

Генопротективные эффекты мелатонина при химических и радиационных воздействиях (экспериментальное исследование)
На правах рукописи
ПИКАЛОВА
Лидия Васильевна
ГЕНОПРОТЕКТИВНЫЕ ЭФФЕКТЫ МЕЛАТОНИНА ПРИ
ХИМИЧЕСКИХ И РАДИАЦИОННЫХ ВОЗДЕЙСТВИЯХ
(Экспериментальное исследование)
14.03.04 – токсикология
03.01.01 – радиобиология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Санкт-Петербург
2012
Работа выполнена в ФГВОУ ВПО «Военно-медицинская академия имени С.М. Кирова» Министерства обороны Российской Федерации
Научные руководители:
доктор медицинских наук Иванов Максим Борисович
доктор медицинских наук, профессор Легеза Владимир Иванович
Официальные оппоненты:
доктор биологических наук Назаров Виктор Борисович
доктор биологических наук Чигарева Наталия Григорьевна
Ведущая организация:
ФГУП «Научно-исследовательский институт промышленной и морской медицины» ФМБА России
Защита диссертации состоится «24» января 2012 г. в _______ на заседании диссертационного совета Д 208.030.01 при ФГУН «ИНСТИТУТ ТОКСИКОЛОГИИ» ФМБА России (192019, Санкт-Петербург, ул. Бехтерева, 1).
С диссертацией можно ознакомиться в научно-медицинской библиотеке ФГУН «ИНСТИТУТ ТОКСИКОЛОГИИ» ФМБА России
Автореферат разослан «21» декабря 2011 г.
Ученый секретарь диссертационного совета
доктор медицинских наук, профессор
Луковникова Любовь Владимировна
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность. Одним из фундаментальных свойств живого организма является способность поддерживать стабильность генома. На молекулу дезоксирибонуклеиновой кислоты (ДНК) постоянно воздействуют различные факторы экзогенной (ультрафиолетовое и ионизирующее излучение, химические мутагены) и эндогенной природы (активные радикалы кислорода), способные нарушить ее структуру. В живой клетке существуют молекулярные механизмы, способные распознавать повреждения молекулы ДНК, устранять их или элиминировать генетически-нестабильные клетки путем индукции клеточной гибели (Бочков Н.П., 2008; Jackson S.P., Bartek J, 2009). Однако при определенной силе воздействия могут наблюдаться нарушения в системе ответа на повреждение ДНК и репарации, что способствует развитию различных генетически-ассоциированных патологических состояний, включая иммунодефициты, преждевременное старение и рак (Hoeijmakers J.H., 2001).
Сегодня практически невозможно представить существование человечества без использования химических веществ и источников ионизирующих излучений в различных областях промышленности, медицины, научных исследованиях и т.д. В связи с этим наблюдается неуклонный рост числа людей находящихся под прессингом повышенной ксенобиотической нагрузки. В результате формируются различные формы патологических процессов (токсические или радиационно-обусловленные), проявляющихся канцерогенезом, мутагенезом и тератогенезом (на уровне организма) и/или повышением заболеваемости по различным классам болезней (на уровне популяции). В связи с изложенным выше одним из актуальных направлений современной токсикологии и радиобиологии является поиск средств, способных снижать вредное воздействие токсикантов и ионизирующего излучения. Активно ведется поиск средств растительного и животного происхождения, оказывающих генопротективный эффект (Куценко С.А., 2002). По данным литературы, одним из таких соединений является эпифизарный гормон мелатонин (Анисимов В.Н., 2006; Anisimov V.N., Popovich I.G., 2006; Claustrat B. et al., 2005; Musatov S.A., 1998; Vijayalaxmi Ph.D., 2004).
Вместе с тем, многие вопросы, касающиеся выяснения конкретных особенностей генопротеторного действия мелатонина при воздействии различных кластогенных факторов химической и радиационной этиологии, а также механизмы проявления этого эффекта остаются малоизученными, что и обусловило актуальность проведения настоящей работы.
Цель: исследование влияния мелатонина на кластогенез химической и радиационной этиологии и уточнение механизмов его генопротективного действия.
Для достижения поставленной цели определены основные задачи исследования:
- В опытах на крысах изучить влияние мелатонина в различных дозах на кластогенные эффекты цитостатика (циклофосфамида) или -излучения;
- Исследовать влияние мелатонина на состояние процессов свободно-радикального окисления в крови и тканях крыс, подвергнутых воздействию циклофосфамида или -излучения;
- Изучить влияние мелатонина в различных концентрациях на индуцированное циклофосфамидом ингибирование роста культуры тканей селезенки;
- Исследовать действие мелатонина в различных концентрациях на митогенно-миграционную активность лейкоцитов периферической крови человека, ингибированную циклофосфамидом.
Основные положения, выносимые на защиту:
- Мелатонин способствует существенному уменьшению кластогенного действия токсикантов (цитостатиков), оказывает нормализующее действие на процессы перекисного окисления липидов, миграционную и пролиферативную активность иммунокомпетентных клеток и пролиферацию лимфоидной ткани;
- Мелатонин снижает повреждающее действие облучения на генетический аппарат клетки и уменьшает выраженность проявлений постлучевого оксидативного стресса.
Научная новизна. Впервые установлено, что при однократном профилактическом применении синтетический отечественный мелатонин обладает выраженным антимутагенным действием, защищая хромосомный аппарат клеток от повреждающего действия токсикантов (цитостатиков) и ионизирующего излучения.
Показано, что мелатонин существенно снижает выраженность проявлений оксидативного стресса при химических и радиационных воздействиях. Под влиянием препарата уменьшается количество эндотоксинов, образующихся в условиях усиления перекисного окисления липидов, и повышается активность системы антиоксидантной защиты.
Впервые установлено, что мелатонин способствует нормализации угнетаемой циклофосфамидом пролиферативной и митогенно-миграционной активности клеток периферической крови и пролиферации лимфоидной ткани.
Практическая значимость результатов работы. В ходе проведенных исследований разработаны оптимальные режимы (дозы и сроки) использования мелатонина в качестве средства защиты хромосомного аппарата от кластогенного действия цитостатических препаратов и ионизирующих излучений. Предложена комплексная система критериев оценки профилактического эффекта мелатонина в условиях воздействия на организм ксенобиотиков и ионизирующего излучения, включающая определение цитогенетических показателей (различных типов хромосомных аберраций в клетках костного мозга), пролиферативной и миграционной активности лейкоцитов периферической крови, а также пролиферацию лимфоидной ткани.
Апробация работы. Основные результаты диссертационного исследования доложены на Всероссийской научной конференции «Медицинские и биологические аспекты токсикологии и радиобиологии» (2008); II-м Санкт-Петербургском международном экологическом форуме (2008); I-й Международной студенческой конференции «Лицейские чтения» (2009); VI-м съезде Всероссийского общества медицинских генетиков (2010); Конференции молодых ученых (2010); VI-м съезде по радиационным исследованиям (радиобиология, радиоэкология, радиационная безопасность) (2010); Конференции «Актуальные проблемы медицины и биологии» (2010); XXXXIII научной конференции СПбМАПО «Хлопинские чтения» (2010); Всероссийской научно-практической конференции «Химическая безопасность Российской Федерации в современных условиях» (2010); Объединенном пленуме «Научно-методические и законодательные основы обеспечения генетической безопасности факторов и объектов окружающей и производственной среды в целях сохранения здоровья человека» (2010); Научно-практической конференции молодых ученых «Актуальные вопросы клинической и экспериментальной медицины» (2011); Российской научной конференции «Актуальные проблемы токсикологии и радиобиологии» (2011).
Реализация результатов работы. Полученные в ходе диссертационного исследования материалы внедрены в учебный процесс на профильных кафедрах Военно-медицинской академии им. С.М.Кирова, Северо-западного государственного медицинского университета им. И.И.Мечникова при рассмотрении вопросов, связанных с изучением хромосомных повреждений при действии факторов химической и радиационной природы, а также в научно-исследовательскую работу Санкт-Петербургского научно-исследовательского института скорой помощи имени И.И. Джанелизде и научно-производственного центра «Фармзащита» ФМБА России при разработке медицинских средств защиты.
Связь темы диссертации с плановой тематикой научно-исследовательской работы учреждения. Исследование выполнялось в соответствии с плановой тематикой научно-исследовательских работ НИЛ (военной терапии) НИО (экспериментальной медицины) НИЦ Военно-медицинской академии им. С.М. Кирова: VMA.03.12.07.0912/0286, шифр «Аберрация», VMA.02.02.01.0910/0043, шифр «Селен».
Личное участие автора. Автор принимал личное участие в планировании, организации и выполнении экспериментальных исследований, проводил учет и оценку их результатов, статистическую обработку, обобщение и анализ полученных данных.
Публикации. По теме диссертации опубликовано 19 научных работ, в том числе 2 публикации в центральных медицинских изданиях, входящих в перечень ВАК, 4 в электронных научных изданиях, зарегистрированных в ФГУП НТЦ «Информрегистр», оформлено 4 рационализаторских предложения: № 11772/7; 11773/7; 11774/7; 11775/7 от 06.11.2009 г.
Объем и структура диссертации. Диссертация состоит из введения, обзора литературы и собственных данных, включающих описание материалов и методов, результатов исследования, их обсуждения, выводов, практических рекомендаций, списка цитируемой литературы. Объем работы – 111 страниц машинописного текста. Иллюстративный материал включает 11 таблиц и 9 рисунков. Список литературы включает 241 источник: 99 отечественных и 142 зарубежных авторов.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Для достижения цели и решения поставленных задач проведены экспериментальные исследования, включавшие опыты in vivo, ex vivo и in vitro с использованием токсикологических, цитогенетических, биохимических и иммунологических методов.
Анализ частоты хромосомных аберраций в миелокариоцитах костного мозга и биохимические исследования выполнены на 620 белых беспородных трехмесячных крысах-самцах массой 180–270 г. Для изучения влияния мелатонина и циклофосфана на рост клеток в культуре ткани использовали 800 фрагментов селезенки трехмесячных крыс-самцов линии Вистар. Животных приобретали в питомнике «Рапполово» РАМН. Содержание и использование животных в эксперименте проводили согласно положениям, изложенным в работе И.П.Западнюка (1983).
Оценка реакций миграционной и пролиферативной активности выполнена на периферической венозной крови 55 мужчин. Состояние здоровья добровольцев оценивали по данным анамнеза (отсутствие острых и хронических заболеваний), субъективным показателям (отсутствию жалоб на состояние здоровья), в ходе физикально-клинического (термометрия, показатели артериального давления, частота дыхания и др.) исследования.
В работе использовали мелатонин (НПЦ «Фармзащита ФМБА России»). Для профилактики химически- и радиационно-индуцированного мутагенеза препарат применяли в дозах 5, 10, 25 и 50 мг/кг. В течение суток после введения мелатонина у крыс не наблюдалось изменений физического состояния и поведенческих реакций. В биохимических исследованиях препарат использовали в дозе 50 мг/кг. В иммунологических и культуральных исследованиях мелатонин вводили в среду в концентрациях 0,01 нг/мл – 1 мкг/мл.
В качестве токсиканта применяли импортный аналог циклофосфана – эндоксан (циклофосфамид) фирмы Baxter (Германия). ЛД50 для крыс по данным наших исследований составила 250 мг/кг. Для моделирования мутагенеза у крыс использовали дозу 200 мг/кг. В иммунологических и культуральных исследованиях применяли циклофосфамид в концентрациях 0,05 – 200 мкг/мл.
Препараты разводили 0,9% раствором натрия хлорида и вводили внутрибрюшинно в объеме 0,1 мл на 100 г массы тела стерильным стандартным шприцом с иглой длиной 20 мм. Введение фармакологических средств животным проводили в утренние часы. Через сутки животных выводили из опыта методом цервикальной дислокации или декапитации. В экспериментах in vitro препараты добавляли в культуральную среду.
В качестве источника ионизирующего излучения использовали установку для облучения животных «Игур-1» с изотопом 137Cs; кожно-фокусное расстояние 1 м; облучение двустороннее боковое. Животных подвергали внешнему острому однократному общему воздействию -облучения в дозах 2 или 4 Гр, мощность дозы – 1 Гр/мин. Дозиметрический контроль условий облучения осуществляли расчетным методом. В ходе каждого облучения животных проводилась физическая дозиметрия с помощью индивидуального дозиметра ИД-11.
Динамику формирования генетических повреждений в миелокариоцитах костного мозга при воздействии циклофосфамида, -облучения и мелатонина изучали с помощью метафазного метода. Приготовление цитогенетических препаратов осуществляли стандартным суховоздушным способом (Moorhead P.S. et al., 1960; Preston R.J. et al., 1987). Цитогенетический анализ проводили на основе рекомендаций D. Scott et al. (1983) с помощью светового микроскопа "Jenaval" 10х100. В каждом препарате анализировали по 100 метафазных пластинок, учитывая одиночные и парные фрагменты хромосом, дицентрические хромосомы, транслокации, а также полиплоидные клетки.
Органотипическое культивирование тканей проводили по методике Н.И. Чалисовой и соавт. (2003). Выделенные фрагменты селезенки (1 мм3) помещали в стерильные чашки Петри с коллагеновым покрытием и культивировали в течение 3 сут в 3 мл питательной среды (раствор Хенкса 41 мл, среда Игла 30 мл, фетальная телячья сыворотка 25 мл, глюкоза 40% - 1 мл, инсулин 2,5 мл (100 ЕД) и гентамицин 0,5 мл (20 мг) на 100 мл среды) в СО2-инкубаторе при температуре 36,7оС с 5% содержанием СО2. В качестве модельных веществ в питательную среду добавляли мелатонин в концентрациях 0,01; 0,05; 1,0; 10,0 и 100 нг/мл; циклофосфамид в концентрациях 0,01; 0,05; 0,5; 1 и 10 мкг/мл; циклофосфамид в концентрации 1 мкг/мл и мелатонин в концентрациях 0,01; 0,05; 1,0; 10,0 или 100 нг/мл. В контрольных образцах в питательную среду препараты не добавляли. Визуализацию осуществляли с помощью микротеленасадки к микроскопу (серия 10, МТН-13 «Альфа-Телеком», Россия). Для расчета индекса площади эксплантатов использовали программу PhotoM 1.2, позволяющую определить отношение общей площади эксплантата к площади центральной зоны.
При исследовании реакции торможения миграции лейкоцитов использовали образцы периферической крови человека. Учет результатов проводили под световым микроскопом (объектив х10), определяя длину зоны миграции основной массы лейкоцитов от границы эритроцитарного осадка в опыте и контроле. Результаты выражали в виде процента миграции:
Р = Lm L x 100, где Р – процент миграции; Lm – длина зоны миграции в присутствии митогена; L – длина зоны миграции в контроле.
Величину общей антиоксидантной (ОАА) и общей оксидантной (ООА) активности плазмы крови определяли по методу Л.П. Галактионовой (1998).
Результаты ОАА рассчитывали по формуле:
ОАА = (ОП – К) К 100, где ОАА – общая антиоксидантная активность (%); ОП – оптическая плотность опытной пробы; К – оптическая плотность контрольной пробы.
Результаты ООА рассчитывали по формуле:
ООА = (ОП – К) ОП 100, где ООА – общая оксидантная активность (%); ОП – оптическая плотность опытной пробы; К – оптическая плотность контрольной пробы.
Определение индуцированной биохемилюминисценции в плазме крови проводили с помощью биохемилюминометра БХЛ-06 (Россия), используя модельную систему – 2% раствор перекиси водорода и 0,05 mM раствор сульфата железа. Регистрировали следующие параметры сигнала: S – общая светосумма сигнала, mVc; Imax – значение максимальной интенсивности сигнала, mV; STmax – светосумма сигнала до момента достижения его максимальной интенсивности, mVc. Состояние процессов свободнорадикального окисления (СРО) оценивали по показателям Imax, а активность антиоксидантной системы (АОС) - по соотношению общей светосуммы сигнала к светосумме сигнала до момента достижения его максимальной интенсивности (S/STmax).
Концентрацию малонового диальдегида (МДА) рассчитывали с использованием методики В.Б. Гаврилова и соавт. (1976) по разнице оптических плотностей Е535 – 580 с учетом разведения и коэффициента пересчета К=1,88·105 М-1·см-1 и выражали в нмоль/г ткани (для гомогенатов тканей) и нмоль/г гемоглобина (для гемолизата эритроцитов).
Определение концентрации диеновых конъюгат (ДК) осуществляли по методике И.Д. Стальной (1977). Концентрацию ДК рассчитывали с учетом разведения с использованием молярного коэффициента светопоглощения на длине волны 233 нм (=2,2 7105 М-1·см-1) и выражали в нмоль/г ткани (для гомогенатов тканей) или в нмоль/г гемоглобина (для гемолизата эритроцитов).
Активность каталазы определяли методом М.А. Королюка (1988) по разнице содержания Н2О2 в опытных и контрольных пробах в соответствии с законом Бугера–Ламберта–Бэра с учетом молярного коэффициента светопоглощения на длине волны 260 нм, =22 M-1·см-1. Активность выражали в мкмоль/(мин·г белка) или мкмоль/(мин·г гемоглобина) (для эритроцитов).
Активность супероксиддисмутазы (СОД) оценивали по методу В.А. Костюк и соавт. (1990), определяя соотношение Еконтр./ Еопыт, и рассчитывали по формуле в условных единицах (А):
А= Еконтр. Еопыт – 1, где: Еконтр.– изменение оптической плотности за 20 мин. в контрольной пробе; Еопыт – изменение оптической плотности в исследуемой пробе за 20 мин.
Удельную активность СОД выражали в условных единицах, отнесенных к 1мг белка супернатанта или гемоглобина (для эритроцитов).
Активность глутатионпероксидазы (ГП) определяли по методу А.Р. Гавриловой (1986). Расчет активности фермента производили по калибровочной кривой на основании разницы количества восстановленного глутатиона в опытной и контрольной пробах и выражали в ммоль/(мин·г белка) или мкмоль/(мин·г гемоглобина) (для эритроцитов).
Содержание белка определяли методом Лоури в модификации G.L. Peterson(1977) и выражали в г/л.
Содержание гемоглобина в гемолизатах эритроцитов определяли с помощью стандартных наборов “ЭКОлаб” (Россия) гемиглобинцианидным методом и выражали в г/л.
Статистическую обработку полученных результатов проводили на персональном компьютере с помощью пакета прикладных программ Microsoft Excel. В каждой группе рассчитывали средние значения и ошибку от среднего. Оценку различий между выборками осуществляли с использованием t-критерия Стьюдента. Различия сравниваемых результатов (M±m, где M — выборочное среднее арифметическое, m — ошибка среднего арифметического) считались достоверными при достигнутом уровне значимости p<0,05 (Гланц С., 1998).
результаты исследования и их обсуждение
Генопротективные эффекты и механизмы действия мелатонина при химическом воздействии
В результате проведенных исследований установлено, что, спустя сутки после введения циклофосфамида в дозе 200 мг/кг частота хромосомных аберраций в клетках костного мозга крыс достигала почти 10 %. При этом общее количество аберрантных клеток составило 57. Наиболее часто встречались нестабильные хромосомные аберрации: парные фрагменты (в 36 клетках с аберрациями), реже одиночные фрагменты и полиплоиды (в 8 клетках). В единичных случаях (2–3 клетки) отмечались дицентрические хромосомы и транслокации (табл. 1).
Таблица 1
Влияние мелатонина на частоту возникновения и типы хромосомных аберраций, индуцированных циклофосфамидом (200,0 мг/кг), в клетках костного мозга крыс (М±m, n=6)
| Показатель | Группы опытов | ||||
| циклофосфамид (контроль) | Мелатонин, | ||||
| 5 мг/кг | 25 мг/кг | 50 мг/кг | |||
| Частота аберрантных клеток, % | 9,8±1,2 | 9,2±1,1 | 9,3±1,4 | 6,7±1,0* | |
| Общее количество аберраций | 57,0±2,2 | 34,0±2,6* | 44,0±2,4* | 40,0±2,2* | |
| Одиночные фрагменты | 8,0±1,2 | 3,0±0,9* | 4,0±0,9* | 5,0±1,0 | |
| Парные фрагменты | 36,0±2,0 | 25,0±2,1* | 30,0±2,2 | 29,0±2,4* | |
| Дицентрические хромосомы | 2,0±0,8 | 2,0±0,5 | 2,0±0,6 | 1,0±0,4 | |
| Транслокации | 3,0±0,7 | 2,0±0,5 | 2,0±0,6 | 2,0±0,6 | |
| Полиплоиды | 8,0±1,2 | 6,0±1,3 | 6,0±1,3 | 3,0±0,7* | |
Примечание: *) р<0,05 по сравнению с контролем
Как видно из табл. 1, профилактическое (за 30 мин до введения циклофосфамида) применение мелатонина в дозе 5 мг/кг способствовало снижению общего числа хромосомных аберраций на 20%. Существенно снижалось количество клеток с различными типами нарушений генетического аппарата: одиночных и парных фрагментов.
При увеличении дозы препарата до 25 мг/кг общее количество аберраций также снижалось на 20%. Отмечалось уменьшение числа одиночных фрагментов, а также имелась тенденция к снижению парных фрагментов.
Наиболее эффективным защитным действием на генетический аппарат клетки мелатонин обладал в дозе 50 мг/кг. При этом отмечалось не только снижение частоты хромосомных аберраций с 9,8 до 6,7%, общего количества аберрантных клеток (с 57 до 34), но и числа аберраций различных типов (одиночных фрагментов – в 1,6 раза, парных фрагментов – в 1,5 раза, полиплоидов – в 2,5 раза).
Таким образом, мелатонин оказывал отчетливое защитное действие на генетический аппарат клетки, существенно снижая количество хромосомных аберраций в миелокариоцитах костного мозга крыс, отравленных циклофосфамидом.
С целью уточнения механизмов цитопротекторного действия мелатонина изучено его влияние на пролиферативную активность клеток в органотипической культуре ткани селезенки.
Как показано на рис. 1, мелатонин, введенный в культуральную среду в концентрациях 0,01 – 100 нг/мл, дозозависимо изменял пролиферативную активность клеток.
Так, в концентрациях 0,01; 0,05 и 1 нг/мл препарат статистически достоверно увеличивал зоны роста эксплантата: индекс площади в сравнении с контролем составил 124±0,4; 154±1,0 и 122±0,4% соответственно. В концентрациях 10 и 100 нг/мл мелатонин не оказывал статистически достоверного влияния на зоны роста эксплантатов.
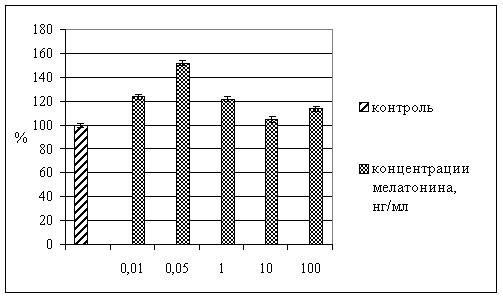
Рис. 1. Влияние мелатонина на скорость роста эксплантата селезенки крыс. По оси абсцисс – концентрация мелатонина. По оси ординат – изменения площади зоны роста эксплантата, % к контролю. *) р<0,05 по сравнению с контрольными значениями.
Циклофосфамид во всем исследованном диапазоне концентраций вызывал торможение роста эксплантатов селезенки (рис. 2).

Рис. 2. Влияние циклофосфамида на рост эксплантатов селезенки крыс. По оси абсцисс – концентрация циклофосфамида. По оси ординат – изменения площади роста эксплантата, % к контролю. *) р<0,05 по отношению к контрольным значениям.
Так, при концентрации цитостатика 0,05 мкг/мл наблюдалось снижение роста эксплантатов до 12% от контрольных значений. При увеличении концентрации циклофосфамида до 100 мкг/мл угнетение развития эксплантата селезенки крыс достигало 84% в сравнении с контролем (рис. 2).
Мелатонин практически во всех исследованных концентрациях предотвращал ингибирующее действие циклофосфамида на рост эксплантата селезенки. Как показано на рис. 3, уже в дозе 0,01 нг/мл мелатонин существенно снижал, а в дозах 0,05–10,0 нг/мл – полностью предотвращал индуцированную цитостатиком задержку зоны роста культуральных клеток. Наконец, в дозе 100,0 нг/мл мелатонин не только нормализовал вызванные циклофосфамидом нарушения клеточной пролиферации, но и способствовал существенному увеличению зоны роста эксплантатов.

Рис. 3. Влияние мелатонина на выраженность ингибирования роста эксплантата селезенки крыс, индуцированной циклофосфамидом (1 мкг/мл). По оси абсцисс – концентрация мелатонина. По оси ординат – изменения площади зоны роста эксплантата, % к контролю. *) р<0,05 по отношению к контрольным значениям.
Таким образом, мелатонин стимулировал пролиферацию клеток селезенки, подавленную циклофосфамидом.
Ниже представлены результаты изучения влияния мелатонина на миграционную активность лейкоцитов периферической крови человека.
На рис. 4 показано, что мелатонин, введенный в образцы крови в концентрациях 0,01 нг/мл – 1 мкг/мл, вызывал увеличение миграционной активности лейкоцитов периферической крови.
В концентрации 1 мкг/мл препарат увеличивал индекс миграционной активности лейкоцитов на 13,0% от контрольных значений. Наибольшее увеличение миграционной активности наблюдалось при добавлении в образцы крови мелатонина в концентрации 1 нг/мл и составило 122,0±2,2%.

Рис. 4. Влияние мелатонина на миграционную активность лейкоцитов периферической крови человека. По оси абсцисс – концентрация мелатонина. По оси ординат – изменения уровня миграционной активности лейкоцитов, % к контролю. *) р<0,05 по сравнению к контрольными значениями.
Циклофосфамид изменял индекс миграционной активности в зависимости от концентрации (рис. 5).

Рис. 5. Влияние циклофосфамида на миграционную активность лейкоцитов периферической крови человека. По оси абсцисс – концентрация циклофосфамида. По оси ординат – изменения уровня миграционной активности лейкоцитов, % к контролю. *) р<0,05 по сравнению с контрольными значениями.
В концентрациях 0,01 – 0,05 мкг/мл отмечалось некоторое увеличение индекса миграции лейкоцитов. Начиная с концентрации 0,5 мкг/мл, наблюдалось достоверное снижение миграционной активности клеток (до 90,0% от контрольных значений).
Мелатонин во всех исследуемых концентрациях нивелировал цитостатический эффект циклофосфамида (рис. 6).
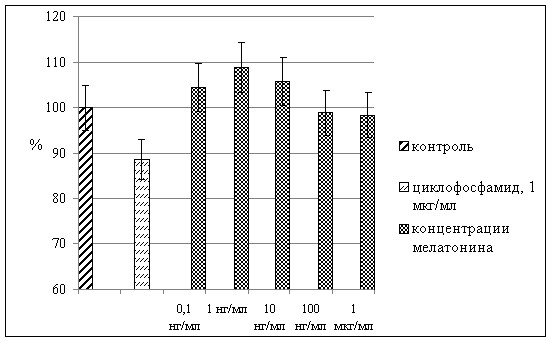
Рис. 6. Влияние мелатонина на миграционную активность лейкоцитов периферической крови человека в присутствии циклофосфамида в концентрации 1 мкг/мл. По оси абсцисс – концентрация мелатонина. По оси ординат – изменения уровня миграционной активности лейкоцитов, % к контрольным значениям. *) р<0,05 по сравнению с контрольными значениями.
Как показали проведенные исследования, после введения крысам циклофосфамида в дозе 200 мг/кг в печени животных отмечалось увеличение количества МДА (более чем в 3 раза по сравнению с биоконтролем), а также повышение активности СОД (табл. 2). В эритроцитах циклофосфамид способствовал увеличению содержания ДК.
Профилактическое введение мелатонина в этих условиях способствовало снижению количества ДК в печени (на 15% по сравнению с контролем), повышению активности каталазы и ГП, а также нормализации активности СОД.
В эритроцитах крыс, отравленных циклофосфамидом, под влиянием мелатонина существенно (на 30 – 40%) снижалось содержание продуктов перекисного окисления липидов (МДА и ДК), отмечалось увеличение активности СОД и каталазы в 1,1 – 1,2 раза.
Таблица 2
Влияние мелатонина в дозе 50 мг/кг на показатели продуктов ПОЛ и антиоксидантной защиты в тканях печени и эритроцитах крыс при остром отравлении циклофосфамидом (200 мг/кг) (M±m, n=50)
| Исследуемый орган (ткань) | Показатель | Группа исследования | ||
| Биоконтроль | Циклофосфамид (контроль) | Мелатонин | ||
| Печень | МДА, нмоль/г ткани | 108,09±17,91 | 362,97±91,57* | 382,91±151,45* |
| ДК, нмоль/г ткани | 39,72±2,193 | 43,43±2,74 | 36,75±3,50^ | |
| СОД, УЕ/мг белка ткани | 5230,3±432,8 | 6526,8±316,7* | 5547,9±358,7^ | |
| Каталаза, моль/мин г белка ткани | 47,24±1,67 | 43,12±3,9 | 50,92±2,53^ | |
| ГП, нмоль/мин г белка ткани | 0,489±0,053 | 0,558±0,086 | 0,78±0,126*^ | |
| Эритроциты | МДА, нмоль/г Hb | 3,58±0,18 | 3,23±0,33 | 2,02±0,17*^ |
| ДК, нмоль/г Hb | 290,66±6,52 | 326,94±16,14* | 222,59±19,03^ | |
| СОД, УЕ/г Hb | 3786,51±98,98 | 3674,42±231,89 | 4371,69±358,82*^ | |
| Каталаза, моль/мин г Hb | 78,87±5,47 | 74,93±3,94 | 82,18±2,4^ | |
| ГП, мкмоль/мин г Hb | 0,7±0,02 | 1,2±0,06 | 1,1±0,02 | |
Приложение: * - р<0,05 по сравнению с группой биоконтроля;
^ - р<0,05 по сравнению с группой контроля (циклофосфамид)
Таким образом, мелатонин у отравленных циклофосфамидом животных повышает активность ферментов антиоксидантной защиты и снижает концентрацию продуктов перекисного окисления липидов, препятствуя повреждению тканей активными формами кислорода.
Генопротективное действие мелатонина при радиационном воздействии и его возможные механизмы
Как показали проведенные исследования, облучение в дозах 2–4 Гр сопровождалось выраженным повреждением генетического аппарата активно пролиферирующих клеток костного мозга крыс. Если у необлученных животных количество клеток с хромосомными аберрациями различных типов составляло в среднем 1,5 %, то через 24 ч после облучения в дозах 2–4 Гр – 14–16 %, т.е. возрастало в 9–10 раз. Результаты представлены в табл. 3.
Таблица 3
Влияние мелатонина в условиях профилактического введения на частоту возникновения и типы хромосомных аберраций в клетках костного мозга крыс, подвергнутых -облучению в дозе 2 Гр (M±m, n=6)
| Показатель | Группы опытов | ||||
| Облучение 2 Гр | Облучение 4 Гр | ||||
| Контроль | Мелатонин, 10 мг/кг | Контроль | Мелатонин, 10 мг/кг | Мелатонин, 50 мг/кг | |
| Частота аберрантных клеток, % | 15,8±0,3 | 8,0±0,1* | 13,8±0,2 | 13,3±0,2 | 6,5±0,1* |
| Общее количество аберраций | 87,0±1,7 | 51,0±1,0* | 92,0±1,8 | 79,0±1,5 | 42,0±0,8* |
| Одиночные фрагменты | 26,0±0,5 | 9,0±0,1* | 32,0±0,6 | 12,0±0,2 | 5,0±0,1* |
| Парные фрагменты | 38,0±0,7 | 18,0±0,3* | 40,0±0,8 | 46,0±0,9 | 28,0±0,5* |
| Дицентрические хромосомы | 17,0±0,3 | 9,0±0,1* | 12,0±0,2 | 0+0,01* | 0+0,01* |
| Транслокации | 2.0±0,04 | 1,0±0,02* | 1,0±0,02 | 3,0±0,06 | 2,0±0,04 |
| Полиплоиды | 4,0±0,08 | 0+0,01* | 8,0±0,1 | 8,0±0,1 | 4,0±0,08* |
Примечание: *) р<0,05 по сравнению с контролем.
Общее количество аберраций составило 85–90, одиночных фрагментов – 25–30, парных фрагментов – 38–40, дицентрических хромосом – 10–15. Значительно реже (в 2–5 случаях) наблюдали наличие транслокаций и полиплоидных клеток.
Мелатонин в дозе 10 мг/кг в условиях профилактического введения существенно уменьшал выраженность постлучевых нарушений в хромосомах клеток костного мозга облученных крыс. Особенно заметным этот эффект был у животных, облученных в дозе 2 Гр. Как видно из табл. 3, под влиянием препарата частота выявления аберрантных клеток снижалась по сравнению с облученным контролем в два раза; общее количество аберраций – на 45%; число одиночных фрагментов – в 3 раза; парных фрагментов, дицентриков и транслокаций – в два раза; полиплоидные клетки обнаружить не удалось.
У крыс, облученных в дозе 4 Гр, применение мелатонина в дозе 10 мг/кг оказалось менее эффективным. С увеличением дозы мелатонина до 50 мг/кг защитное действие препарата на генетический аппарат клеток костного мозга облученных животных существенно увеличивалось и проявлялось уже по пяти показателям (табл. 3).
Так, общее количество аберраций, частота полиплоидных клеток и число парных фрагментов снижались по сравнению с контролем в 1,5–2 раза; количество одиночных фрагментов – в 6 раз; а дицентрические хромосомы вообще не обнаруживались.
Таким образом, в изученных дозах (10 и 50 мг/кг) мелатонин, введенный в профилактических целях, способствовал существенному снижению кластогенного действия облучения у крыс.
В результате исследования обнаружено, что сразу после облучения у подопытных животных наблюдается увеличение интенсивности свободно-радикального окисления (СРО) и активности антиоксидантной системы (по показателям хемилюминесценции на – 12–13%, по показателям ООА и ОАА – на 14–17%).
Через 1 ч после облучения интенсивность процессов СРО по показателю хемилюминесценции Imax/ОЛ увеличивалась на 15% на фоне повышения активности антиоксидантной системы (S/STmax/ОЛ) на 34%. При этом в плазме крови подопытных животных отмечалось увеличение уровня ООА (на 19%) и снижение ОАА (на 10%), что свидетельствует о развитии у них признаков оксидативного стресса (табл. 4).
Применение мелатонина способствовало увеличению интенсивности процессов СРО в плазме крови у необлученных животных (по показателю хемилюминесценции Imax/ОЛ – на 28%, а по показателю ООА/ОЛ – на 20%). Активность антиоксидантной системы в плазме крови у крыс этой группы по показателю хемилюминесценции S/STmax/ОЛ также была повышена на 12%, а по показателю ОАА/ОЛ – на 22%.
Профилактическое применение мелатонина за 30 мин до облучения способствовало увеличению интенсивности процессов СРО в сравнении с группой «контроль I» (по показателю хемилюминесценции на 17% при незначительном увеличении активности антиоксидантной системы), а также снижению уровня ООА при увеличенном уровне ОАА. Через 1 ч после облучения у крыс, которым профилактически вводили мелатонин, по-прежнему отмечалось снижение уровня ООА (до 25%), а ОАА, напротив, увеличивалась на 28%. Профилактическое введение мелатонина приводило к снижению активности антиоксидантной системы в плазме крови у экспериментальных животных по показателю хемилюминесценции S/STmax/ОЛ на 27% по сравнению с «контролем II».
В целом, полученные данные свидетельствуют о стимулирующем действии мелатонина на метаболическую активность в организме необлученных животных, а применение препарата за 30 мин до облучения в дозе 4Гр предотвращает или ослабляет развитие в организме оксидативного стресса.
Таблица 4
Влияние мелатонина (50 мг/кг) на показатели процессов свободно-радикального окисления у крыс, облученных в дозе 4 Гр (M±m, n=6)
| Группы опытов | Сроки измерения показателя | Imax/ОЛ | S/STmaxОЛ | ООА/ОЛ | ОАА/ОЛ |
| Биологический контроль | -- | 0,87±0,01 | 1,87±0,03 | 15,28±0,3 | 9,18±0,1 |
| Контроль I | Через 5 мин после облучения | 0,98±0,011 | 2,12±0,041 | 17,44±0,31 | 10,77±0,21 |
| Контроль II | Через 1 ч после облучения | 1,01±0,021 | 2,49±0,041 | 18,23±0,31 | 8,32±0,11 |
| Мелатонин | Через 30 мин после введения препарата | 1,12±0,021 | 2,09±0,041 | 18,3±0,31 | 11,22±0,21 |
| Облучение + мелатонин | Через 5 мин после облучения | 1,15±0,021 | 2,30±0,041,2 | 15,7±0,31,2 | 10,98±0,21 |
| Облучение + мелатонин | Через 1 ч после облучения | 0,95±0,011,3 | 1,82±0,033 | 13,71±0,21,3 | 10,57±0,21,3 |
Примечание: 1) р<0,05по сравнению с биологическим контролем; 2) р<0,05 по сравнению с контролем I; 3) р<0,05 по сравнению с контролем II.
Таким образом, в исследовании показано, что циклофосфамид и ионизирующее излучение вызывают повреждение хромосомного аппарата миелокариоцитов костного мозга, проявляющееся увеличением уровня хромосомных аберраций в 9–14 раз по сравнению с интактным контролем. При этом рост аберраций происходил за счет увеличения уровня нестабильных аберраций (одиночных, парных фрагментов и дицентрических хромосом). Эти результаты находятся в полном соответствии с данными литературы (Севанькаев А.В. и соавт., 1990; Grisolia C.K., 2002).
Известно, что в результате утраты части генетического материала нестабильные аберрации хромосом создают клеточный дисбаланс, приводят к гибели клетки и, таким образом, подвергаются элиминации. В тканях с ограниченной пролиферативной активностью такой эффект может приводить к невосполнимой утрате специализированных клеточных элементов и создавать основу для размножения менее дифференциальных клеток (Севанькаев А.В. и соавт., 1994). В нашем исследовании установлено, что мелатонин при профилактическом введении существенно снижает общее количество хромосомных аберраций, вызванное введением циклофосфамида или облучением животных. Вызываемое мелатонином снижение уровня аберраций отмечалось преимущественно за счет спектра нестабильных аберраций хромосом (фрагменты и дицентрические хромосомы). В связи с этим не происходит утраты клеточных элементов и большая часть клеток сохраняет свою жизнеспособность и способность к дальнейшему делению. Полученные результаты согласуются с данными, полученными Vijayalaxmi Ph.D. et al. (2004).
Обнаружено, что снижение частоты нестабильных хромосомных аберраций активно пролиферирующих клеток костного мозга позволяет предположить, что введение мелатонина способствует либо сохранению большего количества функционирующих клеток в ряду поколений в момент неблагоприятного воздействия, либо активизации механизмов, восстанавливающих повреждение клеточной ДНК.
Ряд авторов (Голиченков В.А. и соавт., 2010; Jou M.J. et al., 2007; Paradies G. et al., 2010; Reiter R.J. et al., 2007) предполагают, что антикластогенный эффект мелатонина при неблагоприятных воздействиях связан с его способностью улавливать свободные радикалы (в частности, ·ОН) и образовывать менее токсичные соединения. Антиоксидантное воздействие мелатонина, благодаря его липофильности, происходит во всех клеточных структурах (Hardeland R., 2005). Полученные нами данные свидетельствуют, что мелатонин обладает антиоксидантными свойствами при химических и радиационных воздействиях. В присутствии мелатонина активизируется мощный эндогенный фактор ферментативной защиты от радикального окисления – глутатионпероксидаза, которая принимает участие в утилизации активных форм кислорода и продуктов пероксидации, а также восстановлении гидроперекисей жирных кислот и других органических перекисей.
В то же время широту спектра генопротективного действия мелатонина вряд ли возможно объяснить только его антиоксидантными свойствами. Вполне вероятны и другие механизмы защитного действия препарата. Наличие множественных изоформ рецепторов мелатонина на поверхности клеточной мембраны и на мембране ядра клетки обеспечивает избирательность его взаимодействия с естественными лигандами и различия в регуляции экспрессии рецепторов в отдельных тканях (Anisimov S.V et al., 2004).
Изменение индекса площади органотипической культуры лимфоидной ткани является интегральным показателем, отражающим процессы пролиферации и апоптоза. Этот эффект продемонстрирован в работах Н.И. Чалисовой и соавт. (2003), где в экспериментах с пептидами и аминокислотами показано, что при увеличении зоны роста эксплантатов увеличиваются количественные плотности PCNA-позитивных ядер и интерфазных клеток, содержащих ядрышковые организаторы, что свидетельствует об усилении пролиферативных процессов. Одновременно происходит снижение индекса апоптоза и экспрессии проапоптозного белка р53. Обнаружена также отрицательная корреляция между экспрессией проапоптозного белка р53 и степенью клеточной пролиферации (Чалисова Н.И. и соавт., 2003, 2006).
В связи с этим добавление мелатонина в культуральную среду могло привести к активации пролиферации клеток за счет снижения экспрессии белка р53 и индекса апоптоза. В пользу такого предположения свидетельствуют работы некоторых исследователей (Gonca A.K. et al., 2011; Baydas G et al., 2007).
Таким образом, проведенные исследования убедительно показывают, что мелатонин при однократном профилактическом применении обладает антикластогенными свойствами как при токсических, так и при ионизирующих воздействиях, усиливая антиоксидантную и стимулируя пролиферативную и миграционную активность клеток.
В целом полученные в работе данные свидетельствуют о перспективности использования синтетического аналога эпифизарного гормона мелатонина для профилактики развития неблагоприятных цитогенетических последствий воздействия на организм факторов радиационной и химической природы.
ВЫВОДЫ
- Циклофосфамид в дозе 200 мг/кг вызывает существенное повреждение генетического аппарата соматических клеток, проявляющееся в увеличении общего количества хромосомных повреждений до 10% от контрольных значений (главным образом, нестабильных аберраций). Мелатонин, введенный в дозах 5 – 50 мг/кг за 30 мин до цитостатика, способствует снижению выраженности индуцированных циклофосфамидом повреждений хромосомного аппарата: общее число аберраций снижается в 1,4 – 1,6 раза, количество одиночных фрагментов уменьшается в 1,5 – 2,5 раза, парных фрагментов – в 1,2 – 1,4 раза, полиплоидов – в 1,3 – 2,5 раза;
- При добавлении в культуральную среду циклофосфамида в концентрациях 0,05 – 100 мкг/мл наблюдается значительное угнетение пролиферативной активности спленоцитов (до 84% по сравнению с контрольными значениями). Мелатонин в концентрациях 0,05–10 нг/мл полностью предотвращает индуцированную цитостатиком задержку зоны роста клеток, а в концентрации 100 нг/мл помимо нормализации нарушения клеточной пролиферации, способствует существенному увеличению зоны роста эксплантатов.
- Циклофосфамид в концентрации от 0,5 мкг/мл и выше, существенно снижает миграционную активность лейкоцитов периферической крови человека. Мелатонин в концентрациях 0,1 нг/мл–1,0 мкг/мл нивелирует выявленный эффект цитотоксиканта, увеличивая уровень миграционной активности лейкоцитов;
- В дозе 200 мг/кг циклофосфамид вызывает активацию процессов перекисного окисления липидов, что выражается в значительном увеличении содержания малонового диальдегида в печени (более чем в 3 раза) и диеновых конъюгат в эритроцитах (в 1,2 раза). Мелатонин способствует снижению содержания этих продуктов (особенно в эритроцитах) и активирует ферменты перекисной защиты (супероксиддисмутаза, каталаза, глутатионпероксидаза);
- Под влиянием ионизирующего излучения в дозах 2–4Гр общее количество хромосомных аберраций в клетках костного мозга крыс возрастает до 15%. Мелатонин в дозах 10–50 мг/кг, введенный за 30 мин до облучения, способствует существенному снижению (в 2 – 3 раза) кластогенного эффекта радиационного воздействия преимущественно за счет спектра нестабильных хромосомных аберраций (фрагменты, дицентрици, полиплоиды).
- Облучение крыс в дозе 4Гр сопровождается увеличением интенсивности процессов перекисного окисления липидов и снижением активности антиоксидантной системы, что свидетельствует о развитии пострадиационного оксидативного стресса. Применение мелатонина за 30 мин до облучения способствует нормализации процессов свободно-радикального окисления. У необлученных крыс мелатонин вызывает увеличение как прооксидантной, так и антиоксидантной актиности.
ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ
- Мелатонин целесообразно использовать для профилактики повреждающего действия на хромосомный аппарат клетки токсикантов (цитостатиков) и ионизирующего излучения;
- Использованные в ходе исследования критерии оценки кластогенного действия ионизирующего излучения и цитостатиков могут быть применены при отборе перспективных средств профилактики генотоксических эффектов широкого спектра соединений, потенциально опасных для хромосомного аппарата клетки.
список основных работ,
опубликованных по теме диссертации
- Пикалова Л.В. Применение цитогенетических методов исследования хромосом в радиологии // Medline.ru. – 2008, Т.9, ст. 17. – С. 160-168. – Режим доступа: http//www.medline.ru, свободный.
- Легеза В.И. Отдаленные последствия у ликвидаторов аварии на Чернобыльской АЭС / В.И. Легеза, А.Е. Антушевич, Л.В. Пикалова, А.Н. Жекалов // Medline.ru. – 2008, Т.9, ст. 33. – С. 362-372. – Режим доступа: http//www.medline.ru, свободный.
- Пикалова Л.В. Перспективы использования Daphnia magna straus в качестве тест-объекта при определении мутагенной активности загрязнителей окружающей среды // Вестник Российской Военно-медицинской Академии. Приложение II. – СПб, 2008, № 3 (23). – С.146.
- Пикалова Л.В. Генопротективные эффекты мелатонина у крыс, облученных гамма-излучением / Л.В. Пикалова, А.А. Сазанов // Лицейские чтения: Шаг в будущее: современное студенчество как творческий, научный и профессиональный потенциал России: материалы I Междунар. науч. студ. конф. – СПб: ЛГУ им. А.С. Пушкина, 2009. – С.75.
- Чалисова Н.И. Протекторное влияние пептидов и аминокислот на развитие культуры лимфоидной ткани в присутствии циклофосфана / Н.И. Чалисова, М.Б. Иванов, Л.Г. Аржавкина, Л.В. Пикалова, А.В. Смирнов // Токсикологический вестник. – 2010. - №4. – С.41-45.
- Пикалова Л.В. Генопротективные эффекты мелатонина при экстремальных радиационных воздействиях на живые системы / Л.В. Пикалова, В.И. Легеза, М.Б. Иванов // Сборник тезисов докладов VII Всероссийской межвузовской конференции молодых ученых, вып. 3. – СПб, 2010. – С. 74-75.
- Легеза В.И. Экспериментальное изучение генопротективных эффектов мелатонина при радиационном воздействии / В.И. Легеза, М.Б. Иванов, Л.В. Пикалова // VI съезд по радиационным исследованиям (радиобиология, радиоэкология, радиационная безопасность): Тезисы докладов. Т.I. – М.: РУДН, 2010. – С.63.
- Пикалова Л.В. Изучение генопротективных свойств мелатонина при гамма-облучении в эксперименте / Л.В. Пикалова, В.И. Легеза, М.Б. Иванов // Медицинская генетика. Материалы VI съезда всероссийского общества медицинских генетиков. – Ростов-на-Дону. – 2010. – С.140.
- Пикалова Л.В. Защитное действие пептидных препаратов на иммунокомпетентные клетки селезенки при поражении циклофосфаном в эксперименте / Л.В. Пикалова, М.Б. Иванов, Л.Г. Аржавкина, С.В. Петленко, Н.И. Чалисова // Актуальные проблемы медицины и биологии // Сборник тезисов докладов научно-практической конференции / под ред. акад. РАМН А.В. Шаброва, проф. В.Г. Маймулова. – СПб: СПбГМА им. И.И. Мечникова. – 2010. – С. 115-116.
- Иванов М.Б. Механизмы генопротективного действия мелатонина при радиационном воздействии / М.Б. Иванов, Л.В. Пикалова, В.И. Легеза, Н.И. Чалисова // Актуальные проблемы медицины и биологии // Сборник тезисов докладов научно-практической конференции / под ред. акад. РАМН А.В. Шаброва, проф. В.Г. Маймулова. – СПб: СПбГМА им. И.И. Мечникова. – 2010. – С. 110-111.
- Пикалова Л.В. К вопросу о возможности использования мелатонина в качестве генопротекторного средства при чрезвычайных ситуациях радиационной и химической природы / Л.В. Пикалова, В.И.Легеза, М.Б. Иванов, Н.И. Чалисова, Е.Б. Жаковко // Чрезвычайные ситуации и эпидемиологические проблемы. Материалы XXXXIII научной конференции СПбМАПО «Хлопинские чтения». – СПб: СПбМАПО, 2010. – С. 93-95.
- Пикалова Л.В. Перспективы использования мелатонина для профилактики хемоиндуцированного повреждения клеток / Л.В. Пикалова, М.Б.Иванов, С.В. Петленко, Н.И. Чалисова, Е.Г. Богданова, А.В. Смирнов // Химическая безопасность Российской Федерации в современных условиях (Сборник трудов Всероссийской научно-практической конференции) / под ред. д.м.н., профессора В.Р. Рембовского и д.м.н., профессора А.С. Радилова. – СПб: Фолиант, 2010. – С.359.
- Пикалова Л.В. Возможность использования мелатонина для профилактики кластогенных эффектов факторов окружающей и производственной среды / Л.В. Пикалова, В.И. Легеза, М.Б. Иванов, С.В. Петленко, Е.Г. Богданова // Научно-методические и законодательные основы обеспечения генетической безопасности факторов и объектов окружающей и производственной среды в целях сохранения здоровья человека // Материалы объединенного Пленума. – М. – 2010. – С.117-118.
- Петленко С.В. Новый подход к интегральной оценке иммунной системы человека в условиях воздействия комплекса факторов химически опасных объектов / С.В. Петленко, С.П. Лось, М.Б. Иванов, Е.Г. Богданова, А.В. Голубков, С.Б. Комнатный, Л.В. Пикалова // Medline.ru. – 2010. – Т.11 (ст. 17). – С. 195-216. – Режим доступа: http//www.medline.ru, свободный.
- Пикалова Л.В. Экспериментальное исследование цитопротективного действия мелатонина при радиационном воздействии / Л.В. Пикалова, В.И. Легеза, М.Б. Иванов, Е.Б. Жаковко // Бюллетень экспериментальной биологии и медицины. – 2011. – Т.152, №7. – С.83-86.
- Пикалова Л.В. К вопросу о механизмах защитного эффекта мелатонина при радиомиметических воздействиях / Л.В. Пикалова, В.И. Легеза, М.Б. Иванов, С.В. Петленко, Е.Г. Богданова // Актуальные проблемы токсикологии и радиобиологии: Тезисы докладов Российской научной конференции с международным участием. – СПб: ООО «Издательство Фолиант», 2011. – С.238.
- Пикалова Л.В. Влияние циклофосфана на пролиферативную и митогенно-миграционную активность клеток в эксперименте (in vitro) / Л.В. Пикалова, В.И. Легеза, М.Б. Иванов, С.В. Петленко, Е.Г. Богданова, Н.И. Чалисова, М.А. Войцеховская, Е.А. Концевая // Эколого-гигиенические и клинические проблемы управления здоровьем населения. Материалы XXXXIV научной конференции СПбМАПО «Хлопинские чтения» / Под ред. А.П. Щербо. – СПб.: Издательство СПбМАПО, 2011. – С.221-222.
- Пикалова Л.В. Экспериментальное изучение генопротективных свойств мелатонина при химическом воздействии // Актуальные вопросы клинической и экспериментальной медицины: сборник тезисов юбилейной научно-практической конференции молодых ученых / Под ред. А.В. Силина, И.Ю. Стиф. – СПб: СПбМАПО, 2011. – С.43-44.
- Голубков А.В. Методология иммуноэпидемиологических исследований при профессиональном и экологическом воздействии факторов химической опасности / А.В. Голубков, С.В. Петленко, М.Б. Иванов, Е.Г. Богданова, Д.А. Синячкин, Л.В. Пикалова // Medline.ru. – 2011. – Т.12 (ст. 3). – С. 21-31. – Режим доступа: http//www.medline.ru, свободный.