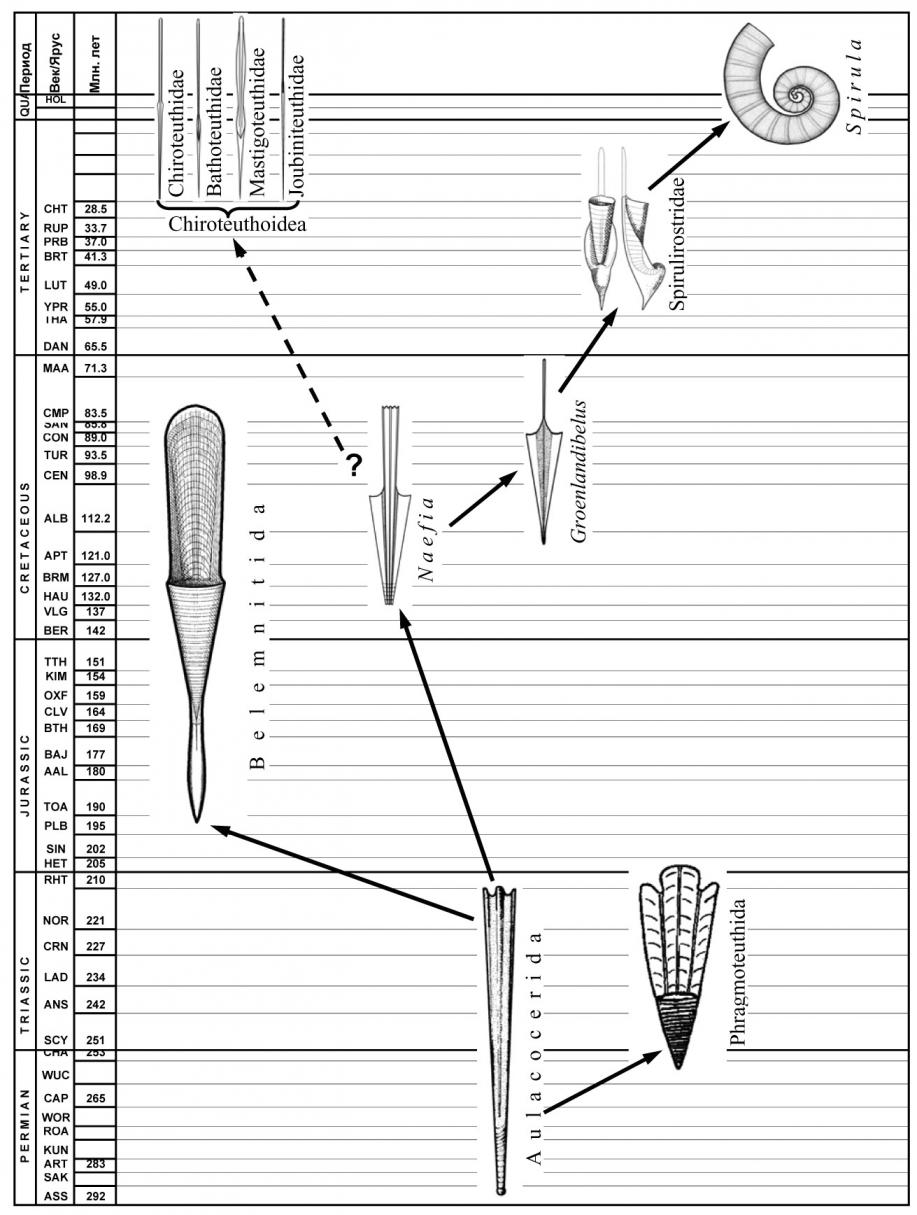
Рис. 11. Ранняя дивергенция эволюционных стволов в подклассе Coleoidea; происхождение и эволюция спирулидного ствола.
Строение раковины Spirulirostridae также указывает на наличие в онтогенезе этой группы планктонной личиночной стадии: на ранних этапах раковина росла эндогастрической спиралью, а у взрослых она становилась прямой. Горизонтальная ориентация взрослых спирулирострид не вызывает сомнений, поскольку она обеспечивалась массивным рострумом и распрямленной раковиной. Мальки спирулирострид и близких им семейств, имея спиральную раковину без рострума и проостракума, могли ориентироваться только вниз головой и вести планктонный образ жизни. Наличие личиночной планктонной стадии у спирулирострид объясняет появление рецентного рода Spirula, который, вероятно, является неотенической формой одной из упомянутых групп. В отличие от предков, спиральный рост у Spirula сохраняется в течение всей жизни, и формирование рострума на взрослой стадии не происходит.
Строение раковин раннемеловых спирулид (Adygeyidae, Groenlandibelidae) поразительно сходно с гладиусами современных мезопелагических «шилохвостых» кальмаров сем. Chiroteuthidae, Mastigoteuthidae, Joubiniteuthidae, Bathoteuthidae. Гладиусы кальмаров этой группы семейств имеют характерную игловидную форму и выделяются в отдельный (хиротеутидный) морфологический тип, отличимый уже на личиночных стадиях. Сходство гладиусов хиротеутидного типа с раковинами ископаемых спирулид дает основание предполагать происхождение «шилохвостых» кальмаров от раннемеловых спирулид, близких к Adygeyidae и Groenlandibelidae. На возможность такого происхождения указывают прямая раковина, узкая палочковидная форма проостракума, узкая игловидная форма конуса/конотеки у представителей обеих групп. Есть еще один признак, сближающий «шилохвостых» кальмаров уже с современной Spirula: наличие в обеих группах характерного смещения переднего отдела висцерального мешка, выступающего внутрь мантийной полости. Возможное время дивергенции «шилохвостых» кальмаров от Adygeyidae Groenlandibelidae – ранний или средний мел. Утрата фрагмокона не только облегчила раковину, но и сняла ограничения на вертикальное распространение. Она открыла для этой группы спирулид новую адаптивную зону – мезо – и бати-пелагиаль открытого океана. Освоение этой зоны при сохранении исходной «парящей» планктонной формы привело к формированию спектра морфологически сходных семейств, составляющих современную группу «шилохвостых» кальмаров.
Сохранение исходной планктонной жизненной формы на протяжении эволюции спирулид, по-видимому, стало основной причиной того, что раковина в этой филогенетической ветви подверглась наименьшим изменениям по сравнению с раковинами других групп колеоидей. Spirula – единственный представитель современных колеоидей, имеющий типичный газовый фрагмокон, а хиротеутоидные кальмары имеют гладиусы, сохранившие черты сходства с раннемеловыми прямораковинными спирулидами.
Отряд Phragmoteuthida и его значение в филогении колеоидей. Фрагмотеутиды занимают ключевое положение в филогении колеоидей, поскольку являются, согласно современным представлениям, наиболее вероятными предками большинства групп современных головоногих моллюсков, включая каракатиц, осьминогов, вампиров и большинство кальмаров. На происхождение фрагмотеутид от аулакоцерид указывает наличие в обеих группах фрагмокона с узким вентральным сифоном и длинной жилой камерой с широким языковидным выступом на дорсальной стороне. Обособление фрагмотеутид от аулакоцерид, очевидно, сопровождалось редукцией вентральной стенки жилой камеры и камерной части фрагмокона, а также развитием длинных сифональных трубок, вложенных одна в другую, вследствие чего стенка сифона у них стала многослойной и пористой [Mutvei and Donovan, 2006]. Редукция вентральной стенки жилой камеры указывала на появление на её месте мускульной мантии и повыение подвижности фрагмотеутид по сравнению с аулакоцеридами. Появление у фрагмотеутид многослойного пористого сифона, повысило механическую прочность фрагмокона и многократно увеличило поверхность водо- и газообмена. Пример современных каракатиц-сепиид показывает, что пористый многослойный сифон позволяет быстро и обратимо менять соотношение газа и жидкости в раковине и, кроме того, делает возможным вторичное заполнение жидкостью наиболее ранних камер для обеспечения горизонтальной балансировки тела. Другой важной чертой, способствовавшей эволюционному успеху фрагмотеутид, было наличие у них широкого проостракума с хорошо дифференцированными продольными элементами – зонами мышечного крепления. Пятичленный проостракум фрагмотеутид оказался удачной основой для формирования мускулатуры широкого спектра экологических форм. Таким образом, с самого начала эволюции фрагмотеутид в этой группе наметилась тенденция ослабления ее гидростатической функции и усиления функции опорной.
Эволюция сепиоидной ветви. До сих пор древнейшими сепиидами принято считать сем. Belosepiidae из эоцена [Young et al., 1998]. Обнаружение признаков, свидетельствующих о принадлежности к сепиидам верхнеюрских Trachiteuthidae (глава IХ) и увеличивает возраст сепиидной ветви более чем на 100 млн. лет. На принадлежность трахитеутид к сепиидной ветви указывают: сходство в строении проостракума и архитектуре раковины; наличие бугристой скульптуры на дорсальной стороне раковины; наличие уплощенного фрагмокона и широкого и мелкого конуса, раскрытого на вентральной стороне; слабое развитие рострума. Перечисленные признаки включают в себя практически все характерные черты строения раковин сепиид, что дает основания не просто причислять трахитеутид к сепиидной ветви, но и считать их наиболее вероятными предками современных каракатиц.
Сем. Trachiteuthidae ведет происхождение, по-видимому, непосредственно от фрагмотеутид (рис. 12). В пользу этого свидетельствует сходство в строении проостракума, фрагмокона (широкий апикальный угол) и слабое развитие рострума у трахитеутид и примитивных фрагмотеутид. Перечисленные признаки образуют устойчивый адаптивный комплекс, последовательно эволюционирующий в ряду Phragmoteuthidae – Trachiteuthidae – Sepiidae. Появление трахитеутид стало закономерным результатом наметившейся у фрагмотеутид тенденции формирования некто-бентосных форм с широкой уплощенной раковиной с трансформированным фрагмоконом и слабо развитым рострумом. Не будет преувеличением сказать, что формирование раковины сепиидного типа произошло в основном на этапе обособления трахитеутид от фрагмотеутид.
Имеющиеся палеонтологические данные свидетельствуют, что в пределах трахитеутид происходила дивергенция, давшая начало двум эволюционным линиям: в одной из них фрагмокон сохранился, а в другой редуцировался. Эволюция первой линии (сохранение фрагмокона) шла по пути освоения придонного биотопа прибрежных, а затем шельфовых и склоновых вод и привела к появлению современных каракатиц-сепиид (рис. 12). Раковина в этой линии менялась незначительно: современные примитивные каракатицы подрода Acanthosepion имеют раковину, поразительно сходную по форме и структуре с раковиной трахитеутид. Не менее успешно эволюционировала и другая линия сепиоидей, сопровождавшаяся редукцией фрагмокона. Эта линия довольно рано (уже в ранней юре) привела к появлению бесфрагмоконных форм с декальцинированной раковиной (Teudopsidae). Раковины примитивных теудопсид по внешней форме столь схожи с трахитеутидами, что многие исследователи объединяли их в один таксон [Jeletzky, 1966; Doyle et al., 1994]. Потеря газового фрагмокона определила дальнейшую эволюцию теудопсид; она открыла им возможность оторваться от дна и начать осваивать толщу воды. Эволюция в этом направлении уже в конце юрского периода привела к появлению форм с гладиусами перовидной формы, которые практически не отличались от гладиусов современных кальмаров-лолигинид: семейству Palaeololiginidae (рис. 12).
По-видимому, не позднее середины палеололигиниды дивергировали на прибрежно-неритическую ветвь, давшую начало современным кальмарам-лолигинидам, и пелагическую ветвь, ведущую к семействам Chtenopterygidae и Bathyteuthidae. Представители прибрежно-неритической ветви эволюционировали в основном по пути формирования нектонных прибрежных и шельфовых форм. Представители пелагической ветви миопсид, по всей видимости, оказались вытеснеными из эпипелагиали более успешными группами океанических колеоидей и рыб в менее конкурентные зоны: в мезо- и батипелагиаль, где они существуют по настоящее время. В процессе освоения глубин пелагические миопсиды приобрели некоторые признаки, конвергентно сходные с другими группами океанических кальмаров: в частности открытую глазную камеру («эгопсидный» глаз).

Рис. 12. Эволюция сепиоидного ствола Coleoidea.
От юрских теудопсид происходит, вероятно, еще одна группа колеоидей: Muensterellidae, у которых произошла редукция боковых полей медиальной пластинки гладиуса, а конус развернулся в плоское лопатовидное раширение (рис. 12). Разворачивание конуса привело к тому, что гладиус мюнстереллид оказался существенно короче мускульной мантии, и его задний апекс сместился в субтерминальное положение. Из современных колеоидей такое взаимоотношение мантии и гладиуса встречается только у сепиолид и осьминогов. Интересно, что в обеих группах прослеживается тенденция редукции раковины вплоть до полного ее исчезновения, сопровождающаяся срастанием переднего дорсального края мантии с затылочной частью головы.
Первые мюнстереллиды (Muensterella scutellaris) были, подобно их предкам-теудопсидам, донными прибрежными колеоидеями средних размеров. Последующая эволюция группы шла по пути развития бенто-пелагических и некто-бентосных форм. Гладиус при этом становился все более узким, лопатовидное расширение на его заднем конце постепенно редуцировалось, а медиальный киль удлинялся и укреплялся. В меловом периоде среди мюнстереллид появились нектонные формы гигантских размеров (Tusoteuthis). Однако все они вымерли в конце мела, а некто-бентосные формы выжили и, видимо, дали начало современному отр. Sepiolida.
У сепиолид гладиус подвергся значительной редукции. Исчезло лопатовидное расширение на его заднем конце: от него сохранилась лишь узкая полоска у некоторых примитивных форм (Rossinae). Боковые элементы проостракума редуцировались до едва заметной узкой оторочки. Эволюция Sepiolida сопровождалась постепенной редукцией гладиуса вплоть до его полного исчезновения. Этот процесс хорошо прослеживается в ряду Rossiinae-Sepiolinae-Heteroteuthinae-Sepiadariidae. Примитивные сепиолиды представлены в основном бентосными или бенто-пелагическим среднеразмерными формами. Ддальнейшая эволюция сепиолид шла по двум основным направлениям: бентонизации и пелагизации [Хромов, 1990]. В обоих направлениях прослеживается постепенное уменьшение размеров тела и редукция гладиуса вплоть до полного исчезновения. Наиболее успешным оказалось первое направление: его современные представители (Rossiinae, Sepiolinae, Sepiadariidae) широко распространены в Мировом океане, а первые две группы – весьма многочисленны. Второе, пелагическое направление эволюции сепиолид оказалось менее успешным. В современной фауне оно представлено единственным подсемейством Heteroteuthinae (сем. Sepiolidae) – мелкими (менее 3 см) пелагическими нерито-океаническими головоногими с короткой мешковидной мантией с большими плавниками. Именно в этой ветви сепиолид сохранились такие архаичные признаки, как глубокая перепонка-умбрелла между руками и широкий языковидный вырост на вентральной стороне мантии, имевшийся еще у раннемеловых мюнстереллид.
Эволюция теутоидной ветви. Древнейшими кальмароподобными колеоидеями являются лолигосепииды (Loligosepiidae) – группа, впервые появившаяся в конце триаса, около 210 млн. лет назад [Jeletzky, 1966]. Судя по строению раковины, это были довольно медлительными донными животными с уплощенным телом, напоминавшим тело современных караткатиц. Строение проостракума лолигосепиид явно указывает на происхождение этой группы от фрагмотеутид (рис. 13). Появление лолигосепиид стало эпохальным событием в эволюции колеоидей: это были первые головоногие, раковина которых полностью утратила фрагмокон и превратилась в декальцинированную пластинку-гладиус. У лолигосепиид гладиус появился уже в конце триаса, т.е. примерно на 20 млн. лет раньше, чем в сепиоидной ветви, и на 100 млн.лет раньше, чем в спирулидной ветви. На независимое происхождение теудопсид и лолигосепиид указывают различия в строении проостракума обеих групп, а также сходство гладиусов лолигосепиид с проостракумом фрагмотеутид, с одной стороны, и гладиусов теудопсид с раковинами трахитеутид, с другой.
Возникает вопрос: почему на рубеже триаса и юры гладиус неоднократно и независимо появлялся в различных филогенетических ветвях колеоидей? Ответ следует искать в изменениях экологического окружения головоногих того времени. В конце триаса между головоногими и рыбами нарастала конкуренция, преимущество в которой получали наиболее быстрые и маневренные виды [Packard, 1972]. Необходимость повышения скорости и маневренности требовала отказа от аппарата пассивного поддержания плавучести: образно говоря, дирижабли уступали место самолетам. Фрагмокон стал помехой для развития мантийного гидрореактивного аппарата и редуцировался. Последовавшая за этим декальцинация превратила раковину в гладиус - легкий и гибкий внутренний скелет, опору развивающегося локомоторного аппарата. Появление гладиуса у лолигосепиид дало гигантские преимущества этим первым кальмарам перед их сородичами с фрагмоконом: аулакотеутидами, белемнитами и фрагмотеутидами. Это преимущество выразилось в том, что уже в начале юрского периода лолигосепииды дали широкий спектр кальмароподобных форм. Одним из первых обособилось сем. Geopeltidae (рис. 13), гладиусы которых отличались от лолигосепиидных удлинением переднего отдела медиальной пластинки. Сужение гладиуса геопелтид обеспечило распространению мантии на латеральные и дорсолатеральные стороны тела и вполне соответствовало тенденции усиления пловцовских качеств этих животных.
Две основные филогенетические линии прослеживаются в пределах Geopeltidae: в одной гладиус становился прогонистым, у него удлинялся передний отдел рахиса и сужался конус (G. symplex); в другой конус становился более широким, а рахис оставался коротким (G. munsteri). Представители "узкоконусной" линии имели, вероятно, узкую прогонистую мантию и эволюционировали в направлении нектонизации. "Ширококонусные" эволюционировали как некто-бентосные или бенто-пелагические формы. Обе линии оказались конкурентноспособными и дали начало группам, потомки которых дожили до наших дней.

Рис. 13. Эволюция теутоидного ствола Coleoidea.
Эволюция "узкоконусной" ветви привела к появлению в ранней юре Plesioteuthidae - кальмароподобных колеоидей с узким мечевидным гладиусом (рис. 13). В пределах плезиотеутид прослеживается постепенное сужение гладиуса и развитие в нем опорных элементов (в ряду Paraplesioteuthis – Boreopeltis – Plesioteuthis – Maioteuthis), указывающее на то, что основным направлением эволюции семейства было формирование активных нектонных хищников. Усиление нектонных тенденций сопровождалось формированием в гладиусах плезиотеутид комплекса признаков, который позволяет уверенно считать плезиотеутид прямыми предками современных океанических кальмаров-оммастрефид. Гладиус наиболее позднего представителя плезиотеутид, Maioteuthis, по внешнему виду уже неотличим от гладиусов современных Ommastrephidae.
Эволюция «ширококонусной», линии геопелтид привела к появлению в средней юре сем. Mastigophoridae (рис. 13) – одной из немногих групп ископаемых колеоидей, чья принадлежность к кальмарам не подвергается сомнению из-за уникальной степени сохранности этих животных [Donovan, 1983]. Строение гладиуса мастигофорид – единая латеральная оторочка, узкий (у продвинутых форм) палочковидный рахис, широкий мелкий конус позволяют предполагать происхождение от этой группы современных кальмаров сем. Cranchiidae через неизвестные пока промежуточные позднемеловые формы. Современные кранхииды – это в основном малоподвижные планктонные кальмары с уникальным набором приспособлений к пассивному парению в океанской пелагиали. Можно предположить, что эта группа вышла в пелагиаль примерно в то же время, что и плезиотеутиды, но изначально эволюционировала в направлении планктонизации. По мере нарастания конкуренции в эпипелагиали кранхииды были вытеснены в мезо- и батипелагиаль, где и сохранились до сих пор, приспособившись к пассивному планктонному существованию.
Проследив эволюционную историю геопелтид и их вероятных потомков вплоть до наших дней, мы снова вернемся в юрский период, чтобы рассмотреть судьбу других ветвей лолигосепиид. В конце юры в океане Тетис появилась многочисленна группа кальмаров, имевших гладиус с широким проостракумом, коротким конусом и рудиментарным рострумом – сем. Leptoteuthidae. Строение проостракума лептотеутид явно указывают на происхождение этой группы от лолигосепиид (рис. 13). Судя по сохранившимся останкам, лептотеутиды были крупными прибрежными некто-бентосными хищниками с мускулистой торпедовидной мантией и весловидными плавниками [Naef, 1922; Jeletzky, 1966]. Эволюция потомков лептотеутид слабо представлена в палеонтологической летописи. Единственной группой, имеющей очевидное родство с лептотеутидами, является сем. Eoteuthoidae из позднего мела (турон) Чехии [Kostak, 2002; 2003]. Строение гладиуса Eoteuthoidae свидетельствуют о филогенетической близости этого семейства к современным кальмарам-эгопсидам, имеющим онихотеутидный тип строения гладиуса. Таким образом, сходство строения гладиусов юрских лептотеутид, меловых Eoteuthoidae и ряда современных эгопсидных кальмаров позволяет отнести их к единой эволюционной ветви, которую для удобства можно называть онихотеутоидной. Основные тенденции изменения гладиуса в этой ветви – сужение медиальной пластинки до узкого утолщенного рахиса, укорочение и сужение парных элементов проостракума, развитие продольных ребер жесткости и медиального киля в задней части гладиуса. Главным направлением эволюции в онихотеутоидной ветви стало освоение эпи- мезо- и батипелагиали открытого океана. Однако представители онихотеутоидной ветви, по-видимому, вышли в пелагиаль позже предков оммастрефид. Вероятно, именно это обстоятельство определило основное направление эволюции онихотеутоидной ветви – освоение мезо- и батипелагиали с формированием широкого спектра среднеглубинного и глубоководного нектона с редкими примерами перехода к планктону [Несис, 1985].
Единственный представитель сем. Thysanoteuthidae, гигантский кальмар-ромб (Thysanoteuthis rhombus) по ряду морфологических, физиологических и экологических признаков стоит особняком среди кальмаров отр. Oegopsida [Nigmatullin, Arkhipkin, 1998]. Строение гладиуса Thysanoteuthis, а также взаимоотношение его с мягким телом, подробно описанное в главе VIII, не имеет аналогов среди современных колеоидей, но поразительно сходно с гладиусами юрских лолигосепиид. Это позволяет считать Thysanoteuthis прямым, хотя и сильно изменившимся, потомком Loligosepiidae (рис. 13). Наличие гладиуса архаичного строения не помешало тизанотеутидной ветви в ходе длительной эволюции освоить океаническую эпи- и мезопелагиаль и сохраниться до наших дней, уйдя от активной конкуренции с кальмарами-оммастрефидами и эпипелагическими рыбами за счет пищевой специализации: кальмары-ромбы питаются днём на больших глубинах (от 400 до 700 м), охотясь в основном на микронектонных рыб, а оммастрефиды питаются ночью у поверхности, в основном никтоэпипелагическими рыбами и кальмарами [ Nigmatullin and Arkhipkin, 1998].
Эволюция вампироподной ветви. Вампироподная ветвь колеоидей происходит, по-видимому, от некто-бентосных представителей сепиидного ствола – теудопсид или их непосредственных предков (рис. 14). Эволюция в этой ветви шла в двух направлениях. Первое, вампиротеутидное, шло по пути специализации к малоподвижному образу жизни на больших глубинах океанской пелагиали. В этом направлении произошло ослабление локомоторной роли мантийно-вороночного комплекса, основным органом движения стали плавники, а основной функцией гладиуса стала их поддержка. Снижение общей активности вампиров и их уход от активной конкуренции в глубоководный рефугиум привело к «консервации» многих архаичных черт организации этих удивительных животных. В результате гладиус современного вампира поразительно сходен с гладиусами юрских ископаемых теутид.

Рис. 14. Эволюция вампироподного ствола Coleoidea.
Другая линия вампиропод эволюционировала в направлении бенто-пелагических форм, плавающих с помощью плавников (Cirrata). Гладиус в этой линии претерпел редукцию осевой пластинки проостракума и превратился в подковообразную структуру, служащую поддержкой плавников. Дальнейший переход осьминогов к бентосному образу жизни и хождению на руках (Incirrata) привел к отказу от плавникового движения и редукции раковины до двух палочек-стилетов, служивших опорой вороночным ретракторам. В ходе последующей эволюции в ряде линий бесплавниковых осьминогов (Ameloctopus, Hapalochlaena и др.) гладиус полностью редуцировался в связи с ослаблением или утерей способности к реактивному плаванию (рис. 14). Важно отметить, что полная утрата раковины у продвинутых представителей Incirrata не повлекал за собой изменения плана их строения, который остался таким же, как и у примитивных представителей этого таксона.
На первый взгляд, идея о происхождении вампиропод от древних представителей сепиоидной ветви кажется экстравагантной и фантастичной. Уж очень отличаются между собой современные представители этих двух весьма многочисленных групп, каждая из которых имеет свой характерный «экологический облик». Однако привлечение данных по морфологии раковины позволяет достаточно уверенно говорить о филогенетической близости современных вампиротеутид и юрских теудопсид. Кроме того, происхождение вампиропод от древних бесфрагмоконных сепиоидей объясняет ряд сходств современных вампиропод и некоторых представителей сепиоидного ствола: срастание переднего дорсального края мантии с головой; широкое расположение звездчатых ганглиев; редукция проостракума, наличие вентрального аддуктора; субтерминальное положение раковины, обусловленное задним выпячиванием мантии.
XII. ЗАКЛЮЧЕНИЕ: СИСТЕМА И ФИЛОГЕНИЯ ПОДКЛАССА COLEOIDEA
Изложенная в главе XI точка зрения на эволюцию и филогению колеоидей предполагает соответствующее изменение их системы. Поскольку признаки строения раковины можно проследить и у современных, и у ископаемых моллюсков, предлагаемая новая система объединяет ранее разрозненные системы вымерших и рецентных форм. Новые таксоны сопровождаются диагнозами. Система подкласса Coleoidea, построенная с учетом строения их раковины, имеет следующий вид:
Подкласс COLEOIDEA Bather, 1888
Надотряд BELEMNOIDEA Hyatt, 1884 (?девон; карбон – мел)
Отряд AULACOCERIDA Stolley, 1919 (?девон; карбон – юра)
(сем. HEMATITIDAE, BOLETZKYIDAE, AULACOCERATIDAE,
MUTVEICONIDAE, DICTYOCONIDAE, PALAEOBELEMNOPSIDAE,
XIPHOTEUTHIDAE, CHITINOTEUTHIDAE)
Отряд BELEMNITIDA Zittel, 1895 (юра – мел)
(сем. PASSALOTEUTHIDAE, SALPINGOTEUTHIDAE, HASTITIDAE,
CYLINDROTEUTHIDAE, OXYTEUTHIDAE, BELEMNOPSEIDAE,
DICOELITIDAE, PSEUDODICOELITIDAE, DUVALIIDAE, BELEMNITELLIDAE, DIMITOBELIDAE, BELEMNOTHEUTIDAE, DIPLOBELIDAE)
Отряд PHRAGMOTEUTHIDA Jeletzky, 1965 (пермь – юра)
(cем. PHRAGMOTEUTHIDAE, DONOVANICONIDAE)
Надотряд SPIRULOIDEA superordo nov. (мел – настоящее время)
Отряд SPIRULIDA Haeckel, 1896 (мел – настоящее время)
(сем. ADYGEYIDAE, GROENLANDIBELIDAE, VASSEURIDAE,
BELEMNOSEIDAE, BELOPTERIDAE, SPIRULIROSTRIDAE, SPIRULIDAE)
Отряд CHIROTEUTHIDA ordo nov. (настоящее время)
(сем. CHIROTEUTHIDAE, MASTIGOTEUTHIDAE, JOUBINITEUTHIDAE,
BATOTEUTHIDAE, MAGNAPINNIDAE, PROMACHOTEUTHIDAE)
Надотряд SEPIOIDEA superordo nov. (юра – настоящее время)
Отряд SEPIIDA Zittel, 1895 (юра – настоящее время)
(сем. TRACHITEUTHIDAE, BELOSEPIELLIDAE, BELOSEPIIDAE, SEPIIDAE)
Отряд SEPIOLIDA Keferstein, 1866 (юра – настоящее время)
(сем. MUENSTERELLIDAE, SEPIOLIDAE, SEPIADARIIDAE, IDIOSEPIIDAE)
Отряд LOLIGINIDA ordo nov. (юра – настоящее время)
(сем. TEUDOPSIDAE, PALAEOLOLIGINIDAE, LOLIGINIDAE,
PICKFORDIATEUTHIDAE, AUSTRALITEUTHIDAE,
CHTENOPTERYGIDAE, BATHYTEUTHIDAE)
Надотряд TEUTHOIDEA superordo nov. (поздний триас – наст. время)
Отряд LOLIGOSEPIIDA Jeletzky, 1965 (поздний триас – наст. время)
(сем. LOLIGOSEPIIDAE, GEOPELTIDAE, LIOTEUTHIDAE,
THYSANOTEUTHIDAE)
Отряд OMMASTREPHIDA ordo nov. (юра – настоящее время)
(сем. PLESIOTEUTHIDAE, OMMASTREPHIDAE)
Отряд CRANCHIIDA ordo nov. (юра – настоящее время)
(сем. MASTIGOPHORIDAE, CRANCHIIDAE)
Отряд ONYCHOTEUTHIDA ordo nov. (поздняя юра – наст. время)
(сем. LEPTOTEUTHIDAE, EOTEUTHOIDAE, ONYCHOTEUTHIDAE,
ARCHITEUTHIDAE, GONATIDAE, NEOTEUTHIDAE, LYCOTEUTHIDAE,
ENOPLOTEUTHIDAE, ANCISTROCHEIRIDAE, PYROTEUTHIDAE,
PSYCHROTEUTHIDAE, HISTIOTEUTHIDAE, LEPIDOTEUTHIDAE,
BRACHIOTEUTHIDAE, PHOLIDOTEUTHIDAE, OCTOPOTEUTHIDAE,
CYCLOTEUTHIDAE)
Надотряд VAMPYROPODA Boletzky, 1992 (мел – настоящее время)
Отряд VAMPYROTEUTHIDA Jeletzky, 1965 (настоящее время)
(сем. VAMPYROTEUTHIDAE)
Отряд OCTOPODA Leach, 1817 (мел–настоящее время)
Подотряд CIRRATA Grimpe, 1916 (настоящее время)
(сем. OPISTHOTEUTHIDAE, STAUROTEUTHIDAE, CIRROTEUTHIDAE)
Подотряд INCIRRATA Grimpe, 1916 (мел – настоящее время)
(сем. PALAEOCTOPODIDAE, OCTOPODIDAE, AMPHITRETIDAE,
ALLOPOSIDAE, TREMOCTOPODIDAE, OCYTHOIDAE, ARGONAUTIDAE,
BOLITAENIDAE, VITRELEDONELLIDAE)
Пути эволюции и филогения колеоидей. Сравнительный анализ раковин ископаемых и рецентных Coleoidea позволяет сделать вывод о том, что современная фауна внутрираковинных головоногих представлена потомками четырех филогенетических ветвей: спирулоидной, сепиоидной, теутоидной и вампироподной. Пятая ветвь – белемнитиды – появилась на рубеже триаса и юры, пережила период расцвета в юре - мелу и полностью вымерла на рубеже мела и кайнозоя. Все филогенетические ветви колеоидей имеют общий корень (пермско-триасовые Aulacocerida), но разошлись на ранних этапах эволюции подкласса.
Анализ эволюции и филогении подкласса Coleoidea показывает, что гладиус – внутренняя декальцинированная раковина без фрагмокона – появлялся неоднократно и независимо во всех четырех эволюционных ветвях колеоидей: спирулоидной, сепиоидной, теутоидной и вампироподной. Можно сказать, что колеоидей «создавали» гладиус каждый раз, когда в этом появлялась потребность. Из этого заключения неизбежно следует другой важный вывод: «кальмары» в их современном понимании являются не систематической, а экологической категорией, не таксоном, а жизненной формой. Бесфрагмоконные колеоидеи, объединяемые в настоящее время в отряд Teuthida, Naef, 1916, представляют собой полифилетическую группу, включающую потомков теутоидной, сепиоидной и спирулоидной ветвей. Логическим выводом из этого является разделение нынешнего отряда Teuthida на три таксона отрядного уровня и отнесение их к различным надотрядам в новой системе Coleoidea.
Подводя итог анализу эволюции формы и функции раковины в подклассе Coleoidea, нельзя не отметить важную деталь: эволюция локомоторного аппарата Coleoidea повторяла в основных чертах принципиальные морфо-функциональные решения, реализованные в эволюции локомоторного аппарата примитивных бесчелюстных панцирных хордовых, предков рыб. Однако у колеоидей она проходила на основе принципиально иного – моллюскового – плана строения. Параллелизм в эволюции колеоидей и примитивных позвоночных выразился в следующем: 1) и у колеоидей, и у примитивных рыб сформировался внутренний продольный опорный элемент, соразмерный с длиной тела: гладиус и нотохорд, соответственно; 2) в обеих группах вокруг продольного опорного элемента развился внутренний хрящевой скелет, обеспечивающий прикрепление мышц локомоторного аппарата и защиты головного мозга; 3) на основе этого опорного комплекса сформировались органы локомоции, использующие ундуляционный способ движения: мускулистые плавники у колеоидей; хвостовой плавник – у примитивных позвоночных; 4) формирование внутреннего скелета сопровождалось редукцией скелета наружного: у головоногих это выражалось в трансформации наружной раковины во внутреннюю; у примитивных позвоночных – в редукции обширного внешнего костного панциря, имевшегося у примитивных остракодерм (Pteraspidomorphi, Galeaspida и др.).
К принципиальным отличиям головоногих и рыб, обусловленным различиями в исходных планах строения, относятся: 1) мономерное строение локомоторной мускулатуры головоногих и метамерное у рыб; 2) различные исходные способы движения: реактивный у головоногих; ундуляционный – у рыб.
ВЫВОДЫ
- Проостракум – опорный элемент мантийно-вороночного комплекса и плавников – является важнейшим эволюционным приобретением внутрираковинных головоногих моллюсков. В линиях, утративших фрагмокон, проостракум стал основным структурным элементом раковины, и ее дальнейшая эволюция шла главным образом как эволюция проостракума.
- У колеоидей имеются два типа зон контакта мышц с раковиной: первичные, унаследованные от наружнораковинных предков, и вторичные, появившиеся после погружения раковины внутрь тела. Первичные контакты – непосредственное прирастание мышц-ретракторов к раковине – весьма консервативны. Напротив, вторичные контакты – опосредованное взаимодействие мышц и раковины через раковинный мешок – очень разнообразны, но всегда осуществляются с участием хряща.
- Мускулы-ретракторы современных головоногих моллюсков – медиальные и латеральные сегменты ретракторов головы, ретракторы воронки, затылочные ретракторы у Nautilus – происходят от дорсо-вентральных мускулов примитивных моллюсков.
- Современная фауна внутрираковинных головоногих моллюсков представлена потомками четырех эволюционных ветвей: спирулоидной, сепиоидной, теутоидной и вампироподной. Представители разных ветвей различаются строением проостракума и фрагмокона (в случае редукции последнего – строением конотеки).
- Во всех эволюционных ветвях подкласса Coleoidea прослеживается тенденция к редукции раковины, проходящая через этапы утраты фрагмокона, превращения раковины в декальцинированный гладиус, а в некоторых линиях – и утраты самого гладиуса. Окончательная потеря раковины происходит у бентосных (Incirrata, Sepiida) или планктонных форм (некоторые Incirrata, Sepiolida, Cranchiidae) и сопровождается потерей способности к гидрореактивному движению. Потеря раковины у колеоидей не влекла за собой изменения плана строения, который у безраковинных форм остался таким же, как у близкородственных форм с раковиной.
- Разработана новая система подкласса Coleoidea, включающая как современных, так и ископаемых представителей этого подкласса.
Список основных публикаций по теме диссертации
- Филиппова Ю.А., Бизиков В.А., 1983. Систематическое значение скелетов современных кальмаров. Тезисы докл. Всес. Конференции: «Систематика и экология головоногих моллюсков». Ленинград: изд. ЗИН АН СССР, с. 45-46.
- Алексеев Д.О., Бизиков В.А. 1986. Некоторые аспекты биологии и экологии командорского кальмара Berryteuthis magister (Gonatidae) в районе острова Симушир в январе 1985 г. Ресурсы и перспективы использования кальмаров Мирового океана. Изд-во ВНИРО: с.50-57.
- Алексеев Д.О., Бизиков В.А., Хромов Д.Н. 1986. Подводные наблюдения за командорским кальмаром. Тезисы докл. IV Всесоюзной конф. по промысловым беспозвоночным. Ч.1. М., изд-во ВНИРО: с.126-127.
- Бизиков В.А. 1987. Рекомендации по развитию промысла командорского кальмара в северо-западной части Тихого океана и состояние его запасов. М., изд-во ВНИРО, - 15с.
- Бизиков В.А. 1987. Новые данные о строении гладиуса кальмаров. Зоологический журнал, т. LXVI, вып.2: с.177-184.
- Бизиков В.А. 1987. Морфо-функциональные аспекты строения гладиуса кальмаров. Тезисы докладов VIII Всес. совещания: Моллюски: результаты и перспективы их исследований: Тезисы докл. - Ленинград: «Наука»: с. 214-215.
- Алексеев Д.О., В.А. Бизиков и Д.Н. Хромов. 1987. Эволюция раковины в подклассе Coleoidea (Cephalopoda). Тезисы докладов VIII Всес. совещания: Моллюски: результаты и перспективы их исследований: Тезисы докл. - Ленинград: «Наука»: с. 52-53.
- Алексеев Д.О., Бизиков В.А., Хромов Д.Н. и Помозов А.А. 1989. Подводные наблюдения за поведением и распределением командорского кальмара и других головоногих моллюсков северной части Тихого океана. Подводные исследование в биоокеанологических и рыбохозяйственных целях: Сб. науч. трудов. – М.: изд-во ВНИРО: с.66-77.
- Бизиков В.А. 1990. Новый метод изучения индивидуального роста кальмаров. Авторское свидетельство № 1565440(51)5 А 01 К 61 00. Бюлл. «Открытия и изобретения». №19: с.25.
- Алексеев Д.О., Бизиков В.А. 1990. Эволюция хитиновых структур (гладиусов и роговых присосок) в семействе Loliginidae (Cephalopoda). Тез. докл. на V Всес. конф. по промысловым беспозвоночным. – М.: изд-во ВНИРО – с.77-78.
- Bizikov V.A. 1991. A new method of squid age determination using the gladius. In: Jereb,P., Ragonese,S. and Boletzky,S. (eds), Squid age determination using statoliths. N.T.R.-I.T.T.P. Special publications №1. Mazara del Vallo, Italy, pp. 39-51.
- Arkhipkin A.I., Bizikov V.A. 1991. Comparative analysis of age estimates using statolith and gladius in squids. In: Jereb,P., Ragonese,S. and Boletzky,S. (eds), Squid age determination using statoliths. N.T.R.-I.T.T.P. Special publications №1. Mazara del Vallo, Italy, pp.19-33.
- Jackson, G.D., Arkhipkin A.I., Bizikov V.A., and Hanlon R.T. 1993. Laboratory and field corroboration of age and growth from statoliths and gladii of the loliginid squid Sepioteuthis lessoniana (Mollusca; Cephalopoda). In: Okutani T., O'Dor, R.K. and Kubodera T. (eds.), Recent Advances in Cephalopod Fisheries Biology (Tokai University Press, Tokyo), pp. 189-199.
Bizikov V.A. 1995. Growth of Sthenoteuthis oualaniensis, using a new method based on gladius microstructure. ICES mar. Scientific Symposium. - №199, pp.445-458.
- Бизиков В.А. 1996. Атлас морфологии и анатомии гладиуса кальмаров. – М. Изд-во ВНИРО, - 248 с.
- Бизиков В.А. 1996. Размерно-половой состав уловов командорского кальмара Berryteuthis magister (Oegopsida; Cephalopoda) в северо-западной части Берингова моря в 1995 г. В кн.: Елизаров А.А. (ред.) «Промысловые аспекты биологии командорского кальмара и рыб склоновых сообществ в западной части Берингова моря». – М.: изд-во ВНИРО: с. 43-74.
- Бизиков В.А. 1996. Межгодовая изменчивость размерно-полового состава командорского кальмара в северо-западной части Берингова моря в 1993-1995 гг. В кн.: Елизаров А.А. (ред.) «Промысловые аспекты биологии командорского кальмара и рыб склоновых сообществ в западной части Берингова моря». – М.: изд-во ВНИРО: с. 74-81.
- Бизиков В.А. 1996. Распределение и запас командорского кальмара в северо-западной части Берингова моря. В кн.: Елизаров А.А. (ред.) «Промысловые аспекты биологии командорского кальмара и рыб склоновых сообществ в западной части Берингова моря». – М.: изд-во ВНИРО: с. 82-86.
- Бизиков В.А., Архипкин А.И. 1996. Возрастная структура молоди командорского кальмара в северо-западной части Берингова моря. В кн.: Елизаров А.А. (ред.) «Промысловые аспекты биологии командорского кальмара и рыб склоновых сообществ в западной части Берингова моря». – М.: изд-во ВНИРО: с. 96-101.
- Бизиков В.А., Архипкин А.И. 1996. Структура запаса командорского кальмара и её сезонная динамика в северо-западной части Берингова моря. В кн.: Елизаров А.А. (ред.) «Промысловые аспекты биологии командорского кальмара и рыб склоновых сообществ в западной части Берингова моря». – М.: изд-во ВНИРО: с. 142-143.
- Бизиков В.А. 1996. Сезонная и межгодовая изменчивость размерно-половой и возрастной структуры уловов командорского кальмара в северо-западной части Берингова моря. В кн.: Елизаров А.А. (ред.) «Промысловые аспекты биологии командорского кальмара и рыб склоновых сообществ в западной части Берингова моря». – М.: изд-во ВНИРО: с. 144-145.
- Бизиков В.А., Архипкин А.И., Верхунов А.В. 1996. Размерно-возрастная структура, вертикальное распределение и возможные пути миграции молоди командорского кальмара в северо-западной части Берингова моря. В кн.: Елизаров А.А. (ред.) «Промысловые аспекты биологии командорского кальмара и рыб склоновых сообществ в западной части Берингова моря». – М.: изд-во ВНИРО: с. 145-148.
- Бизиков В.А. 1996. Межгодовые изменения в распределении и численности командорского кальмара в северо-западной части Берингова моря. В кн.: Елизаров А.А. (ред.) «Промысловые аспекты биологии командорского кальмара и рыб склоновых сообществ в западной части Берингова моря». – М.: изд-во ВНИРО: с. 148-150.
- Бизиков В.А., Архипкин А.И., 1996. Распределение, структура запаса и перспективы промысла командорского кальмара. Рыбное хозяйство. №1: с. 42-45.
- Arkhipkin, A.I., Bizikov, V.A., Krylov, V.V. and Nesis, K.N., 1996. Distribution, stock structure, and growth of the squid Berryteuthis magister (Berry, 1913) (Cephalopoda, Gonatidae) during summer and fall in the western Bering Sea. Fishery Bulletin, U.S., 94 (1), pp. 1-30.
- Arkhipkin A.I., Bizikov V.A. 1996. Possible imitation of jellyfish by the squid paralarvae of the family Gonatidae (Cephalopoda, Oegopsida). Polar Biology. 16, pp. 531-534.
- V.A. Bizikov, A.I. Arkhipkin. 1997. Morphology and microstructure of the gladius and statolith from the boreal Pacific giant squid Moroteuthis robusta (Oegopsida; Onychoteuthidae). Journal of Zoology, London. 241: 475-492.
- Филиппова Ю.А., Д.О. Алексеев, В.А. Бизиков и Д.Н. Хромов. 1997. Справочник-определитель промысловых и массовых головоногих моллюсков Мирового океана. – М. Изд-во ВНИРО. – 272 с.
- A.I. Arkhipkin, V.A. Bizikov., 1997. Statolith shape and microstructure in studies of systematics, age and growth in planctonic paralarvae of gonatid squids (Cephalopoda, Oegopsida) from the western Bering Sea. – Journal of Plancton Research 19, pp. 1993-2030.
- A.I. Arkhipkin, V.A. Bizikov, A.V. Verkhunov. 1998. Distribution and growth in juveniles of the squid Berryteuthis magister (Cephalopoda; Gonatidae) in the western Bering Sea. – Sarsia 83, pp. 45-54.
- Бизиков В.А., Филиппова Ю.А., 1999. Российский промысел головоногих моллюсков имеет шансы на возрождение. Рыбное хозяйство. №1: с. 32-35.
- Arkhipkin A.I. and V.A. Bizikov. 2000. Role of the statoliths in functioning of the acceleration receptor system in squids and sepioids. Journal of Zoology, London, 250, pp. 31-55.
- Arkhipkin A.I., Bizikov V.A. 1998. Statoliths in accelerometers of squids and cuttlefish. Rutenica 8(1), pp. 81-84.
- Arkhipkin A.I., Bizikov V.A. 1999. Statolith shape as an indicator of the life>
- Bizikov V.A. 2000. Functional morphology and evolution of the shell in Coleoid cephalopods. In: Abstracts of CIAC Symposium ‘Cephalopod Biomass & Production. University of Aberdeen, Scotland. July 3d -7th, 2000: p. 88-89.
- Bizikov V.A. 2002. Reanalysis of functional design of Nautilus locomotory and respiratory systems. American Malacological Bulletin, Vol. 17(1/2), pp. 17-30.
- Bizikov V.A. 2004. The shell in Vampyropoda (Cephalopoda): morphology, functional role and evolution. Ruthenica, Supplement 3, 2004. – 88 pp., 57 figs.
- Bizikov V.A. 2004. Evolution of the shell in Octopodiformes. Тезисы докладов конференции "Моллюски северо-восточной Азии и Северной Пацифики: биоразнообразие, экология, биогеография и история фауны", ИБМ ДВО РАН, Владивосток, 4-6 октября 2004: с.122-123 (на англ.).
- Рогов М.А., Бизиков В.А. 2004. Ископаемые теутиды в коллекциях государственного геологического музея им. В.И. Вернадского. Проблемы региональной геологии: музейный ракурс (Материалы научно-практической конференции). М.: Акрополь, 2004: 176-178.
- Rogov M.A. and V.A. Bizikov. 2006. New data on mddle Jurassic-lower Cretaceous Belemnotheutidae from Russia. What can shell tell about the animal and its mode of life. Acta Universitatis Carolinae – Geologia. Vol. 49, pp. 155-169.
- Бизиков В.А. 2008. Эволюция формы и функции раковины головоногих моллюсков (Coleoidea; Cephalopoda). Ruthenica, Supplement **, 2008. – 500 с.