

Фаунистическое и экологическое разнообразие панцирных клещей (acariformes: oribatei) в экосистемах кольского севера
На правах рукописи
ЛИСКОВАЯ
Алена Анатольевна
Фаунистическое и экологическое разнообразие
панцирных клещей (Acariformes: Oribatei)
в экосистемах Кольского Севера
03.02.04 – зоология
03.02.08 – экология
Автореферат диссертации на соискание учёной степени кандидата
биологических наук
Петрозаводск – 2011
Работа выполнена в Институте проблем промышленной экологии Севера Кольского научного центра Российской академии наук
| Научные руководители | доктор биологических наук профессор заслуженный эколог России Евдокимова Галина Андреевна |
| кандидат биологических наук доцент Зенкова Ирина Викторовна |
Официальные оппоненты доктор биологических наук
профессор
Иешко Евгений Павлович
кандидат биологических наук
Мелехина Елена Николаевна
Ведущая организация Институт проблем экологии и эволюции
им. А.Н. Северцова РАН
Защита диссертации состоится 23 марта 2011 г. в 14 часов на заседании диссертационного совета Д 212.190.01 при Петрозаводском государственном университете по адресу: 185910, Республика Карелия, Петрозаводск, пр. Ленина, 33; эколого-биологический факультет.
С диссертацией можно ознакомиться в библиотеке Петрозаводского государственного университета. Электронная версия автореферата размещена на официальном сайте http://www.petrsu.ru.
Автореферат разослан « 15 » февраля 2011 г.
| Ученый секретарь диссертационного совета канд. биол. наук |  | И.М. Дзюбук |
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность. Панцирные клещи или орибатиды (Acariformes: Oribatida) – эволюционно древняя, широко распространенная и чрезвычайно разнообразия в таксономическом и экологическом отношении группа клещей. В почвах северных широт эти микроартроподы преобладают по обилию и видовому разнообразию над представителями мезофауны. Зоо-микробиальные взаимосвязи орибатид с мицелиальными грибами, определяют значимую роль этих клещей в процессах биотрансформации органического вещества в экосистемах полярных и приполярных регионов, включая зональные почвы Кольского Севера, в которых грибы являются ведущей группой микроорганизмов-деструкторов (Евдокимова, Мозгова, 2001).
На территории области, несмотря на небольшую протяженность с юга на север (~400 км), выделяют пять климатических районов (Яковлев, 1961). Здесь выражена зональная смена северотаежного, лесотундрового и тундрового типа растительности. В горах широтной зональности соответствует высотная поясность: горно-таежный пояс сменяется субальпийским (горно-лесотундровым), альпийским (горно-тундровым) и поясом арктических пустынь. В пределах региона на породах разного генезиса сформированы подзолы (иллювиально-железистые, иллювиально-гумусовые, торфяно-подзолы, глееподзолы), подбуры, торфяные почвы, литоземы, криоземы (Переверзев, 2007). В результате развитой горнодобывающей и перерабатывающей промышленности почвы региона подвергаются аэротехногенному загрязнению поллютантами различной природы; на обширных территориях складированы отходы обогащения полезных ископаемых.
Указанные факторы не могут не отражаться на фаунистическом и экологическом разнообразии панцирных клещей, сведения о котором ограничены данными лишь по двум климатическим районам: Терскому берегу и островам Белого моря, насчитывающим немногим более 100 видов (Бызова и др., 1986; Каталог биоты …, 2008) и побережью Баренцева моря, для которого ранее указывалось 79 видов орибатид (Криволуцкий, 1966; Панцирные клещи, 1995; Криволуцкий и др., 1999). Исследования последних лет, направленные на оценку вклада мигрирующих птиц в распространение клещей путем форезии, пополнили список орибатид Кольской тундры 44 видами, населяющими специфические местообитания баренцевоморского побережья – орнитогенные субстраты и гнезда птиц (Лебедева, Криволуцкий, 2003; Лебедева и др., 2006; Лебедева, Лебедев, 2007; Лебедев, 2009). Фауна орибатид основной территории региона, включающей широкий спектр природных и техногенно трансформированных биогеоценозов, а также горные экосистемы, остается практически неизученной.
Цель работы состояла в обобщении разрозненных литературных данных и результатов собственных исследований о фаунистическом и экологическом разнообразии, биоиндикационной значимости панцирных клещей в природных и техногенно трансформированных почвах и техногенных субстратах Мурманской области, а также составлении сводного списка орибатид этого заполярного региона.
Для достижения поставленной цели решали следующие задачи:
1. Дать сравнительную характеристику сообществ орибатид в природных почвах Мурманской области с учетом факторов широтной зональности (почвы тундры, лесотундры и подзоны северной тайги) и высотной поясности (почвы Хибинского горного массива).
2. Сравнить сообщества орибатид в техногенно нарушенных почвах в зонах воздействия промышленных предприятий разного типа (медно-никелевый комбинат, алюминиевый завод). Выявить виды-индикаторы промышленного загрязнения.
3. Исследовать комплексы орибатид в техногенных субстратах на примере самозарастающих и рекультивированных отходах обогащения апатит-нефелиновых руд разного срока складирования.
Научная новизна. Впервые обобщены литературные данные и результаты собственных исследований фауны панцирных клещей в 11 районах Мурманской области, охватывающих естественные (равнинные и горные) и техногенно трансформированные экосистемы тундровой зоны и северотаежной подзоны. Региональный список орибатид увеличен на 79 видов (или 30%), из них 45 видов из 30 родов и 22 семейств указаны впервые. Значительно расширен видовой состав семейств Suctobelbidae, Carabodidae, Oppiidae, наиболее богатых по числу видов в данной области. Установлено влияние широтной зональности и разных механизмов расселения орибатид на их разнообразие, соотношение высших (Brachypylina) и низших (Macropylina) таксонов и соотношение одно-, мало- и многовидовых семейств. Определены виды с широким и локальным распространением в пределах области, а также виды, доминирующие по численности.
Теоретическая значимость. Выявленные виды панцирных клещей дополняют сведения по фауне Мурманской области и значительно расширяют имеющиеся сведения по разнообразию этой группы на территории Европейского Севера России, что важно с точки зрения инвентаризации и сохранения биоразнообразия почв северных регионов. Полученные результаты дают представление о механизмах расселения и формирования фауны клещей в высоких широтах, а также об особенностях экологии орибатид в зональных и техногенно нарушенных почвах Севера.
Практическая значимость. Установлены различные изменения структуры сообществ панцирных клещей в зависимости от степени техногенной нагрузки. Выявлены виды – положительные и отрицательные индикаторы загрязнения почв выбросами алюминиевого и медно-никелевого предприятий, а также виды, устойчивые к воздействию промышленных источников разного типа. Установлена значимость панцирных клещей как индикаторов микологической активности и накопления органического вещества в техногенных субстратах (нефелинсодержащих песках) в процессе первичного почвообразования. Выявлена индикаторная роль орибатид при оценке эффективности способов восстановления территории техногенной пустоши в окрестностях медно-никелевого комбината. Уточненный и дополненный список панцирных клещей Мурманской области опубликован в тематическом выпуске «Зеленый пояс Фенноскандии» Трудов Карельского НЦ РАН.
Апробация работы. Основные результаты исследований были доложены и получили положительную оценку на 8 научных конференциях с международным участием: «Наука и образование» (Мурманск 2008); «Актуальные проблемы сохранения биоразнообразия в экстремальных условиях северного климата» (Кировск 2008); «Сохранение биологического разнообразия наземных и морских экосистем в условиях высоких широт» (Мурманск 2009); «Сбалансированное природопользование» (Апатиты 2009); «Биогеграфия почв» (Москва 2009); «Современные проблемы загрязнения почв» (Москва 2010); «Экологические проблемы северных регионов и пути их решения» (Апатиты 2010); XV Всероссийское совещание по почвенной зоологии (Москва 2008).
Публикации. По результатам диссертации опубликовано 13 работ, в том числе две статьи в рецензируемых журналах из списка, рекомендованного ВАК.
Структура и объем диссертации. Диссертация состоит из введения, 7 глав, выводов, 6 приложений с видовым списком панцирных клещей Мурманской области, перечня цитируемой литературы, включающего 233 источника, в т.ч. 92 на иностранных языках. Работа изложена на 178 страницах, содержит 57 рисунков, 55 таблиц.
Благодарности. Автор выражает искреннюю благодарность д.б.н., профессору Г.А. Евдокимовой и к.б.н., доценту И.В. Зенковой за руководство работой, идеи и постоянную помощь при подготовке диссертации, за предоставленные данные по микробиологическим и физико-химическим свойствам исследованных почв; к.б.н. Л.Г. Исаевой за предоставленные данные и консультации.
Особую благодарность за таксономическое определение панцирных клещей, предоставленные данные и персональные консультации выражаем к.б.н. Л.М. Ласковой, к.г.н. А.С. Зайцеву, к.г.н. Е.А. Сидорчук и Л.В. Залиш.
СОДЕРЖАНИЕ РАБОТЫ
1. СИСТЕМТИческое положение И ЭКОЛОГИЯ ПАНЦИРНЫХ клещей
Приведены сведения о систематическом положении, таксономическом разнообразии, особенностям биологи и экологии панцирных клещей, их использовании в зоологической индикации почв. Показана степень изученности фауны панцирных клещей на территории Мурманской области.
2. ХАРАКТЕРИСТИКА МУРМАНСКОЙ ОБЛАСТИ
Рассмотрено разнообразие природных условий и охарактеризованы типы промышленного загрязнения почв Мурманской области как факторов, определяющих биоразнообразие региональной фауны панцирных клещей.
3. ОБЪЕКТЫ И МЕТОДЫ
Исследования охватили 11 районов Мурманской области от побережья Баренцева моря (тундрово-лесотундровая флористическая область, районы 1-3) до побережья Кандалакшского залива Белого моря (районы 10, 11), включая природные, техногенные и горные экосистемы центральной части региона (подзона северной тайги, районы 4-9) (рис. 1, табл. 1). Собственные исследования выполнены в 4 районах: 4-7. Материалы по районам 1, 2, 8, 9 предоставлены сотрудниками ИППЭС Кольского НЦ РАН. По районам 3, 10, 11 использованы литературные данные (Криволуцкий, 1966; Бызова и др., 1986; Панцирные клещи, 1995; Криволуцкий и др., 1999; Лебедева, Криволуцкий, 2003; Лебедева и др., 2006; Лебедева, Лебедев, 2007; Лебедев, 2009). Видовой состав панцирных клещей приведен для 9 районов: 1, 2, 3, 5, 6, 8, 9, 10, 11. Виды, выявленные в районах 4 и 7, будут идентифицированы позднее. Всего исследовано 43 биотопа.
 | Рис. 1. Районы исследований: 1 –пос. Видяево; 2 – пос. Териберка; 3 – пос. Дальние Зеленцы; 4, 5 –участки в 1 и 5 км от медно-никелевого комбината «Североникель» (г. Мончегорск); 6 – Хибинский горный массив, 7 –хранилища нефелинсодержащих песков, 8 – пос. Уполокша, зональные сосняки, 9 – зона в 2 км от алюминиевого завода (КАЗ, г. Кандалакша), 10 – беломорские о-ва Кандалакшского заповедника, 11 – Терский берег Белого моря. |
Таблица 1. Характеристика исследованных районов
| № | Коорд. | Экосистемы, почвы |
| Тундрово-лесотундровая флористическая область | ||
| 1 | 69°19 с.ш. 32°52 в.д. | Березняк лесотундровый кустарничково-лишайниковый; вершина скалы, 60 м над ур. м. Почвы: Al-Fe-гумусовые подзолы, торфяно-подзолы. |
| 2 | 69°10' с.ш. 35°08' в.д. | Горные тундра лишайниково-кустарничковая и березняк лесотундровый воронично-лишайниковый, 40-60 м над ур. м. Подзолы Al-Fe-гумусовые. |
| 3 | 69°07' с.ш. 36°03' в.д. | Тундра кустарничково-лишайниковая, 50-80 м над ур. м. Подзолы Al-Fe-гумусовые оторфованные. |
| Подзона северной тайги | ||
| 4 | 67°56' с.ш. 32°49' в.д. | Площадки по восстановлению техногенной пустоши в 1 км от комбината «Североникель», 138-177 м над ур. м. |
| 5 | 67°34' с.ш. 34°17' в.д. | Импактная зона в 5 км от комбината «Североникель», 80 м над ур. м. Подзолы Al-Fe-гумусовые оторфованные, сильно эродированные. В верхнем слое содержится (мг/кг): Ni 1700-2600, Cu 600-1000, Co 300-350, сульфат-ионов 400-600. Зольность достигает 80-90%, pH = 3.5. |
| 6 | Хибинский горный массив (ХГМ), 6 биотопов | |
| 67°38' с.ш. 33°39' в.д. | Гора Вудъяврчорр, склон северо-восточной экспозиции. Субальпийский пояс березового криволесья (В-СВЭ-бк), 393 м над ур. м. Подзол иллювиально-многогумусовый. | |
| 67°38' с.ш. 33°39' в.д. | Гора Вудъяврчорр, склон северо-восточной экспозиции. Горная тундра кустарничковая, 434 над ур. м. (В-СВЭ-т/куст). Почва скрытоподзолистая иллювиально-многогумусовая оторфованная. | |
| 67°38' с.ш. 33°40' в.д. | Гора Вудъяврчорр, южный склон. Предгорная ерниковая тундра (В-ЮЭ-т/ерн), 324 м над ур. м. | |
| 67°10' с.ш. 33°12' в.д. | Западная часть ХГМ, правый берег реки Малая Белая, горно-таежный пояс (ХГМ-ЗЧ-гт). | |
| 67°14' с.ш. 33°34' в.д | Восточная часть ХГМ, пойма реки Тульйок, горно-таежный пояс (ХГМ-ВЧ-гт), 280 м над ур. м. | |
| 67°10' с.ш. 38°00' в.д. | Гора Ловчорр, западный склон. Тундра каменистая с фрагментарным лишайниково-кустарничковым покровом (Л-ЗЭ-т/кам). | |
| 7 | 67°35' с.ш. 33°35' в.д. | Хранилища с нефелинсодержащими песками разного срока хранения: свеженамытые (срок экспозиции <1 года); самозарастающие мхами и закрепленные методом посева многолетних злаков более 10-, 20- и 30-ти лет назад; рекультивированные более 40 лет назад – новообразованные почвы под лесной и луговой растительностью. |
| 8 | 67°33' с.ш. 31°54' в.д. | Зональный сосняк вороничный. Подзолы Al-Fe-гумусовые на песчаной морене. Содержание углерода – 52% от сухого вещества, азота – 1.43%. |
| 9 | 67°09' с.ш. 32°24' в.д. | Сосняк вороничный в зоне сильного загрязнения в 2 км от КАЗ, функционирующего с 1951 г. Древостой разреженный, с отмершими ветками и пожелтевшей хвоей в нижней и средней частях кроны. Подзолы Al-Fe-гумусовые в состоянии техногенной трансформации. Содержание фтора в подстилке до 1200 мг/кг. Органическое вещество подстилки обеднено углеродом и азотом: содержание С-27% от сухого вещества, N-0.63%. Зольность повышена до 60%, величина pH – до 5.5. |
| 10, 11 | 66o34' с.ш. 33o08' в.д. | О-ва Северного Архипелага и Терское побережье Белого моря. Сосновые, еловые, березовые леса, приморские луга, полоса галофитов, наскальная растительность (31 биотоп). Подзолы, торфяно-подзолы. |
Клещи были извлечены из почвенных проб размером 2525 см, отобранных из органогенного горизонта 0-5 см, а также из образцов с растительными остатками (лесная подстилка, листья березы, хвоя сосны), заложенными в органогенный горизонт в мешочках из нейлоновой сетки с диаметром отверстий 11 мм. В районах 4 и 6 образцы отбирали металлическим цилиндром диаметром 10 см и высотой 5 см. Для выгонки клещей использовали электропрогрев по методу эклекторов Тулльгрена в течение нескольких суток до полного высушивания субстратов. В период с мая 2007 по июнь 2010 года выполнено 16 полевых выездов. Собрано и обработано более 860 образцов. Экстрагировано и определено около 19.4 тыс. экземпляров панцирных клещей.
Одновременно был выполнен анализ физико-химических свойств субстратов: температуры, влажности, зольности, кислотности (4 тыс. определений). Связь фаунистических и микробиологических показателей, почвенных свойств и показателей ферментативной активности проанализированы по результатам корреляций. Сходство видового состава клещей в исследованных районах оценивали с помощью кластерного анализа. В качестве базовой меры сходства взят индекс общности Серенсена Is. Рассчитывали прогнозное разнообразие орибатид по формуле: Ds= S(2-0.08n), n 11, где n – число исследованных районов, S – число видов в данном районе (Криволуцкий и др., 1999). Данные обрабатывали в программах Microsoft Excel (описательная статистика, корреляционный анализ) и Statistica 7.0 (кластерный анализ). Использовали трофическую классификацию панцирных клещей (Shuester, 1956; Wallwork, 1958; Xavier, Haq, 2007) и их подразделение на жизненные формы (Криволуцкий и др., 1990). Типы ареалов приведены в соответствии с системой Субиаса (Subias, 2010).
4. ФАУНА ПАНЦИРНЫХ КЛЕЩЕЙ ЗОНАЛЬНЫХ ПОДЗОЛОВ КОЛЬСКОГО СЕВЕРА
4.1. Панцирные клещи в почвах зональной тундры. Список панцирных клещей в окрестностях пос. Дальние Зеленцы (район 3), обобщенный по разным источникам, насчитывает 123 вида из 72 родов, 38 сем. В результате исследований районов 1 и 2 северо-западнее района 3 на побережье Баренцева моря, состав орибатид Кольской тундры пополнился 7 новыми видами и родом Diapterobates. Теперь он насчитывает 143 вида, 78 родов, 38 сем. Наиболее разнообразны 17 семейств (или 45%) с числом видов от 4 до 10, в первую очередь – сем. Brachychthoniidae. Треть семейств одновидовые.
В районе 1 в лесотундровых подзолах под березняками найдено 14 видов орибатид, принадлежащих к 12 родам, 11 сем. Виды Ph. anonymus, H. gibba и C. minusculus указываются для Мурманской области впервые. За исключением сем. Carabodidae (3 вида рода Carabodes) все семейства включали по 1 роду с единственным видом, что соответствует организации сообществ педобионтов, находящихся за пределами экологического оптимума (Ганин, 2006, 2009). Экстремальность условий местообитания подтверждалась супердоминированием наиболее эврибионтного среди орибатид вида T. velatus – 65% (рис. 2).
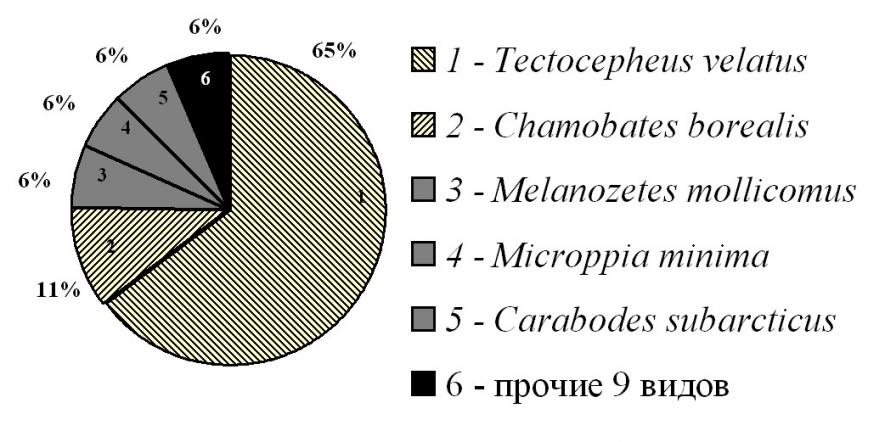 | Рис. 2. Структура доминирования панцирных клещей в почвах зональной тундры в окрестностях поселка Видяево. |
В горно-тундровых и лесотундровых формациях района 2 обнаружено 27 видов из 24 родов, 18 сем. При доминировании вида T. velatus (28%) шире был спектр как доминантов (8 видов), так и видов c обилием менее 1% каждый (19 видов). Структура сообщества была полидоминантной (рис. 3). Четыре малочисленных вида – Ph. crenophilus, M. translamellata, E. acromios и D. humeralis выявлены в регионе впервые. Таксономический статус нескольких интересных экземпляров требует уточнения. Возможно, в эти районы с Нордкапским течением проникают виды, характерные для атлантического побережья Европы.
 | Рис. 3. Структура доминирования панцирных клещей в почвах зональной тундры в окрестностях поселка Териберка. |
Общими для районов 1 и 2 были 6 видов (Is = 29%): T. velatus, N. coronata, C. traegardhi, H. initialis, C. subarcticus, C. forsslundi. Лесной ксерофильный вид C. forsslundi, известный из оперения птиц (Лебедева, 2005), отмечен только в этих районах (табл. 2).
Для трех тундровых районов побережья Баренцева моря общими оказались лишь 2 вида: T. velatus, широко распространенный по территории Мурманской области, и C. subarcticus, тяготеющий к зональным и горным тундрам региона и не выявленный в широком спектре островных местообитаний Белого моря. Подавляющее большинство – 125 видов (или 88%) встречены лишь в одном из трех районов, для трети из них (40 видов или 32%) известна форезия на птицах. Потенциально птицами может переноситься 55 видов или 40% орибатид Кольской тундровой зоны.
Таблица 2. Панцирные клещи, впервые указанные для Мурманской области
| Семейство | Род | Вид | Район |
| Achipteriidae | Achipteria | A. coleoptrata (Linneus 1758) | 8, 9 |
| Parhypochthoniidae | Parhypochthonius | P. aphidinus (Berlese 1904) | 9 |
| Camisiidae | Camisia | C. borealis (Hermann 1804) | 5 |
| Heminothrus | H. septentrionalis (Sellnick 1944) | 6 | |
| Euphthiracaridae | Mesotritia | M. flagelliformis (Ewing 1909) | 9 |
| Euphthiracarus | E.
monodactylus (Willmann 1919) | 8 | |
| Microtritia | M. minima (Berlese 1904) | 1 | |
| Trhypochthoniidae | Trhypochthonius | T. cladonicola (Willmann 1919) | 9 |
| Oppiidae | Medioppia | M. globosa (Miheli 1956) | 9 |
| Moritzoppia | M. unicarinata (Paoli 1908) | 8, 9 | |
| Scheloribatidae | Paraleius | P. leontonychus (Berlese 1910) | 9 |
| Hemileius | Hemileius sp. | 6 | |
| Phenopelopidae | Eupelops | E. acromios (Hermann 1804) | 2 |
| E. occultus (C.L. Koch 1836) | 5, 8, 9 | ||
| Haplozetidae | Protoribates | P. capucinus (Berlese 1908) | 6 |
| Phthiracaridae | Phthiracarus | Ph. anonymus (Grandjean 1933) | 1 |
| Ph. crenophilus (Willmann 1951) | 2 | ||
| Ph. laevigatus (C.L. Koch 1844) | 2, 8 | ||
| Ph. piger (Scopoli 1763) | 8 | ||
| Hermanniidae | Hermannia | H. gibba (C.L.Koch 1840) | 1 |
| Carabodidae | Carabodes | C. coriaceus (C.L. Koch 1835) | 8 |
| C. forsslundi (Sellnick 1953) | 1, 2 | ||
| C. minusculus (Berlese 1923) | 1 | ||
| C. reticulatus (Berlese 1913) | 8 | ||
| Liacaridae | Liacarus | Liacarus sp. | 2 |
| Suctobelbidae | Suctobelbella | S. alloenasuta (Moritz 1971) | 8 |
| S. baloghi (Forsslund 1958) | 8 | ||
| S. falcata (Forsslund 1941) | 8, 9 | ||
| S. forsslundi (Strenzke 1950) | 8, 9 | ||
| S. latirostris (Strenzke, 1950) | 1, 9 | ||
| S. perforata (Strenzke 1950) | 9 | ||
| S. singularis (Strenzke 1950) | 8, 9 | ||
| S. subtrigona (Oudemans 1900) | 8, 9 | ||
| Peloppiidae | Ceratoppia | C. quadridentata (Haller 1882) | 8 |
| Ceratozetidae | Ceratozetella | Ceratozetella sp. | 6 |
| Diapterobates | D. humeralis (Hermann 1804) | 2 | |
| Diapterobates sp. | 6 | ||
| Chamobatidae | Chamobates | Chamobates sp. | 6 |
| Damaeidae | Belba | Belba sp. | 6, 8, 9 |
| Epidamaeus | Epidamaeus sp. | 6, 8, 9 | |
| Eremaeidae | Euеremaeus | E. silvestris (Forsslund 1957) | 6, 8 |
| Galumnidae | Pergalumna | P. dorsalis (C.L. Koch 1841) | 8, 9 |
| Galumna | Galumna sp. | 6 | |
| Nanhermanniidae | Nanhermannia | N. sellnicki (Forsslund 1958) | 6 |
| Tectocepheidae | Tectocepheus | T. velatus sarekensis (Trgrdh 1910) | 6 |
| 22 семейства | 30 родов | 45 видов | 6 |
Примечание. Жирным шрифтом выделены впервые указанные семейства и роды.
4.2. Панцирные клещи в почвах зональной тайги (северотаежная подзона)
4.2.1. Панцирные клещи в почвах Хибинского горного массива. В 6 биоценозах таежного, субальпийского и альпийского высотных поясов выявлено 49 видов орибатид из 37 родов, 23 сем. (рис. 4). Сем. Haplozetidae (P. capucinus) и виды H. septentrionalis и N. sellnicki и T. v. sarekensis найдены только в почвах Хибин.
 | Рис. 4. Районы исследования в пределах Хибин: 1, 2 – западная и восточная части массива, 3 – западный склон кара оз. Ловчорр, 4 – г. Вудъяврчорр. |
Установлено большее фаунистическое сходство горных экосистем (горно-лесотундровых и горно-тундровых) по сравнению с предгорными (горно-таежными). Наиболее таксономически близкими оказались почвы субальпийского и альпийского поясов северо-восточного склона горы Вудъяврчорр: 10 общих видов из 7 родов (E. ribagai, H. longisetosus, H. septentrionalis, N. humicolus, N. selnicki, T. v. velatus, T. v. sarekensis, Ceratozetella sp., C. tienemanni, Scheloribates sp.); Is = 67%. Далее следовали пары одноименных поясов: тундровые местообитания на разных склонах горы Вудъяврчорр (8 общих видов из 7 родов; Is = 57%) и предгорные леса в западной и восточной частях ХГМ (15 общих видов из 15 родов; Is = 54%). Сходство видового состава орибатид убывало в ряду: соседние растительные пояса одного склона одной горы одноименные (тундровые) пояса на разных склонах одной горы одноименные (тундровые) пояса на разных горах = одноименные (предгорно-таежные) пояса в разных частях массива.
4.2.2. Панцирные клещи в почвах зональных сосняков. В сосняке вороничном в центральной части области (район 8) разнообразие орибатид насчитывало 30 видов панцирных клещей из 17 родов, 16 сем. Низкая степень доминирования самого массового вида T. velatus и широкий спектр как доминирующих (9), так и малочисленных видов (20) отражают многовидовую в условиях северной тайги, полидоминантную и сбалансированную структуру сообщества орибатид (рис. 5). Полидоминантность подтверждается высокими значениями индексов разнообразия Шеннона (4.2) и выравненности Пиелоу (0.86), а низкое обилие основного доминанта – малой величиной индекса доминирования Симпсона (0.07).
С учетом 23 видов, экстрагированных из опытных образцов с растительными остатками, заложенными в подстилку сосняка, список орибатид района 8 расширен до 39 видов из 22 родов, 19 сем. Виды E. monodactylus, Ph. piger, C. coriaceus, C. reticulates, S. alloenasuta, S. baloghi, C. quadridentata встречены только в этом районе (см. табл. 2).
 | Рис. 5. Полидоминантная структура сообщества панцирных клещей в подстилке зонального сосняка вороничного. |
4.2.3. Панцирные клещи Беломорских островов. На о-вах Кандалакшского заповедника (21 остров, 31 биотоп) выявлено 112 видов орибатид из 75 родов, 39 сем. Число видов в биотопах варьировало от 5 до 55 (Бызова и др., 1986). Разнообразие орибатид в этом районе связано с наличием специфических местообитаний (приморские луга, скопления галофитов, гнезда околоводных птиц, орнитогенные субстраты), отсутствующих в центральных районах области. В 10 биотопах найдено 17 «локально встречающихся» видов, т.е. обнаруженных в единственном местообитании. Беломорские о-ва включают половину всех биотопов региона, на которых найдены такие виды. Общими для островных биотопов оказались лишь 4 вида: O. tibialis, L. similis, T. velatus, C. areolatus. К лесным местообитаниям было приурочено 38% доминантных видов, к приморским – 34%.
5. ПАНЦИРНЫЕ КЛЕЩИ В ТЕХНОГЕННО НАРУШЕННЫХ ПОЧВАХ
КОЛЬСКОГО СЕВЕРА
Рассмотрено влияние на почвенную микрофауну хронического загрязнения лесных подзолов выбросами промышленных предприятий разного типа – комбината цветной металлургии «Североникель» и Кандалакшского алюминиевого завода.
5.1. Влияние медно-никелевого комбината
5.1.1. Изменение структуры населения микроартропод вдоль градиента загрязнения. Для 16 участков в 0.5-50 км от комбината «Североникель» в южном и юго-западном направлении по розе ветров показана зависимость изменения общей плотности микроартропод от численности орибатид (r = 0.92). Рост численности этих клещей с 85 экз./м2 в 0.5 км от комбината до 34 тыс. экз./м2 в подстилке фонового участка отрицательно коррелировал с зольностью подстилки (-0.50), содержанием Fe (-0.64), Ni, Cu, Co (r от -0.44 до -0.47) и величиной pH (-0.41). Положительными были связи с численностью микромицетов (0.46) и содержанием в почве Mn (0.68): наибольшее количество этого биофильного элемента (307±5 мг/кг) и максимальные скопления орибатид (114 тыс. экз./м2) и их нимф (13 тыс. экз./м2) под лишайниками при средней численности на участке 70 тыс. экз./м2 отмечены в 25 км от комбината (рис. 6).
 | Рис. 6. Изменение численности орибатид вдоль градиента загрязнения почв. |
На расстоянии до 1 км орибатиды выявлены в 60% почвенных образцов. Имаго встречались во всех образцах, начиная со 2 км, нимфы – лишь с 18 км. В зоне до 7 км клещи были приурочены к моховому покрову, с расстояния 15-км – к лишайникам, появляющимся в составе растительности.
По содержанию Ni, Cu, Co в органогенном горизонте предложено выделить зоны сильного (до 3 км), среднего (до 25 км) и слабого загрязнения (рис. 7) с соответствующей классификацией почв на сильно-, слабо- и нетоксичные для биоты (Евдокимова и др., 2011).

Рис. 7. Деление территории, подверженной воздействию выбросов комбината
«Североникель», на зоны по состоянию комплексов панцирных клещей.
На основании данных по численности, встречаемости и обилию (%) панцирных клещей и их нимф выделены 4 зоны: 0-2 км (численность орибатид <300 экз./м2, в т.ч. нимф <100 экз./м2); 2-9 км (100% встречаемость орибатид в образцах почвы; плотность нимф до 300 экз./м2, средняя численность орибатид 1 тыс. экз./м2); 9-18 км (численность орибатид >10 тыс. экз./м2, нимф – до 800 экз./м2); от 18 км и далее (отсутствие проб, не заселенных нимфами; возрастание плотности орибатид до десятков тыс. экз./м2, нимф > 2 тыс. экз./м2, доли нимф – до 30%). На протяжении последней зоны существенных изменений в сообществе орибатид и отличий от контрольного участка не наблюдалось.
5.1.2. Панцирные клещи в почве импактной зоны. В техногенно трансформированных подзолах в 5 км от комбината «Североникель» (район 5) выявлено 12 видов орибатид: N. silvestris, С. borealis (= C. horridа), H. longisetosus, N. dorsalis (= N. coronata), E. oblongus, C. labyrinthicus, O. nova, S. palustris, C. traegardhi, E. occultus, Ch. borealis, S. laevigatus. Только в этом районе найден гемиэдафический вид Camisia borealis, типичный для березовых и сосновых лесов Карелии и побережья Белого моря (Ласкова, 2001; Каталог Биоты …, 2008) (см. табл. 2). Состав орибатид был наиболее сходен с таковым в подстилке зонального сосняка (район 8): 8 общих видов, Is = 38%. Это свидетельствовало о сохранении лесного комплекса орибатид в условиях длительного и сильного загрязнения.
Все виды орибатид относились к разным семействам, жизненным формам (к 4-м из 5 известных) и экологическим группам. Подобная обедненная структура выявлена также в зональной (район 1) и горной тундрах (район 6). Очевидно, что в экосистемах Кольского Севера с наиболее экстремальными природными условиями и зонах сильного техногенного воздействия формирование комплексов панцирных клещей подчиняется общей закономерности: сокращению числа семейств и их обеднению до одного рода с единственным видом. При этом даже при совпадении семейств, представленных в экстремальных природных и техногенно нарушенных экосистемах, набор родов и видов орибатид специфичен.
5.1.3. Комплексы микроартропод на площадках по восстановлению техногенной пустоши. Оценена эффективность восстановления техногенной пустоши в 1-км зоне воздействия комбината «Североникель» по состоянию комплексов микроартропод на 6 площадках с различным типом субстрата и способом восстановления – инвестиционный, ИП или экологический, ЭП подход (Исаева и др., 2010). Численность микроартропод при обоих подходах была выше на минеральном субстрате, что могло быть связано как с его механическими свойствами, так и меньшей токсичностью (рис. 8). Органические субстраты – слой отмершего сфагнума и насыпной слой торфа аккумулировали высокие дозы тяжелых металлов – Ni, Cu, Pb, Al, Co.
 | Рис. 8. Численность микроартропод на площадках. Субстраты: 3м, 4м, 5м – минеральный грунт, 2м – осадки сточных вод, 3о – торф, 5о – отмерший сфагнум. |
Площадки с ИП (2м, 3м) характеризовались более равномерным распределением микроартропод из-за меньшей пространственной гетерогенности почвенных факторов в результате планирования территории пустоши. В ходе фитомелиорации здесь сформировалась растительность лугового типа и «луговой» комплекс микроартропод с преобладанием коллембол и обилием мелких сапрофильных клещей. Это позволяет сделать предположение о более высокой микробиологической активности минеральных субстратов, сформированных в результате применения ИП.
Для панцирных клещей имел значение и способ восстановления, и тип субстрата. При обоих подходах их численность была в 2-12 раз выше на минеральных субстратах по сравнению с органическими (достоверно для большинства сравниваемых пар площадок) при максимальных значениях на пл. 5м, ЭП. Доля орибатид составляла 8-14% от всех микроартропод в минеральных субстратах против 3-5% в органических. Характеристика комплексов панцирных клещей, свидетельствующая в пользу ЭП, приведена в таблице 3.
Таблица 3. Показатели комплексов орибатид на площадках по восстановлению
| Под-ход | Пл. | Встреча-емость, % проб | Плотность, тыс. экз./м2 | Варьирование, CV, % | Агрегиро- вание по Лексису | Доля нимф, % | ||
| M ± m | min - max | Простран- ственное, n = 24 | Сезонное, n = 3 | |||||
| ИП | 2м | 74 | 0.45 ± 0.14 | 0 – 2.7 | 153 | 72 | 23 | 9.8 |
| 3м | 83 | 0.62 ± 0.18 | 0 – 3.4 | 137 | 107 | 29 | 1.1 | |
| 3о | 39 | 0.15 ± 0.09 | 0 – 2.04 | 282 | 110 | 15 | 0 | |
| ЭП | 4м | 65 | 0.34 ± 0.15 | 0 – 3.4 | 213 | 138 | 28 | 6.3 |
| 5м | 43 | 1.31 ± 0.86 | 0 – 18.1 | 316 | 99 | 470 | 0.8 | |
| 5о | 48 | 0.11 ± 0.04 | 0 – 0.7 | 160 | 61 | 3 | 0.9 | |
Сделан вывод о положительном влиянии всех подходов на состояние комплексов микроартропод по сравнению с исходным загрязненным субстратом, и, одновременно, об их низкой эффективности в связи со значительной техногенной трансформацией почвенного покрова и накоплением высоких доз поллютантов в условиях постоянной эмиссии комбината. При любом способе восстановления комплексы микроартропод техногенной пустоши характеризуются низким разнообразием и значительными пространственными и сезонными флуктуациям структурных параметров. В большинстве исследованных субстратов численность орибатид соответствовала ее уровню не далее чем в 2 км вдоль градиента загрязнения от комбината (см. рис. 6).
5.2. Влияние алюминиевого завода
5.2.1. Панцирные клещи в лесной подстилке. Воздействие фтор-содержащих выбросов алюминиевого завода на почвенную фауну менее губительно по сравнению с выбросами медно-никелевого комбината. В подстилке сосняка вороничного в 2 км от КАЗ (район 9) обнаружено 26 видов орибатид из 16 сем., что сопоставимо с контрольным зональным сосняком (район 8). Общими для двух сосняков оказались 12 видов из 8 сем. (Is = 43%), в т.ч. 6 видов микофагов-суктобельбид: S. acutidens, S. falcata, S. forsslundi, S. hammeri, S. singularis, S. subtrigona.
По сравнению с контрольным сосняком выявлен ряд изменений: 1 – исчезновение 17 видов (10 массовых в зональном сосняке: S. confundatus, C. forsslundi, C. coriaceus, C. reticulatus, C. marginatus, S. palustris, S. falcata, N. coronata, E. silvestris, H. longisetosus и 7 видов малочисленных: Belba sp., A. poppei, E. monodactylus, Ph. nitens, Ph. piger, P. nervosa, C. quadridentata); 2 – появление 12 новых видов: M. globosa, S. singularis, S. perforata, E. edwardsi, O. tibialis, P. leontonychus, M. minutissima, S. striculus, S. perforatus, N. humicola, P. dorsalis, M. testacea; 3 – достоверное сокращение численности 6 видов, общих для двух районов, в т.ч. 5 видов, доминирующих в подстилке зонального сосняка: Ch. borealis, S. subtrigona, C. subarcticus, S. forsslundi, Epidamaeus sp., T. velatus; 4 – достоверное возрастание (в 3 раза) численности эуэдафического скважника O. nova и эпиэдафического E. occultus; 5 – отсутствие достоверных различий между районами в плотности 4 мелких эуэдафических видов: M. unicarinata, S. hammeri, S. acutidens, S. forsslundi; 6 – увеличение доли моновидовых семейств с 63% (10 сем.) до 75% (12 сем.); 7 – смена крупных и среднеразмерных обитателей поверхности почвы и толщи подстилки мелкими почвенными скважниками, что типично для загрязненных почв.
В зоне загрязнения, как и в зональном сосняке, по числу видов преобладали сем. Suctobelbidae (8 видов), Oppiidae (4 видов) и Carabodidae (3 видов). Разнообразие мелких эуэдафических оппиид возросло (появились M. globosa, M. minus, M. neerlandica), крупных ксерофильных карабодид обеднилось (исчезли C. forsslundi, C. coriaceus, C. reticulates, C. labyrinthicus и появился C. areolatus), сменился состав мелких эуэдафических микофагов-суктобельбид (исчезли S. alloenasuta, S. baloghi, S. palustris, появились S. latirostris, S. perforatа), что, возможно, отражало изменения в структуре микромицетов в зоне загрязнения: угнетение видов с темнопигментированным, меланинсодержащим мицелием, снижение общей численности и биомассы грибов, смену видов в составе доминирующего рода Penicillium (Евдокимова и др., 2004, 2007).
Наиболее существенно (в 110 раз) сократилась численность T. velatus – основного доминанта в подстилке зонального сосняка. В результате описанных изменений при сохранении полидоминантной структуры (индекс Шеннона 3.8, индекс Пиелоу 0.76, индекс Симпсона 0.13), включающей трех доминантов с долей численности 8-9% и 6 субдоминирующих видов суктобельбид со степенью доминирования 4-7%, комплекс орибатид в 2 км от КАЗ отличался супердоминированием вида O. nova – 30% (рис. 9).
 | Рис. 9. Структура доминирования панцирных клещей в подстилке сосняка вороничного в 2 км от КАЗ. |
Преобладание этого эврибионтного, эуэдафического, партеногенетического вида типично для антропогенно нарушенных ландшафтов: самозарастающих отвалов горных карьеров, нефтезагрязненных почв, пахотных угодий, участков бывших торфоразработок, а также вулканогенных территорий (Чистяков, 1974; Артемьева, 1984; Рябинин, Паньков, 1986, 1987, 2005, 2009; Рябинин, 2003; Эйтминавичюте, Матусявичюте, 2009).
В образцах с растительными остатками, заложенными в загрязненную подстилку, обнаружено 35 видов, с учетом которых в окрестностях КАЗ обитает 42 вида орибатид из 32 родов, 22 сем. Сем. Parhypochthoniidae и 7 видов орибатид указаны для области впервые (см. табл. 2).
5.3. Панцирные клещи – индикаторы промышленного загрязнения почв. На основании встречаемости панцирных клещей в зонах воздействия промышленных предприятий и изменения их численности по сравнению с фоновыми территориями выделены категории видов, устойчивых и чувствительных к промышленному загрязнению почв Мурманской области (рис. 10). В категорию устойчивых включены виды, устойчивые к воздействию выбросов: I – алюминиевого завода, II – медно-никелевого комбината, III – промпредприятий разного типа (населяют техногенно трансформированные почвы в импактных зонах и КАЗ и «Североникеля»). Виды I категории: 1 – массовые в фоновых районах и недостоверно изменяющие численность под КАЗ; 2 – виды со стабильно низкой численностью как в 2 км от КАЗ, так и в других районах области; 3 – положительные индикаторы, найденные только в зоне действия КАЗ и достигающие здесь высокой плотности. Последние могут быть индикаторами повышения зольности (до 60%) и понижения кислотности (до pHводн 5.3-5.5) подстилки в результате ее загрязнения фтор-содержащими выбросами. Виды I-III категорий положительно коррелировали с численностью и биомассой микроорганизмов и величиной pH, отрицательно – с содержанием углерода и соотношением C:N в подстилках.
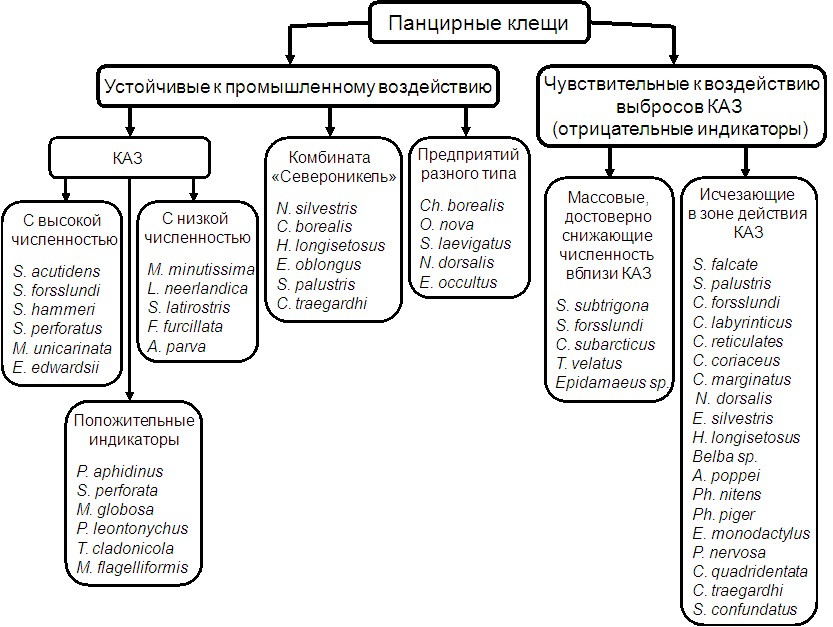
Рис. 10. Панцирные клещи, устойчивые и чувствительные к загрязнению почв
Мурманской области выбросами промышленных предприятий.
Виды, массовые в регионе, но снижающие плотность и/или исчезающие в импактных зонах предприятий, считаются отрицательными индикаторами промышленного загрязнения почв. К ним относятся виды наиболее разнообразных в регионе сем. Suctobelbidae и Carabodidae. Показательный отрицательный индикатор - T. velatus. Этот космополитный, партеногенетический, гемиэдафический вид является самым эвритопным среди клещей. Он встречен в 37 биотопах (или 86%) из 43 исследованных и преобладал по численности в зональных тундрах (районы 1, 2) и лесных подстилках (район 8) со степенью доминирования от 15 до 65%. Вблизи КАЗ вид резко снизил плотность, а в импактной зоне комбината «Североникель» выпал из сообщества орибатид. Причиной исчезновения T. velatus может быть его приуроченность к напочвенным лишайникам, которые угнетены в зоне воздействия алюминиевого завода и отсутствуют в 0-7 км от медно-никелевого предприятия.
5.4. Сравнение фаунистических комплексов орибатид природных и техногенно нарушенных почв. Видовой состав орибатид вблизи промпредприятий (районы 5, 9) оказался наиболее сходным с зональным сосняком в 50 км от источников эмиссии (район 8). Для сосняка в зоне воздействия КАЗ сходство с сосняком, произрастающим в центральной части области, было выше, чем с менее удаленными сосновыми и еловыми лесами на о-вах Кандалакшского заповедника. Из 12 видов, общих для двух сосняков, 10 видов (S. falcata, S. forsslundi, S. hammerae, S. acutidens, S. singularis, S. subtrigona, M. unicarinata, E. occultus, N. dorsalis, P. dorsalis) не выявлены в других районах области. В результате на дендрограмме фаунистического сходства эти сосняки образовали самостоятельный кластер (рис. 11).

Рис. 11. Дендрограмма сходства природных и техногенных биотопов по видовому составу орибатид (Ward`s method, Euclidean distances). БО – биотопы Беломорских островов.
В импактной зоне медно-никелевого предприятия, функционирующего в таежной зоне области, комплекс орибатид с крайне обедненной таксономической структурой был ближе к зональным и горным тундрам. Это свидетельствует о большем стрессирующем воздействии фактора техногенной трансформации почв на разнообразие орибатид по сравнению с экстремальными природно-климатическими условиями Кольского Севера.
6. ПАНЦИРНЫЕ КЛЕЩИ ТЕХНОГЕННЫХ СУБСТРАТОВ
(НА ПРИМЕРЕ ОТХОДОВ АПАТИТ-НЕФЕЛИНОВОЙ ПРОМЫШЛЕННОСТИ)
Обсуждаются результаты исследования пионерных фаунистических комплексов, которые сформировались в самозарастающих и рекультивированных нефелинсодержащих песках – отходах обогащения апатит-нефелиновых руд, складированных на хранилищах 0-40 лет назад и представляющих техногенный субстрат, отличный от зональных почв.
Первопоселенцами техногенного субстрата являются микробофаги-коллемболы, что объясняется наличием жизнеспособных бактериальных клеток в минеральных отходах обогащения апатит-нефелиновых руд (Евдокимова, Калмыкова, 2010). В отвалах 10-30 летнего срока складирования доля орибатид варьировала от 4% (100-200 экз./м2) в песках чистых и самозарастающих мхами до 20-35% (5.5 тыс. экз./м2) в ризосфере злаков, при посеве которых (для закрепления песков) вносили торф.
Формирование органогенного горизонта и растительного покрова в результате рекультивации и с увеличением срока складирования способствовало повышению микробиологической и ферментативной активности в песках (Евдокимова, Калмыкова, 2010), росту плотности панцирных клещей, их более равномерному пространственному распределению (особенно под мхами и лишайниками с более стабильным режимом увлажнения), смене жизненных форм, различиям в сроках и интенсивности репродукции. В песках, закрепленных > 40 лет назад, в составе микробиоты по биомассе преобладали микромицеты, а доминантами фауны стали панцирные клещи, связанные с грибами трофически. Численность клещей была сопоставима с уровнем зональных подзолов: 16-37 тыс. экз./м2 (рис. 12). Различия в численности орибатид в песках, закрепленных 10-30 лет назад и рекультивированных > 40 лет назад, составляли в разные месяцы вегетационного сезона от 9 до 110 раз (достоверно). Достоверного роста плотности коллембол с увеличением срока складирования песков не выявлено.

Рис. 12. Численность коллембол и орибатид в песках разного срока складирования. 1 – чистые пески, 2 – злаки, 3 – мхи, 4 – лишайники, 5 – вороника, 6 – брусника, 7 – клевер.
Установлена высокая корреляция плотности орибатид с ферментативной и микробиологической активностью в песках разного срока складирования (с биотическими факторами) и менее сильная – с их физико-химическими показателями (абиотическими факторами). Положительные корреляции с численностью микромицетов (r = 0.92), сапротрофных бактерий (0.77), активностью каталазы, фосфатазы, уреазы, инвертазы (r 0.83-0.92) и влажностью песков (0.50) отражали увеличение микробиологической и ферментативной активности и повышение влагоемкости под растительностью в «старых» отходах по сравнению с песками меньшего срока складирования. Отрицательные корреляции с температурой и pH водной суспензии объяснялись слабым прогревом песков под растительностью и снижением значений pH исходно щелочного субстрата до 6.7 под воздействием гумусовых кислот и продуктов жизнедеятельности биоты. Таким образом, в процессе первичного почвообразования на техногенном субстрате – отходах нефелинсодержащих пород панцирные клещи проявили себя как индикаторы микологической активности субстрата и накопления органического вещества.
7. ОБОБЩЕННАЯ ХАРАКТЕРИСТИКА
ФАУНЫ ПАНЦИРНЫХ КЛЕЩЕЙ МУРМАНСКОЙ ОБЛАСТИ
Обобщены данные по экологическому разнообразию (тип ареала, трофическая специализация, жизненные формы) для 259 видов панцирных клещей, выявленных на территории Мурманской области. Рассмотрено влияние фактора широтной зональности.
Показано преобладание видов с голарктическими (52% видов) и палеарктическими (38%) ареалами, как и на Европейском Севере России в целом (Мелехина, 2009). Малой мощности подстилки и укороченному профилю зональных почв соответствует высокая доля поверхностно-обитающих видов (50%), 30% приходится на обитателей почвенных скважин, 20% - на обитателей толщи подстилки.
Основу региональной фауны формирует эволюционно более молодая и прогрессивная группа высших панцирных клещей (Brachypylina), разнообразие низших орибатид (Macropylina) примерно в 2,5 раза ниже на уровне видов и надвидовых таксонов – родов и семейств (табл. 4).
Таблица 4. Соотношение высших (Brachypylina) и низших (Macropylina) орибатид
в таежных и тундровых почвах Мурманской области
| Орибатиды | Область в целом | Северная тайга | Зона тундры | ||||||
| Сем | Род | Вид | Сем | Род | Вид | Сем | Род | Вид | |
| Всего, в т.ч.: | 53 | 113 | 258 | 47 | 102 | 216 | 38 | 78 | 142 |
| Brachypylina | 39 | 83 | 182 | 33 | 74 | 154 | 29 | 60 | 101 |
| Macropylina | 14 | 30 | 76 | 14 | 28 | 62 | 9 | 18 | 41 |
| Соотношение, разы | 2,8 | 2,6 | 2,4 | 2,4 | 2,6 | 2,5 | 3,2 | 3,3 | 2,5 |
Треть семейств (34%) представлена единственным видом, 26% включают по 2-3 вида. Многовидовые таксоны Brachychthoniidae, Oppiidae, Suctobelbidae, Damaeidae, Ceratozetidae с числом видов от 15 до 18 составляют лишь 9%, объединяя при этом 30% видов. Клещи трех первых семейств – мелкоразмерные почвенные скважники, микрофитофагами (потребители грибов, бактерий, водорослей). Виды сем. Damaeidae и Ceratozetidae обитают на поверхности почвы и в толще подстилки. Видовая насыщенность семейств орибатид в исследованных районах приведена в табл. 5.
| Таблица 5. Видовая насыщенность семейств орибатид в исследованных районах | |||||||||||
| № | Семейство / Район | 1 | 2 | 3 | 5 | 6 | 8 | 9 | 10 | 11 | |
| Низшие орибатиды (Macropylina) | 1 | Paleacaridae (Grandjean 1932) | 2 | 1 | |||||||
| 2 | Brachychthoniidae (Thor 1934) | 10 | 5 | 14 | |||||||
| 3 | Hypochthoniidae (Berlese 1910) | 1 | 1 | 1 | 1 | ||||||
| 4 | Eniochthoniidae (Grandjean 1947) | 1 | 1 | ||||||||
| 5 | Parhypochthoniidae (Grandjean 1932) | 1 | |||||||||
| 6 | Eulohmanniidae (Grandjean 1931) | 1 | 1 | ||||||||
| 7 | Phthiracaridae (Perty 1841) | 1 | 3 | 3 | 1 | 3 | 1 | 3 | 7 | ||
| 8 | Euphthiracaridae (Jacot 1930) | 1 | 2 | 1 | 2 | 2 | 6 | ||||
| 9 | Malaconothridae (Berlese 1916) | 2 | 2 | ||||||||
| 10 | Trhypochthoniidae (Willmann 1931) | 2 | 1 | 1 | 1 | 1 | |||||
| 11 | Nothridae (Berlese 1896) | 1 | 3 | 1 | 1 | 3 | 6 | ||||
| 12 | Camisiidae (Oudemans 1900) | 2 | 7 | 2 | 6 | 2 | 2 | 8 | 10 | ||
| 13 | Nanhermanniidae (Sellnick 1928) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | |
| 14 | Hermanniidae (Sellnick 1928) | 1 | 1 | 3 | 1 | 3 | |||||
| Высшие орибатиды (Brachypylina) | 15 | Hermanniellidae (Grandjean 1934) | 1 | ||||||||
| 16 | Licheremaeidae (Grandjean 1931) | 1 | |||||||||
| 17 | Damaeidae (Berlese 1896) | 1 | 9 | 3 | 2 | 2 | 3 | 11 | |||
| 18 | Cepheidae (Berlese 1896) | 1 | 1 | 1 | |||||||
| 19 | Eremaeidae (Sellnick 1928) | 1 | 1 | 1 | 1 | 2 | |||||
| 20 | Tenuialidae (Jacot 1929) | 1 | |||||||||
| 21 | Astegistidae (Balogh 1961) | 1 | 1 | 2 | 4 | ||||||
| 22 | Gustaviidae (Oudemans 1900) | 1 | |||||||||
| 23 | Liacaridae (Sellnick 1928) | 2 | 3 | 1 | 2 | 2 | 6 | 8 | |||
| 24 | Peloppiidae (Balogh 1943) | 2 | 1 | 1 | 1 | 3 | |||||
| 25 | Carabodidae (C.L. Koch 1837) | 3 | 3 | 5 | 1 | 4 | 6 | 3 | 3 | 9 | |
| 26 | Tectocepheidae (Grandjean 1954) | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 3 | ||
| 27 | Quadroppiidae (Balogh 1983) | 1 | 1 | ||||||||
| 28 | Caleremaeidae (Grandjean 1965) | 1 | |||||||||
| 29 | Oppiidae (Grandjean 1954) | 1 | 2 | 10 | 1 | 3 | 2 | 5 | 12 | 12 | |
| 30 | Suctobelbidae (Jacot 1938) | 1 | 1 | 6 | 1 | 9 | 7 | 1 | 8 | ||
| 31 | Autognetidae (Grandjean 1960) | 1 | 1 | 1 | 1 | 2 | 4 | 3 | |||
| 32 | Thyrisomidae (Grandjean 1954) | 2 | 3 | 3 | |||||||
| 33 | Hydrozetidae (Grandjean 1954) | 1 | |||||||||
| 34 | Limnozetidae (Grandjean 1954) | 3 | 2 | ||||||||
| 35 | Ameronothridae (Willmann 1931) | 4 | 6 | 6 | |||||||
| 36 | Cymbaeremaeidae (Sellnick 1928) | 1 | 1 | ||||||||
| 37 | Micreremidae (Grandjean 1954) | 2 | |||||||||
| 38 | Passalozetidae (Grandjean 1954) | 1 | |||||||||
| 39 | Scutoverticidae (Grandjean 1954) | 1 | 1 | 2 | 2 | ||||||
| 40 | Haplozetidae (Grandjean 1954) | 1 | |||||||||
| 41 | Phenopelopidae (Petrunkevitch 1955) | 1 | 5 | 1 | 1 | 1 | 1 | 4 | |||
| 42 | Achipteriidae (Thor 1929) | 1 | 1 | 1 | 1 | 2 | 2 | ||||
| 43 | Tegoribatidae (Grandjean 1954) | 1 | 1 | ||||||||
| 44 | Oribatellidae (Jacot 1925) | 1 | 3 | ||||||||
| 45 | Galumnidae (Jacot 1925) | 4 | 1 | 1 | 1 | 5 | 5 | ||||
| 46 | Ceratozetidae (Jacot 1925) | 2 | 2 | 10 | 5 | 1 | 8 | 11 | |||
| 47 | Chamobatidae (Thor 1938) | 1 | 1 | 4 | 1 | 1 | 1 | 1 | 2 | 2 | |
| 48 | Humerobatidae (Grandjean 1970) | 1 | |||||||||
| 49 | Mycobatidae (Grandjean 1954 | 1 | 1 | 1 | |||||||
| 50 | Euzetidae (Grandjean 1954) | 1 | |||||||||
| 51 | Parakalummidae (Grandjean 1936) | 2 | |||||||||
| 52 | Scheloribatidae (Grandjean 1953) | 1 | 2 | 5 | 1 | 3 | 1 | 3 | 5 | 5 | |
| 53 | Oribatulidae (Thor 1929) | 1 | 2 | 2 | 1 | 1 | 2 | 3 | |||
| Видов в каждом районе | 14 | 27 | 123 | 12 | 42 | 38 | 41 | 113 | 178 | ||
Многовидовые семейства формируют основу фауны орибатид и в северотаежных, и в тундровых почвах области, однако для 9 из них в тундре число видов на 4-6 меньше, чем в тайге (рис. 13). Общими для Кольской тундры и тайги являются 48 видов (< 20% региональной фауны) из 23 сем. Степень фаунистического сходства выше на уровне семейств (Is = 38%), чем на видовом уровне (68%), что отражает большую специализацию видов к условиям среды по сравнению с таксонами высших рангов.

Рис. 13. Насыщенность семейств орибатид видами (I) и родами (II) в северотаежных и тундровых почвах Мурманской области. Обозначение семейств с 1 по 53 – как в табл. 5.
Сравнение списков орибатид северной тайги, тундры Мурманской области и граничащей с юга республики Карелия (северная и средняя тайга, 61-66° с.ш.) выявило тенденцию уменьшения числа одно- и многовидовых семейств с увеличением широты местности (табл. 6). В области отсутствуют 7 семейств, представленных в Карелии единственным видом, но есть 7 моновидовых таксонов, не указанных для республики.
Таблица 6. Соотношение семейств орибатид с разным числом видов
в почвах Мурманской области и республики Карелия
| Семейства | Карелия, средняя тайга (Ласкова, 2001) | Мурманская область | ||||||
| Подзона северной тайги | Зона тундры | Область в целом | ||||||
| Сем. | Доля, % | Сем. | Доля, % | Сем. | Доля, % | Сем. | Доля, % | |
| Одновидовые | 18 | 37 | 13 | 28 | 11 | 31 | 18 | 34 |
| 2-3 видовые | 14 | 29 | 17 | 36 | 9 | 25 | 14 | 26 |
| 4 видов | 17 | 35 | 17 | 36 | 15 | 42 | 21 | 40 |
| Всего сем. | 49 | 100 | 47 | 100 | 36 | 100 | 53 | 100 |
Высокая доля одновидовых семейств орибатид в фауне северных регионов отражает миграционный характер ее формирования. Для Мурманской области распространение клещей возможно как минимум тремя путями: с морскими течениями (Нордкапская ветвь Гольфстрима), антропогенным привносом, путем форезии на мигрирующих птицах. Это подтверждается специфичностью видового состава и высокой долей локально встречающихся видов на морских побережьях. Из 37% локально встречающихся видов региональной фауны 24% (или 63 вида) найдены на побережье Баренцевого моря, 7% (17 видов) – на о-вах Белого моря, тогда как в горном районе – лишь 2% или 6 видов. Степень видового сходства выше для морских побережий (Is = 41%), чем для побережий и центральных районов (Is = 33-38%). Без учета локально встречающихся видов фауна орибатид региона насчитывает 162 вида.
«Прогнозное» разнообразие орибатид Мурманской области, рассчитанное по 11 исследованным районам, оценивается в 290 видов, что сопоставимо с разнообразием этих клещей для стран Северной Фенноскандии и превышает известное число видов для республик Карелия и Коми (Lundqvist, 1987; Schatz, 2004; Ласкова, 2001; Мелехина, 2004).
ВЫВОДЫ
- Фауна естественных (равнинные и горные) и техногенно трансформированных северотаежных экосистем Мурманской области включает 216 видов панцирных клещей, принадлежащих к 102 родам, 47 сем. Список орибатид Кольской тундры расширен до 142 видов из 78 родов, 38 сем., общий региональный список – до 259 видов из 113 родов, 53 сем. В зоогеографическом отношении преобладают виды с голарктическими (52%) и палеарктическими (38%) ареалами; 50% видов представлены поверхностно обитающими формами; 70% видов орибатид являются панфитофагами.
- Многовидовые таксоны Brachychthoniidae, Oppiidae, Suctobelbidae, Damaeidae, Ceratozetidae с числом видов от 15 до 18 объединяют треть видов региональной фауны. Высокая доля одновидовых семейств (34%) отражает миграционный характер формирования фауны орибатид Мурманской области, причем разными путями: форезией клещей на птицах, распространением с морскими течениями, антропогенным привносом.
- Влияние широтной зональности на разнообразие панцирных клещей в пределах Мурманской области выражается в сокращении числа видов, родов и семейств, главным образом за счет таксонов низших орибатид (Macropylina), и в снижении доли двух-, и трехвидовых семейств при переходе от таежной зоны к тундровой.
- В пределах таежного, субальпийского и альпийского высотных поясов Хибинского горного массива выявлено 49 видов орибатид из 37 родов, 23 сем. Семейство Haplozetidae и виды Protoribates capucinus, Heminothrus septentrionalis, Nanhermannia sellnicki обнаружены только в почвах Хибин. Установлено большее фаунистическое сходство горно-тундровых и горно-лесотундровых сообществ по сравнению с предгорными (горно-таежными). Предгорные леса на окраинах Хибинского массива, расположенного в северотаежной подзоне, ближе к зональным тундрам по сравнению с зональной тайгой.
- В естественных экосистемах Кольского Севера с наиболее экстремальными природными условиями (зональные и горные тундры) и зонах сильного техногенного воздействия (медно-никелевый комбинат) формирование комплексов орибатид подчиняется общей закономерности: сокращение числа семейств и их обеднение до одного рода с единственным видом. При совпадении семейств, представленных в экстремальных природных и техногенно нарушенных экосистемах, сообщества орибатид имеют своеобразный набор видов.
- Воздействие фтор-содержащих выбросов алюминиевого завода на почвенную фауну менее губительно по сравнению с выбросами медно-никелевого комбината. В зоне максимального загрязнения сохраняется высокое разнообразие орибатид, сопоставимое с зональной тайгой, и полидоминантная структура их сообщества. О негативном воздействии аэротехногенных выбросов на почву свидетельствует преобладание мелких эуэдафических видов из экологической группы почвенных скважников и супердоминирование эврибионтного партеногенетического вида O. nova.
- Выделены категории панцирных клещей, устойчивых и чувствительных к промышленному загрязнению почв Мурманской области; среди них 13 представителей сем. Suctobelbidae и Carabodidae, преобладающих по числу видов в региональной фауне. Для индикаторных видов в загрязненных подстилках значимыми факторами являются численность и биомасса микроорганизмов, уровень кислотности, содержание углерода и соотношение C:N в подстилках.
- Установлена сукцессия пионерных групп микроартропод, связанная со сменой бактериального и грибного компонентов микробиоты, в процессе первичного почвообразования на нефелинсодержащих отходах обогатительных фабрик. Увеличение срока складирования отходов сопровождается ростом численности орибатид, их более равномерным пространственным распределением, снижением доли мелкоразмерных обитателей почвенных скважин и возрастанием количества обитателей подстилки и поверхности почвы, что соответствуют перестройкам в сообществах панцирных клещей в ходе восстановительных сукцессий на нарушенных территориях.
Список публикаций по теме исследования
Публикации изданиях, рекомендованных ВАК
- Зенкова И.В., Калмыкова В.В., Лисковая А.А. Пионерная фауна отвалов нефелин-содержащих пород // Почвоведение, 2009. № 8. С. 1-8.
- Зенкова И.В., Зайцев А.С., Залиш Л.В., Лисковая А.А. Почвообитающие панцирные клещи (Acarina, Oribatei) таежной и тундровой зон Мурманской области // Зеленый пояс Фенноскандии / Труды Карельского научного центра РАН, 2011. № 1. Сер. Биогеография. Вып. 11. С. 54-67.
Публикации в тематических сборниках и трудах конференций
- Зенкова И.В., Лисковая А.А. Пионерная фауна нефелинсодержащих отвалов как показатель процессов первичного почвообразования // Экологические проблемы Северных регионов и пути их решения: Матер. Всерос. конф. Апатиты: Кольский НЦ РАН, 2008. С. 87-92.
- Зенкова И.В., Лисковая А.А. Сукцессия микроартропод в отвалах действующих и законсервированных хранилищ ОАО «Апатит» // Северные территории России: проблемы и перспективы развития: Матер. Всерос. конф. Архангельск: УрО РАН, 2008. С. 486-490.
- Лисковая А.А., Зенкова И.В. Степень изученности почвообитающих панцирных клещей (Acarina, Oribatei) на территории Мурманской области // Актуальные проблемы сохранения биоразнообразия в экстремальных условиях северного климата: Матер. межд. науч. конф. Кировск: ПАБСИ РАН, 2008. С. 52-55.
- Лисковая А.А., Зенкова И.В. Пионерная фауна нефелинсодержащих отвалов как показатель процессов первичного почвообразования // Почвенные сообщества: от структуры к функциям: Тез. XV Всерос. совещ. по почв. зоол. М.: ИПЭЭ, 2008. С. 284-287.
- Зенкова И.В., Лисковая А.А. Разнообразие панцирных клещей (Acariformes, Oribatei) в почвах Мурманской области // Сохранение биологического разнообразия наземных и морских экосистем в условиях высоких широт: Матер. межд. науч.-практ. конф. Мурманск: МПГУ, 2009. С. 96-101.
- Зенкова И.В., Лисковая А.А. Разнообразие панцирных клещей (Acariformes, Oribatei) в целинных и антропогенно нарушенных почвах Мурманской области // Продуктивность и устойчивость лесных почв: Матер. докладов III межд. конф. по лесному почвоведению. Петрозаводск: КарНЦ РАН, 2009. С. 199-203.
- Зенкова И.В., Лисковая А.А. Панцирные клещи (Acariformes, Oribatei) в таежных и тундровых почвах Мурманской области // Биогеография почв: Тез. II Всерос. конф. посвященной 70-летию со дня рождения Д.А. Криволуцкого. М.: МГУ, 2009. С. 34.
- Зенкова И.В., Лисковая А.А. Оценка эффективности способов восстановления техногенной пустоши в окрестностях медно-никелевого комбината «Североникель» по состоянию комплексов микроартропод // Современные проблемы загрязнения почв: Матер. III межд. науч. конф., М.: МГУ, 2010. С. 508-512.
- Зенкова И.В., Лисковая А.А. Почвообитающие беспозвоночные как индикаторы восстановления техногенной пустоши в окрестностях комбината «Североникель» (г. Мончегорск) // Сбалансированное природопользование: Матер. V Школы молодых ученых и специалистов КНЦ РАН. Апатиты: Изд-во КНЦ РАН, 2010. С. 41-47.