

Анализ изменчивости комплексов количественных признаков как методология эколого-генетического изучения селекционируемых и естественных популяций рыб
На правах рукописи
ТЮРИН Владислав Викторович
АНАЛИЗ ИЗМЕНЧИВОСТИ КОМПЛЕКСОВ КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ КАК МЕТОДОЛОГИЯ ЭКОЛОГО-ГЕНЕТИЧЕСКОГО ИЗУЧЕНИЯ СЕЛЕКЦИОНИРУЕМЫХ И ЕСТЕСТВЕННЫХ ПОПУЛЯЦИЙ РЫБ
Специальность 03.02.06 - ихтиология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
доктора биологических наук
Краснодар – 2010
Работа выполнена в Кубанском государственном университете на кафедре генетики, микробиологии и биотехнологии
Научный консультант: доктор биологических наук, профессор
Волчков Юрий Андреевич
Официальные оппоненты: доктор биологических наук, профессор,
Абросимова Нина Акоповна
доктор биологических наук, профессор, заслуженный деятель науки России Пономарев Сергей Владимирович
доктор сельскохозяйственных наук, профессор
Скляров Валентин Яковлевич
Ведущая организация: Государственный научно-исследовательский институт озерного и речного рыбного хозяйства (ГосНИОРХ)
Защита диссертации состоится 27 декабря 2010 года в 12 часов на заседании диссертационного совета Д 212.101.14 в Кубанском государственном университете по адресу: 350049, г. Краснодар, ул. Ставропольская, 149, КубГУ.
С диссертацией можно ознакомиться в библиотеке Кубанского государственного университета.
Автореферат разослан «___» ___________ 2010 года.
Ученый секретарь диссертационного совета
доктор биологических наук, профессор Пескова Т.Ю.
ОБЩАЯ ХАРАКТРИСТИКА РАБОТЫ
Актуальность темы исследования.
Исторически ихтиология оказалась первой биологической наукой, обратившейся для решения ряда задач, в частности изучения популяционной структуры вида, к исследованию комплекса признаков (Heincke, 1889). Дальнейшее развитие многомерного подхода при разработке этой проблемы, а также в изучении полового диморфизма, межвидовой гибридизации, полиплоидии и некоторых вопросов селекции создали необходимую базу для обобщения практического опыта. Общее мнение, разделяемое и исследователями других объектов, состоит в признании существенных преимуществ многомерного подхода к изучению изменчивости перед исследованием отдельных признаков. По выражению Р. Левонтина (1978), он позволяет перейти «от того, что можно измерить, к тому, что есть на самом деле».
По мере расширения области применения многомерного подхода все боле ясной становится его адекватность решению эколого-генетических проблем. Обсуждая перспективы развития теории микроэволюции, Н. В. Глотов (1988), назвав два уже пройденных этапа в ее становлении: работу С.С. Четверикова (1926), органически связавшую генетику и классический дарвинизм, и формирование синтетической теории эволюции, - указал на ожидаемый третий этап синтеза, а именно эколого-генетический. Ожидание тем более оправдано, если речь идет о признаках количественных, которые очевидно предпочтительнее качественных в оценке эффектов взаимодействия «генотип х среда».
Тезис одного из основоположников фенетики А.В.Яблокова (1982), утверждающего, что использование большого числа признаков для описания объектов «увеличивает генетическую мощность анализа фенотипической изменчивости» принципиально расширяет область применения анализа комплекса признаков. В сферу изучаемых проблем естественно включаются: выявление и оценка генетической гетерогенности популяций; изучение динамики их генетической структуры под действием естественного и искусственного отбора; выявление элементов внутрипопуляционной структуры, с которыми оперирует отбор и центральная проблема селекции – распознавание селекционно ценных генотипов по фенотипу.
Теоретическое и экспериментальное обоснование перспективности анализа изменчивости комплекса признаков в качестве методологии исследования перечисленных проблем селекционируемых и природных популяций рыб и определяет актуальность данного исследования.
Цель исследования: с теоретических позиций и по итогам экспериментов обосновать системный анализ изменчивости количественных признаков как наиболее перспективную методологию решения проблем экологической генетики и селекции рыб; усовершенствовать его методы за счет выбора оптимальных категорий признаков и математико-статистических методов анализа.
Задачи исследований в области теории:
- на основе новых экспериментальных данных подтвердить эффективность системного анализа изменчивости комплексов морфологических и остеологических признаков во вскрытии генетической гетерогенности искусственных и естественных популяций рыб;
- в рамках этого подхода выявить элементы структуры популяций рыб, с которыми оперирует отбор, и оценить эффекты естественного и искусственного отбора как фактора динамики популяций;
Прикладные задачи:
- предложить методы оценки перспективности естественных или искусственных популяций как исходного материала для селекции и воспроизводства;
- разработать эколого-генетически обоснованный метод оценки продуктивности в селекции рыб при прудовом выращивании;
- оптимизировать методы распознавания селекционно ценных групп рыб (семей, гибридов) на основе анализа изменчивости комплекса коррелированных признаков.
Научная новизна
С учетом ранее описанных примеров успешного использования системного анализа изменчивости количественных признаков и собственных экспериментальных результатов обоснована универсальность этого подхода в решении задач экологической генетики и селекции рыб. Предложено и испытано сочетание методов кластерного и дискримиантного анализов, позволяющее выявлять генетический полиморфизм популяций рыб, с элементами которого оперирует естественный и должен оперировать искусственный отбор.
На примере исследования популяций черноморской мидии (Mytilus galloprovincialis Lam.) показана возможность расширения сферы применения методов на другие объекты аквакультуры.
Практическая значимость
Разработан эколого-генетически обоснованный метод сравнения различных групп рыб (смей, пород, поколений селекции) по продуктивности при прудовом выращивании, основанный на апостериорной минимизации модифицирующих эффектов различия условий среды.
Предложен новый принцип выбора селекционно-перспективных групп рыб, основанный на их сравнении с селекционной моделью («идеальным» объектом).
Предложен и испытан комплекс многомерных статистических методов, обеспечивающий мониторинг генетической структуры материала по результатам анализа изменчивости системы морфотипа в процессе синтетической селекции.
С эколого-генетических позиций обоснована возможность оптимизации искусственного воспроизводства рыб, основанная на преимущественном размножении тех распознаваемых по морфотипу элементов генетической структуры искусственных популяций, которые численно преобладают в естественных условиях.
Положения, выносимые на защиту:
Системный анализ – оптимальная методология эколого- и селекционно-генетических исследований искусственных и природных популяций рыб.
Системный подход эффективно реализуется в исследовании изменчивости комплексов количественных признаков методами многомерного статистического анализа.
В этой форме его реализации подход обеспечивает выявление и оценку генетической гетерогенности популяций, сопряженной с адаптивностью, на основе анализа только фенотипической изменчивости. Тот же подход позволяет оценить уровень генетически обусловленных различий между группами объектов (породами рыб, семьями от индивидуальных скрещиваний, последовательными поколениями синтетической селекции, маточными стадами, природными и искусственными популяциями).
В рамках системного анализа изменчивости комплексов морфометрических и остеологических признаков возможна оценка эффектов естественного и искусственного отбора и определение «точек» его приложения.
Апробация работы. Результаты исследований докладывались на 11-м всесоюзном совещании «Рыбохозяйственное освоение растительноядных рыб» (Кишинев, 1988); Региональных конференциях «Актуальные вопросы изучения экосистемы бассейна Кубани» (Краснодар, 1988); «Актуальные вопросы экологии и охраны природы Азовского моря и Восточного Приазовья» (Краснодар, 1989); «Актуальные вопросы экологии и охраны природы экосистем малых рек» (Краснодар, 1990); «Современные проблемы экологии» (Анапа, 1996); межреспубликанских научно-практических конференциях «Актуальные вопросы экологии и охраны природы южных и центральных регионов России» (Краснодар, 1997 – 2009); международной научно-практической конференции «Проблемы воспроизводства растительноядных рыб и их роль в аквакультуре» (Адлер, 2000); всероссийской конференции «Проблемы и перспективы развития аквакультуры в России» (Краснодар 2001); международной научной конференции «Проблемы естественного и искусственного воспроизводства рыб в морских и пресноводных водоемах» (Ростов-на-Дону 2004).
Публикации. По материалам диссертации опубликовано 49 работ, в том числе 9 в изданиях из перечня ВАК, рекомендуемых для соискания ученой степени доктора наук.
Структура и объем диссертации. Диссертация изложена на 272 страницах, состоит из введения, пяти глав, выводов, содержит 84 таблицы и 46 рисунков. Список использованной литературы выключает 243 наименования, в том числе 57 на иностранных языках.
Автор выражает глубокую благодарность доктору биологических наук, профессору В.С. Кирпичникову за первые уроки в области генетики и селекции рыб; доктору биологических наук, профессору Ю.А. Волчкову, кандидатам биологических наук С.И. Решетникову, М.В. Ганченко и В.П. Радецкому за оказанную помощь в выполнении и подготовке данной работы.
ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ
Глава 1. Принципы анализа комплексов признаков в селекционно-генетических и популяционных исследованиях
В главе рассматриваются проблемы идентификации селекционно-ценных генотипов по фенотипу, внутривидовая структура с позиций экологической генетики, методы анализа внутрипопуляционной изменчивости, основные направления селекции рыб и их генетические предпосылки.
Глава 2. Материал и методы исследования
В качестве основных объектов исследования выступали следующие представители пресноводной аквакультуры: карп (Cyprinus carpio L.), белый амур (Ctenopharyngodon idella Val.) - БА, белый толстолобик (Hypophthalmichthys molitrix Val.) - БТ, пестрый толстолобик (Aristichthys nobilis Rich.) - ПТ, черноморская кумжа (Salmo trutta labrax Pall), радужная форель (Salmo gairdneri Gibb.). В качестве специального, дополнительного выполнено исследование на черноморской мидии (Mytilus galloprovincialis Lam.).
Для разработки генетически обоснованных методов искусственного отбора необходим экспериментальный материал с гарантированными (или весьма вероятными) генетическими различиями между группами. В исследованиях по теории отбора такие группы могут выполнить роль обучающей выборки при разработке методов распознавания селекционно-ценных генотипов по фенотипу. В данной работе группы, отвечающие этому критерию, были получены в результате четырех различных категорий экспериментов:
- постановка индивидуальных скрещиваний у растительноядных рыб (РЯР) с последующим сравнительным изучением семей;
- выборки из пород и породных групп карпа, взятые из потомств от массовых внутрипородных скрещиваний, и аналогичные выборки из пород форели;
- выборки из простых и сложных гибридов местного и интродуцированных пород карпа, полученных в процессе синтетической селекции;
- выборки из маточных стад рыборазводных заводов и естественных популяций черноморской кумжи.
Исследуемый материал в разной мере был охарактеризован по комплексу признаков продуктивности, комплексу морфометрических и иногда остеологических признаков.
В четырех перечисленных категориях экспериментов общее количество групп, оцененных по продуктивности, составило более 200; общее количество особей, описанных по морфотипу – более 2000; по параметрам осевого скелета – более 300.
В ходе статистической обработки данных использованы как методы классической биометрии, так и многомерные: метод главных компонент, факторный, дискриминантный и кластерный анализы. Ряд существенных деталей по материалу и используемым методам излагается по ходу анализа результатов исследований. Специального предварительного обсуждения заслуживает только дискриминантный анализ, сыгравший особо важную роль в данном исследовании (Клекка, 1989).
Суть подхода иллюстрирует рис. 1 (Тюрин и др., 2003). Пусть имеются две группы объектов (А и В), описанные по двум переменным. Переменные образуют плоскость, на которой каждая из групп представлена в виде облака точек. При рассмотрении проекций крайних объектов каждой группы, например на ось Х, видна область перекрывания групп. Это объекты в интервале xb1 – xa2.

Рис. 1 - Геометрическая интерпретация результатов дискриминантного анализа
А, В – центроиды разделяемых групп; xa1, xb1 – минимальные значения признака Х в группах А и В; xa2, xb2 – максимальные значения признака в этих группах; dfa1, dfa2 – проекции конкретных объектов группы А на ось дискриминантной функции; dfb1, dfb2 – тоже для конкретных объектов группы В.
Но можно построить новую ось (на рис. 1 – DF), значения которой по иному учтут исходную изменчивость. Внутригрупповая изменчивость будет минимизирована и, за счет этого, четко будут выражены межгрупповые различия – перекрывание облаков точек исключено. Новая ось - новый интегральный признак является линейной комбинацией исходных признаков – дискриминантной функцией.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Глава 3. Апостериорная минимизация эффектов среды как метод выявления генотипических межгрупповых различий по продуктивности при прудовом выращивании рыб
При оценке групп рыб по продуктивности каждая из них характеризовалась по комплексу из восьми показателей. Первую их категорию - «признаки - компоненты продуктивности», составили средняя конечная масса рыб (mk), выход рыбы с единицы площади пруда - рыбопродуктивность (pr), прирост массы за сезон (dm), скорость роста (v), выживаемость (ft). Во вторую – «параметры рыбоводного эксперимента», вошли средняя посадочная масса рыб (m0), плотность посадки (pp) и плотность выхода (pv).
Как показали результаты экспериментов по оценке комплекса признаков продуктивности рыб, различия факторов среды неизменно влияют на выращиваемый материал, изменяя его характеристики. В этом смысле продуктивность оказывается, выражаясь математически, функцией многих переменных. Их совокупный эффект столь велик, что выявление генетически обусловленных различий по продуктивности превращается в серьезную проблему. В значительной части сравнительных экспериментов вклад межсемейных (межпородных) различий в общую дисперсию результирующего показателя - выхода рыбы с единицы площади очень мал или отсутствует даже в условиях совместного выращивания групп (таблица 1).
Таблица 1 - Результаты анализа структуры изменчивости рыбопродуктивности (ц/га) в семьях двухлеток белого толстолобика (на примере потомств от индивидуальных скрещиваний 1981 г.)
| Изменчивость | df | mS | F | 2 | Доля в общей дисперсии, % |
| Между семьями | 6 | 3801,0 | 7,4* | 470,0 | 9,7 |
| Между прудами | 2 | 27487,0 | 53,7* | 3853,0 | 79,7 |
| Взаимодействие | 1 | 99,0 | 0,2 | 0,0 | 0,0 |
| Остаточная | 11 | 512,0 | 512,0 | 10,9 |
При столь малом вкладе межсемейной дисперсии вопрос о выборе лучших семей практически не решается без минимизации модифицирующих эффектов условий выращивания. Минимизация может быть только апостериорной, то есть выполненной в ходе специального статистического анализа данных эксперимента. Действительно, условия выращивания рыб в конкретных прудах подлежат только контролю, но не регулированию.
Апостериорно «снять» некоторые средовые эффекты при оценке межгрупповых различий позволяет, например, ковариационный анализ. С его помощью достигается статистический аналог эксперимента, в котором средовые характеристики – pp, pv и m0 были бы одинаковые. Последние вводятся в виде ковариат и позволяют корректировать значения компонент продуктивности в рамках регрессионных процедур.
Безусловно, характеристики рыбоводного режима выращивания: pp, pv, m0 во многом определяют состояние экосистемы пруда и по терминологии Э.Х. Гинзбурга (1984) могут быть названы «эффективными». Однако, выращивание рыбы в разных прудах с равной плотностью и одинаковой посадочной массой не означает уравнивание всех средовых условий. Полная коррекция значений компонент продуктивности потребовала бы измерения и последующего снятия эффектов достаточно большого числа ковариат. Практически это невыполнимо.
Но не технические трудности составляют главную проблему ковариационного анализа. Он не может стать универсальным средством снятия средовой компоненты изменчивости в силу несовершенства самого подхода с биологических позиций. Факторы среды влияют на объект выращивания не порознь, а комплексно, и сила и продолжительность их воздействия находит свое отражение во взаимосвязанных изменениях признаков - компонент продуктивности. Следовательно, адекватная оценка генетически обусловленных различий групп может быть получена только в рамках системного анализа, когда одновременно исследуются обе категории признаков системы «зарыбленный пруд»: характеризующие и условия выращивания, и сам объект.
Наилучшее доказательство взаимозависимости элементов системы может быть получено в результате анализа структуры их связей. Корреляционный анализ выявил достоверную корреляцию всех 8 показателей, включенных в систему оценки продуктивности. Однако факторный анализ позволил объединить показатели в три группы - плеяды (рис. 2).
Первую плеяду составляют скорость роста, прирост и конечная масса тела (v, dm, mk). Ее поэтому естественно интерпретировать как «плеяду массонакопления». Во вторую – «плеяду качества посадочного материала» вошли начальная масса тела и выживаемость (m0, ft). Очевидно, что уровень выживаемости в определенной мере обусловлен генетически, но постоянство выделения плеяды в таком составе заставляет видеть в различии стартовой массы рыб основной источник межгрупповой изменчивости по выживаемости. От вида к виду в наших опытах изменялась только сила положительной связи между ft – m0: БТ 0,89; БА 0,21; ПТ 0,20; карп 0,24. Стабильность плеяды и определила ее название по ведущему показателю.
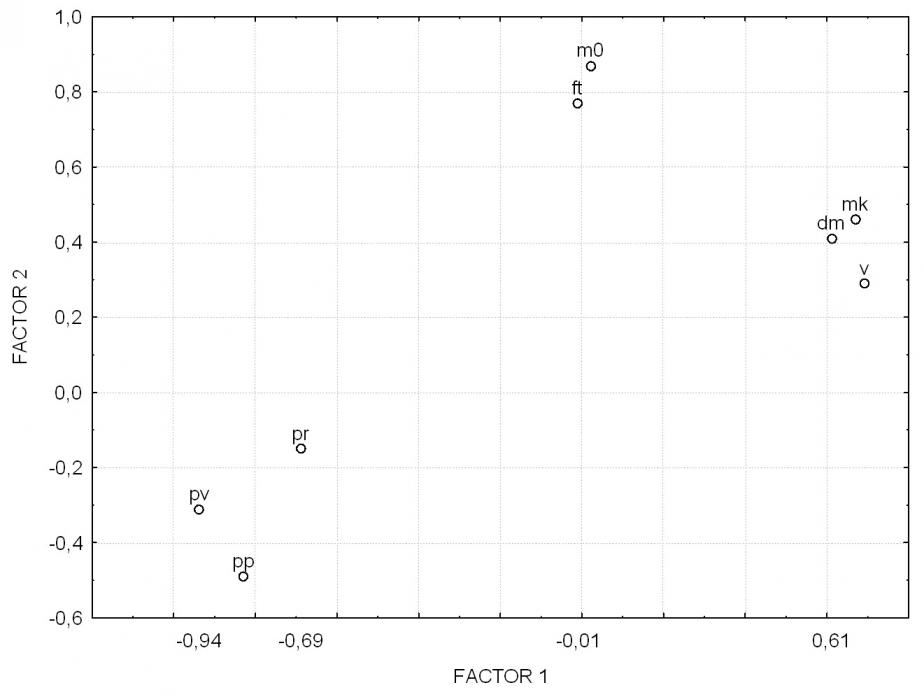
Рис.2 - Распределение признаков в пространстве первого и второго факторов
Третья плеяда образована теми характеристиками системы, в значениях которых, главным образом, заключена информация о рыбоводном режиме выращивания и среде обитания. Это плотность посадки, плотность по выходу и рыбопродуктивность (pp, pv, pr). За ней оставлено название «плеяды условий выращивания».
В существовании трех различных плеяд и содержится ключ к решению проблемы минимизации эффекта сопутствующих факторов при сравнении групп рыб по продуктивности. Следует только уточнить постановку вопроса, подчеркнув как необходимость так и возможность такой минимизации.
Необходимость связана с тем, что показатели плеяд «условий выращивания» и «качества посадочного материала» не подлежат регулированию, и, поэтому, неизбежно варьируют от опыта к опыту. Сколь угодно «одинаковые» пруды фактически различаются по гидрохимическим условиям, температуре воды и ее колебаниям, кормности и другим характеристикам. Равная, например нормативная, плотность посадки не гарантирует одинаковой плотности выращивания из-за различной выживаемости рыб. Практически невозможно устранить без нежелательного в сравнительных опытах отбора и различия групп по стартовой массе тела, особенно если сопоставляются не сеголетки, а последующие возрастные группы рыб.
Возможность обеспечивается относительно независимым варьированием показателей разных плеяд. Оно означает, что на основе учтенного комплекса признаков возможно конструировать такие интегральные оценки семей - линейные комбинации, куда характеристики «объекта» войдут с максимальными, а характеристики «условий» - минимальными (в идеале нулевыми) вкладами.
Селекционно-полезная и генетически обоснованная оценка групп по продуктивности должна удовлетворять двум требованиям: максимум информативности по признакам - компонентам продуктивности и минимум средовой компоненты в структуре изменчивости. Оба требования выполняются, когда способ построения линейных комбинаций отвечает их целевой функции. Ее следует определить как нахождение вектора значений признаков, позволяющего наилучшим образом различить групповые средние. Реализовать такую функцию предлагается в дискриминантном анализе, в который
Когда в дискриминантный анализ вовлекаются группы, объединяющие выборки рыб одной и той же семьи или породы из разных опытов - прудов, в изменчивости значений линейных комбинаций - дискриминантных функций - соотношение дисперсий «между семьями» и «между прудами» оказывается оптимальным для решения поставленной задачи. Межсемейные (межпородные) различия оцениваются на фоне апостериорно минимизированных межпрудовых.
Высокую эффективность дискриминантного анализа в решении поставленной задачи можно иллюстрировать на примере анализа распределений в пространстве дискриминантных функций трех семей БТ 1+ выращенных в 4 прудах в условиях совместной посадки (рис.3).
На рисунке разделение 12 объектов анализа (3 семьи х 4 пруда) за счет минимизации различий в условиях выращивания (разные пруды) разделяются только на три группы - по семейной принадлежности.
Возможность «свертывания» информации об объекте в значение линейной комбинации еще не решает задачу полностью, хотя расстояния между объектами отбора – семьями в пространстве дискриминантных функций приобретает смысл генетических. Значения новых признаков, какими являются линейные комбинации, не могут быть переведены в масштаб селекционно-значимых, и критерий отбора лучших групп еще следует определить.
Нами предложено сравнение групп с некоторым «идеальным для данных условий» объектом (Тюрин, Волчков, 1993; Тюрин, 2008). В определенном смысле его можно назвать селекционной моделью, максимальное приближение к которой и составляет цель селекции.
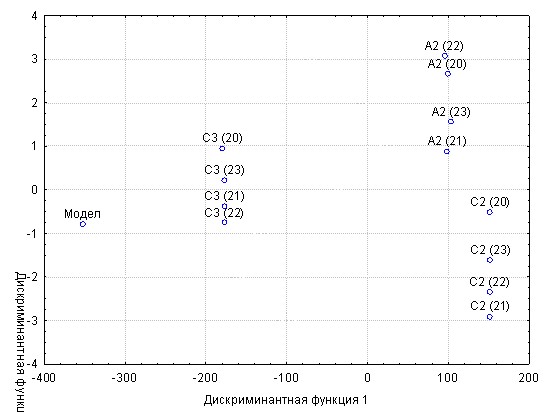
Рис.3 - Распределение семей белого толстолобика генерации 1988 г., пространстве дискриминантных функций
А, С – самки; 2,3 – самцы; 20, 21, 22, 23 – номера прудов.
Подход можно реализовать, поместив в пространство дискриминантных функций некоторую точку (Модель). Ее координаты определяются подстановкой в дискриминантные функции наибольших из встреченных в сравнительном эксперименте значений признаков – компонент продуктивности и средних значений показателей условий выращивания. Задача сравнения групп сводится при этом к оценке их расстояний до модели, и лучшая группа выбирается по наименьшему из них. Тем самым выбор критерия отбора возвращается в русло оценки генетических расстояний, когда получаемая селекционная оценка группы отражает меру ее близости к «идеальному объекту».
Судя по ординации точки модели наиболее близка к ней семья С3. Однако этот факт должен быть подтвержден статистически. В силу ортогональности пространства дискриминантных функций расстояние до точки модели можно вычислить как евклидово, что и было сделано для всех 12 опытов. Дисперсионный анализ изменчивости евклидовых расстояний выявил абсолютное доминирование межсемейной дисперсии в общей – 99,9%. Ранговый тест, выполненный с помощью множественного критерия Стьюдента, указал на статистически достоверные различия всех трех семей по расстоянию до модели (табл.2).
Таблица 2 - Множественный ранговый тест на достоверность различия групповых евклидовых расстояний до модели, усл. ед.
| Семья | Среднее евклидово расстояние до модели | Ранговый тест средних | ||
| C3 | 425,56 | * | ||
| A2 | 872,38 | * | ||
| C2 | 1127,29 | * | ||
| Примечание. Расположение звездочек на разных вертикалях теста свидетельствует о статистически достоверных различиях (p<0,01) | ||||
Аналогично построенный анализ выполнен также на данных по выращиванию двухлеток и трехлеток семи пород карпа, пяти семей двухлеток БА и семи семей двухлеток БТ, генерации 1981 г. Предложенный метод сравнительной оценки групп по продуктивности работал неизменно успешно.
Эффективность апостериорной минимизации модифицирующих эффектов условий выращивания испытана и в существенно иной в генетическом и селекционном отношении ситуации. В соответствии со схемой селекционного процесса (табл. 3) сравнению подлежали 5 групп рыб: местный карп (зеркальный или чешуйчатый) и его гибриды с интродуцированными породами с первого по четвертое скрещивание.
Таблица 3 - Схема синтетической селекции путем межпородных скрещиваний местного карпа с интродуцированными породами
| Номер скрещивания | Компоненты скрещивания | Межпородный гибрид | Год постановки скрещивания |
| Первое | М х У | МУ | 1981 |
| Второе | МУ х И | МУИ | 1985 |
| Третье | МУИ х У | МУИУ | 1989 |
| Четвертое | МУИУ х М | МУИУМ | 1993 |
| Примечание. Здесь и далее М – местный карп; У – украинский; И – интродуцированные породы (румынский или немецкий карп). | |||
Совершенно очевидно, что такой материал создавал наиболее благоприятные условия для оценки именно генетической мощности системного анализа фенотипической изменчивости комплекса признаков – компонент продуктивности. Достаточно вспомнить об определении наследственности К. Пирсоном как «регрессии степени сходства на степень родства».
Оценка продуктивных качеств осуществлялась на втором году жизни при достижении группами рыб товарного возраста. Каждую из сравниваемых групп выращивали в нескольких прудах, причем, чешуйчатые и зеркальные формы каждой группы выращивали в условиях совместной посадки, в одном пруду, что не позволяло вычленить эффект различия чешуйчатой и зеркальной форм.
Анализ данных об изменчивости рыбопродуктивности, массы тела и выживаемости, с единственным фактором – «номер скрещивания» выявил отчетливое доминирование остаточной (между прудами) дисперсии в общей: 82,9%-89,6%. Это сделало очевидной необходимость минимизации остаточной дисперсии.
По аналогии со многими другими биологическими исследованиями и в рыбоводстве разные пруды нередко рассматриваются как повторности опыта. Однако, фактически различия условий разных прудов и их эффекты на продуктивность столь велики, что побудили к предложению не рассматривать группу рыб в конкретном пруду как повторность, а считать ее отдельным экспериментом (Волчков, 1994). При таком подходе цель статистической обработки данных совокупности экспериментов принципиально изменяется и состоит уже не в вычислении среднего по повторностям, а в «снятии» эффектов различных прудов для объективного сравнения групп.
По итогам дискриминантного анализа, в котором первая и вторая функции учли в совокупности 94,5% общей дисперсии, получена следующая матрица парных расстояний Махаланобиса (таб. 4).
Таблица 4 - Матрица парных расстояний Махаланобиса между местным карпом и его гибридами по комплексу признаков продуктивности
| М | МУ | МУИ | МУИУ | МУИУМ | |
| М | 26,90 | 27,88 | 33,35 | 61,48 | |
| МУ | 0,00 | 6,99 | 13,01 | 58,34 | |
| МУИ | 0,00 | 0,00 | 2,00 | 33,16 | |
| МУИУ | 0,00 | 0,00 | 0,11 | 23,11 | |
| МУИУМ | 0,00 | 0,00 | 0,00 | 0,00 | |
| Примечание. Выше главной диагонали матрицы приведены расстояния Махаланобиса, усл. ед.; ниже – вероятность нуль-гипотезы об отсутствии межгрупповых различий. | |||||
Из таблицы следует, что статистически достоверные межгрупповые различия по продуктивности не установлены только в одном единственном парном сравнении групп – тройного и четверного гибридов. Наиболее удаленными оказались местный карп и конечный продукт синтетической селекции - пятерной гибрид (61,48 усл. ед.). В этом можно было бы видеть главное доказательство эффективности проведенной селекции, если бы уже было показано, что комплексно оцененная продуктивность последнего выше, чем у местного карпа и предшествующих гибридных форм. Оценка вкладов учтенных признаков в дискриминантные функции (табл. 5) позволяла на это надеяться, поскольку признаки mk, ft и pr вошли в них со значительно большими вкладами по сравнению m0 и рр эффекты которых минимизировались.
Таблица 5 - Значения стандартизованных коэффициентов дискриминантных функций (вклады признаков в их значения)
| Признак | Дискриминантная функция | Средний взвешенный модуль коэффициента | |
| первая | вторая | ||
| m0 | - 0,15 | 0,20 | 0,08 |
| pp | 0,01 | 0,70 | 0,11 |
| mk | - 1,15 | 0,65 | 0,47 |
| ft | 1,14 | - 0,85 | 0,49 |
| pr | - 0,84 | - 0,28 | 0,31 |
| Примечание. Модуль вклада взвешен на долю общей дисперсии, учтенной каждой дискриминантной функцией. | |||
Оценка расстояний центроидов местного карпа и гибридов с точкой модели это доказало (рис. 4).
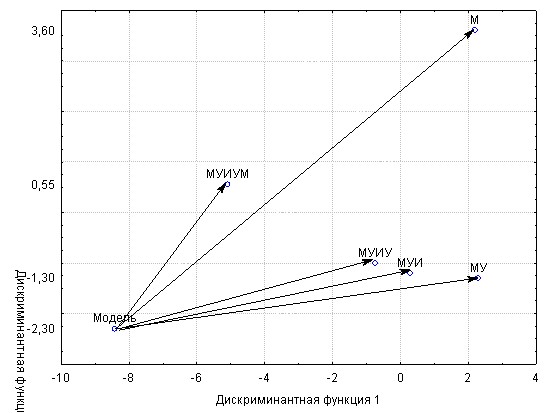
Рис.4 - Распределение центроидов групп местного карпа и его гибридов относительно точки селекционной модели в пространстве дискриминантных функций
Первое важное обстоятельство, ясное из рисунка, заключается в том, что точка модели очевидно выходит за рамки распределения центроидов опытов с местным карпом и его гибридами. Иными словами, ни одна из сопоставляемых групп, как и следовало ожидать, еще не достигает точки селекционной модели. Напомним, что последняя отвечает, в некотором роде, идеальному объекту, которому приписаны наилучшие значения параметров с учетом их взаимных связей. Практическая селекция и не может рассчитывать на полное воплощение идеала в реальном объекте. Причина этому – в хорошо известной «нежелательности» некоторых генетических корреляций признаков (Кирпичников, 1987). С этих позиций селекция может правомерно рассчитывать лишь на максимальное приближение к селекционной модели по комплексу признаков.
Главный же результат состоит в монотонном сокращении расстояний до точки модели в ряду поколений синтетической селекции: М – 12,6 усл. ед; МУ – 11,1 усл. ед; МУИ – 9,2 усл. ед; МУИУ – 8,2 усл. ед; МУИУМ – 5.4 усл. ед. Достоверность различия средних большинства расстояний подтверждена в дисперсионном анализе (p<0,05).
Глава 4. Морфометрический анализ в решении задач селекции рыб
Морфологические признаки несут в себе богатейшую информацию об организме в целом, - так обобщил опыт работы биологов Н.А. Заренков (1976) в «Лекциях по теории систематики». Очевидный успех использования этой категории признаков при построении классификаций уже давно привлек внимание специалистов в области популяционной биологии, микроэволюции, а затем и селекции.
В селекции рыб первая из функций системного морфометрического анализа может состоять в оперативной оценке степени генетических различий между группами (например, потомствами от индивидуальных скрещиваний в семейной селекции) или фиксации сдвига в генетической структуре материала в результате селекционных мероприятий (например, скрещиваний на этапах синтетической селекции). При наличии такой возможности число групп, вовлекаемых далее в трудоемкие эксперименты по сравнительной оценке продуктивности, может быть разумно ограничено.
В описанных ниже опытах изучаемой системой был «морфотип рыбы» - комплекс промеров тела или его частей, измеренных в соответствии с рекомендациями И.Ф. Правдина (1966). Системность анализа изменчивости обеспечивалась использованием адекватных многомерных статистических методов, прежде всего дискриминантного анализа.
В общей сложности с целью оценки морфометрических различий семей от индивидуальных скрещиваний в работе выполнены и проанализированы три масштабных эксперимента. Морфометрически сопоставлены: 5 семей сеголеток БА, выращенных раздельно в прудах КЗРП и КрасНИИРХа; 5 семей годовиков БТ, выращенных в прудах рыбхоза «Горячий Ключ», и 4 семьи двухлеток БТ, выращенных совместно в двух прудах КЗРП.
При работе с комплексом признаков возможно те признаки из числа учтенных, которые, в первую очередь, способствуют разделению групп. Далее такой сокращенный комплекс признаков называется информативным. Определение информативного комплекса обеспечивается специальной процедурой пошагового дискриминантного анализа. Так, например, при сравнении семей двухлеток БТ из 20 первоначально учтенных морфометрических признаков в информативный комплекс вошли только четыре: постдорсальное расстояние (pD), длина головы (С), наименьшая высота тела (h) и ширина лба (io). Возможность не снизить разрешающую способность метода при сокращении числа измеряемых признаков показана во всех экспериментах по морфометрическому сравнению семей. Критерием сохранности разрешающей способности служила частота корректных отнесений особей к «своей» группе.
Сравнение информативных комплексов, выявленных при обработке данных различных экспериментов, указало на важное обстоятельство. Список признаков, входящий в такой комплекс, отнюдь не универсален и изменяется не только от вида к виду, но и в пределах одного вида от возраста рыб, условий их выращивания и состава сопоставляемых групп. По первому, но ошибочному, впечатлению это может породить сомнения в пользе определения такого списка. Несомненно, что состав информативного комплекса однозначно связан со спецификой объектов и условиями их выращивания. Другими словами, список информативен только в рамках конкретного сравнительного эксперимента. Однако целесообразность его определения этим не умаляется. Этот вывод становится очевидным с позиций традиционного в дискриминантном анализе понятия «обучающей выборки». Выполненные на такой выборке детальные морфометрические исследования, в итоге которых и определяется состав сокращенного комплекса, позволяют многократно снизить затраты труда на морфометрическую характеристику остальных особей и целых групп, вовлеченных в конкретный эксперимент. Уместно подчеркнуть, что время, затраченное на производство измерений, определяет основные трудозатраты сравнительного эксперимента. Время, потраченное на подготовку электронных файлов данных и последующие статистические процедуры, настолько мало, что им можно пренебречь.
Опыт сравнительного изучения пяти семей БТ в возрасте годовиков занимает особое место в структуре морфометрических исследований, выполненных в данной работе. Относительно небольшие выборки рыб (17-19 особей) из каждой семьи весной были подвергнуты морфометрическому описанию по комплексу из 21 признака. Те же семьи, выращенные в трехкратной повторности раздельной посадки, осенью того же года были оценены по комплексу признаков – компонент продуктивности с использованием всех описанных ранее процедур по апостериорной минимизации эффекта различий условий пруда. Тем самым открылась возможность сопоставить результаты оценки морфометрических межсемейных расстояний с оценкой различий семей по продуктивности на одном и том же экспериментальном материале.
Матрицы парных межсемейных расстояний Махаланобиса, определенных по морфотипу и продуктивности (табл. 6), свидетельствует об их хорошем соответствии.
Таблица 6 – Матрица парных расстояний Махаланобиса между семьями, определенных по морфотипу и компонентам продуктивности
| Семья | A5 | B7 | B3 | A4 | B8 |
| A5 | 23,6 (6) | 21,1 (5) | 11,5 (3) | 45,7 (10) | |
| B7 | 6,2 (2) | 3,7 (1) | 11,3 (2) | 37,5 (9) | |
| B3 | 21,7 (6) | 12,7 (4) | 15,5 (4) | 33,0 (7) | |
| A4 | 9,7 (3) | 4,3 (1) | 17,6 (5) | 35,2 (8) | |
| B8 | 65,5 (8) | 63,9 (7) | 76,1 (10) | 76,0 (9) | |
| Примечание. Выше главной диагонали приведены расстояния определенные по морфотипу, ниже – по продуктивности. В скобках указан ранг расстояния. | |||||
Коэффициент корреляции рангов Спирмена оказался статистически достоверным и достиг величины 0,72 (р<0,05), позволяющей отнести связь разных типов расстояний к категории сильных.
Итог оценки корреляции двух категорий межсемейных расстояний выявил практически значимый аспект морфометрических исследований в семейной селекции. Когда полученная совокупность семей подразделена на группы морфометрически сходных, нецелесообразно включать в последующую оценку по продуктивным качествам все полученные семьи. Генетически и технологически оправданно и разумно ограничить список оцениваемых семей теми представителями морфометрически разных групп, между которыми можно ожидать различий по селекционно значимым признакам. Роль предварительной оценки генетического различия семей и выполняет сравнительный морфометрический анализ на первом году выращивания потомств от индивидуальных скрещиваний.
Когда возможности морфометрической экспресс – оценки генетически обусловленных различий между семьями уже показаны, были основания полагать, что этот анализ может сыграть ту же роль в мониторинге процесса синтетической селекции, представляющей собой ни что иное, как целенаправленное поэтапное преобразование генетической структуры селекционируемого материала.
Морфометрически достоверно различными оказались гибриды, полученные на всех этапах синтетической селекции. Именно это обстоятельство позволяет говорить о возможности мониторинга эффекта селекционных мероприятий. О мероприятиях пишем потому, что скрещивания постоянно сопровождались массовым отбором. Итоги заключительного этапа (МзУзНзУз х Мз) отражены в табл. 7.
Таблица 7 - Расстояния Махалонобиса между центроидами групп Мз, МзУзНзУз и МзУзНзУзМз (усл. ед.) и оценка их достоверности
| Группа рыб | МзУзНзУз | МзУзНзУзМз | Мз |
| МзУзНзУз | - | 6,3 | 21,3 |
| МзУзНзУзМз | 0,00 | - | 27,3 |
| Мз | 0,00 | 0,00 | - |
| Примечание. Мз – местный зеркальный карп; МзУзНзУз – четверной гибрид местного, украинского и немецкого карпов; МзУзНзУзМз – пятерной гибрид. | |||
Видно, что расстояния между четверным и пятерным гибридами на порядок меньше в сравнении с их расстояниями от исходной формы – местного карпа. Последнее скрещивание сложного гибрида с местным карпом имело целью усилить в конечном продукте селекции высокий адаптационный потенциал местной формы к специфическим условиям существования в Приморско-Ахтарском районе Краснодарского края (высокая минерализация воды, наличие очага краснухи).
Одна из прикладных задач работы состояла в оптимизации методов распознавания пород и породных групп, претендующих на селекционные достижения, на основе анализа изменчивости комплекса коррелированных признаков. Проблема различимости селекционных достижений в рыбоводстве всегда стояла достаточно остро в связи с очевидным недостатком у рыб «хороших морфологических маркеров». Последнее обстоятельство единодушно признавалось специалистами в области селекции и племенной работы (Кирпичников 1979; Слуцкий 1978).
Предлагаемое решение демонстрируется здесь на примере морфометрического разделения 4 пород форели (рис. 5).

Рис. 5 - Ординация особей четырех пород форели в пространстве двух дискриминантных функций
Из рисунка следует, что четыре породы действительно эффективно разделяются в пространстве двух дискриминантных функций. Первая из них дифференцирует породы Камлоопс и Адлер, с одной стороны, и форель Дональдсона и Стальноголового лосося – с другой. Вторая разделяет, соответственно, эти пары: Камлоопс и Стальноголового лосося и Адлер и Долнальдсона.
Сокращение полного списка из 20 морфометрических признаков до информативного из 10 не снизило качества дискриминации (табл. 7).
В группу информативных признаков вошли: отношения к длине тела по Смиту постдорсального расстояния, наибольшей высоты тела, антевентрального расстояния, высоты анального плавника, наименьшей высоты тела, длины основания анального плавника, а также отношения к длине головы ширины лба, длины рыла, горизонтального размера глаза, высоты головы у затылка.
При разделении пород только на информативном комплексе показал, что процент корректных отнесений особей к «своей» породе действительно изменился мало. Соответствующие пары значений этого процента для полного и информативного комплекса составили для форели Камлоопс 95,2% и 93,5%; для форели Дональдсона – 83,3% и 80,0%; для форели Адлер – 88,7% и 82,3%; для Стальноголового лосося – 93,5% и 93,6%.
Таблица 7 - Расстояния Махаланобиса между центроидами пород полученные на полном и информативном комплексе индексов
| Порода | Камлоопс | Дональдсон | Адлер | Стальноголовый |
| Камлоопс | – | 11,80 (3) | 6,81 (5) | 16,44 (2) |
| Дональдсон | 13,90 (3) | – | 11,50 (4) | 5,12 (6) |
| Адлер | 8,26 (5) | 12,39 (4) | – | 23,24 (1) |
| Стальноголовый | 20,05 (2) | 7,20 (6) | 26,89 (1) | – |
| Примечание. Выше главной диагонали приведены межгрупповые расстояния Махаланобиса, определенные на информативном списке, ниже – на полном списке индексов. ранги расстояний (в скобках) сохраняются. | ||||
Таким образом, можно заключить, что дискриминантный анализ на основе предварительно выявленного информативного комплекса эффективно решает задачу разделения четырех пород форели.
Глава 5. Генетически детерминированная и контролируемая отбором внутригрупповая изменчивость рыб как причина межгрупповых различий
По мере накопления данных о достоверных различиях по морфотипу между семьями от индивидуальных скрещиваний, разными поколениями гибридов в синтетической селекции или разными породными группами рыб все острее вставал вопрос о генетической, а точнее популяционно-генетической природе этих различий.
При сравнении групп оценка внутригрупповой изменчивости, например, внутрисемейной выполняет единственную функцию. На ее фоне оценивается достоверность различия групповых средних. Сама внутрисемейная изменчивость при этом анализу не подлежит, хотя в структуре изменчивости комплекса признаков она неизменно доминирует. Возникает опасение, что упускается, может быть, главное.
Необходимость детального исследования внутригрупповой изменчивости для понимания природы изменчивости межгрупповой в данной работе ранее всего стала ясной в результате анализа экспериментов по сравнительному морфометрическому исследованию семей белого амура, выращенных в условиях двух различных хозяйств (КрасНИИРХ и КЗРП). Три важные обстоятельства определили выбор именно этого эксперимента. Во первых, число сопоставляемых семей было наибольшим – шесть. Во вторых, семьи были получены в системе скрещиваний топ-кросс (три самки х два самца; семьи: 10.13; 10.21; 14.13; 14.21; 18.13; 18.21), что позволяло рассчитывать на сходство, по крайней мере, части морфометрических вариантов рыб, возникающих в потомстве от разных индивидуальных скрещиваний. В третьих, различия условий выращивания рыб в прудах двух хозяйств были столь значительны, что были способны определить различные векторы естественного отбора.
О последнем свидетельствовали, прежде всего, результаты дисперсионного анализа результаты дисперсионного анализа изменчивости массы тела рыб. Вклады различий «между семьями» и «между хозяйствами» оказались практически одинаковыми (22,5% и 26,7%, соответственно). Однако значительным оказался и вклад взаимодействия факторов «семья – хозяйство» (36,6%). Он означает, что семьи в разных хозяйствах росли неодинаково, специфически реагируя на изменение условий выращивания. Корреляция рангов семей по средней массе тела в КрасНИИРХ и КЗРП была недостоверной (rs= 0,67; p>0,05).
Результат наилучшим образом объясняется в рамках наиболее вероятного предположения о разной адаптивности неких элементов структуры семей в условиях разных хозяйств и нацеливает на решение трех кардинально важных вопросов: как можно выделить эти элементы и что они собой представляют; различны ли частоты этих элементов в разных семьях изначально по причине происхождения от разных производителей и, наконец, изменяются ли эти частоты с изменением условий выращивания, то есть можно ли связать их изменения с отбором.
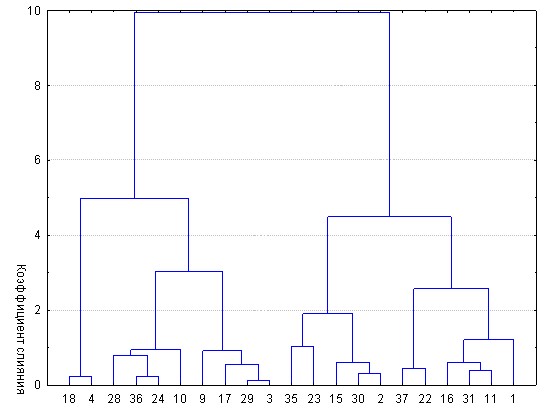
Основную роль в выявлении внутрисемейной изменчивости морфотипа сыграл кластерный анализ. Единицей кластеризации выступала особь, характеризованная по комплексу коррелированных морфометрических признаков.
На первом этапе исследовались выборки рыб каждого из 12 опытов (шесть семей х два хозяйства). Результаты кластеризации семьи 10.13, выращенной в условиях КрасНИИРХ отражены в качестве примера на рис. 6.

Рис. 6 – Результат кластеризации сеголеток белого амура семьи 10.13., выращенных в условиях КрасНИИРХ
Из рисунка следует, что выборка рыб семьи 10.13, выращенная в КрасНИИРХ, разделяется на 4 кластера численностью 3, 13, 9 и 7 рыб (порядок кластеров справа налево). В сумме по шести семьям и двум хозяйствам выделено 40 кластеров, далее и полежавших сравнительному исследованию.
Прежде всего, положительно решен вопрос о гомологичности кластеров рыб, выявленных в одной семье, но различных хозяйствах. Способ решения состоял в следующем. Для каждого из выявленных кластеров вычислены средние значения. Они и послужили основой для определения меры сходства разных кластеров на втором этапе анализа. В качестве примера приведены результаты оценки гомологии кластеров в семье 14.13. (рис. 7).
Приведенный рисунок убедительно свидетельствуют о том, что кластеры, выделяемые в пределах семьи в прудах одного хозяйства, как правило, имеют своих гомологов среди рыб, выращенных в другом. Только один кластер (№ 18 из КрасНИИРХ) не нашел «своей пары».
Крайне существенно, что гомологичные кластеры рыб обнаруживаются не только в пределах одной семьи при выращивании в разных хозяйствах, но и в пределах различных семей при выращивании в одном (рис.8).
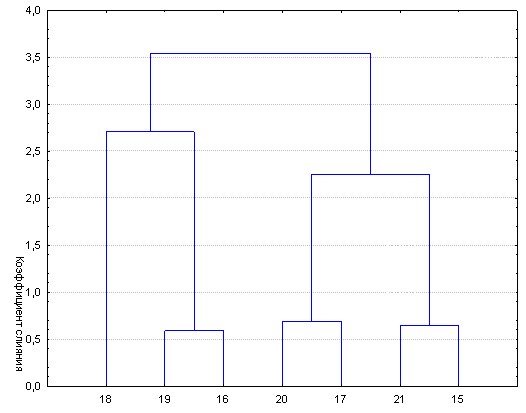
Рис. 7 – Результат группировки кластеров, первоначально выделенных в семье 14.13
Кластеры 15-18 выделены при выращивании в КрасНИИРХ; 19-21 – в КЗРП
Шесть первоначально найденных кластеров: 1, 11, 31, 16, 22, 37, - входящие в I группу, все принадлежат к разным семьям. То же самое можно сказать относительно II группы кластеров (2, 30, 15, 23, 35). Число кластеров, сформировавших III группу, равное восьми, больше числа сопоставляемых семей, поэтому естественны повторы. Их два: кластеры 9 и 10 оба принадлежат к семье 10.21, а 28 и 29 – к семье 18.13. Однако следует обратить внимание на то, что при попытке разрезании дендрита по более низкому уровню, например в 1,5 усл. ед., они оказались бы в разных группах. Кластеры 4 и 8, образовавшие IV группу, принадлежат к семьям 10.13 и 18.13.
Группировка кластеров, первоначально выделенных в семейных выборках из КЗРП, привела к аналогичному результату. Выделено также 4 группы кластеров (обозначены V-VIII), численностью 4, 6, 4 и 5 кластеров.
Гомологичность многих из первоначально выделенных 40 кластеров свидетельствует о том, что изменчивость морфотипа рыб в изученном материале в действительности не так уж и широка. Особенно наглядно это проявляется при попытке описать эту изменчивость геометрически: в виде распределения центроидов разных кластеров в пространстве, дискриминирующем их группы. Судя по рис. 9, распределение центроидов 21 кластера, найденных в семейных выборках из КрасНИИРХ, образует 4 неперекрывающихся облака точек. Они и есть те 4 группы кластеров, которые выделены на втором этапе анализа.
IV III II I
Рис. 8 – Группировка кластеров рыб, выявленных в 6 семьях белого амура, выращенных в прудах КрасНИИРХ
(I – IV – группы кластеров)
Семья 10.13 (кластеры 1-4); семья 10.21 (9-11); семья 14.13 (15-18); семья 14.21 (22-24); семья 18.13 (28-31); семья 18.21 (35-37).
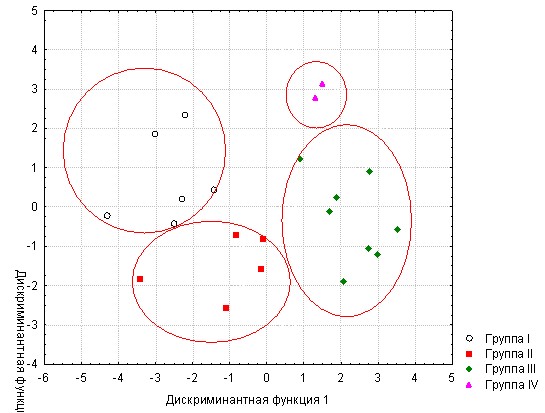
Рис. 9 – Распределение центроидов кластеров, выявленных в семейных ыборках из КрасНИИРХ в пространстве дискриминантных функций
Становится очевидным, что в шести семьях от индивидуальных скрещиваний, выращенных в КрасНИИРХ, встречаются одни и те же четыре различных морфометрических класса рыб, но с различной частотой. В популяционной биологии уже есть термин, обозначающий это явление – «морфизм» (Huxley, 1955, цит по Ригер, Михаэлис, 1967). Это постоянное или временное сосуществование в пределах популяции двух или более генетически различных форм - морф. Так далее в работе и будут именоваться выявляемые в системном анализе изменчивости комплекса морфометрических признаков варианты морфотипа рыб.
Заранее очевидно, что основа их различия не может быть никакой иной, кроме как генетической. Тем не менее, наиболее важным аргументам в пользу генетических различий морф ниже в работе отведено специальное место.
Те же данные по шести семьям из КрасНИИРХ, представленные в виде таблицы двумерного распределения (табл. 8), позволяют количественно оценить межсемейные различия по относительной частоте встречаемости морф.
Таблица 8 – Двумерное распределение частот сеголетков белого амура по принадлежности к семье и группе кластеров в условиях КрасНИИРХ
| Семья | Номер группы кластеров | t-критерий | |||
| I | II | III | IV | ||
| 10.13 | 3 (9,4) | 13 (40,6) | 9 (28,1) | 7 (21,9) | 1,05 |
| 10.21 | 8 (25,8) | 0 (0,0) | 23 (74,2) | 0 (0,0) | 3,97 * |
| 14.13 | 7 (23,3) | 3 (10,0) | 13 (43,3) | 7 (23,3) | 1,66 |
| 14.21 | 4 (15,4) | 4 (15,4) | 18 (69,2) | 0 (0,0) | 4,18 * |
| 18.13 | 4 (13,8) | 7 (24,1) | 18 (62,1) | 0 (0,0) | 3,00 * |
| 18.21 | 6 (20,0) | 6 (20,0) | 18 (60,0) | 0 (0,0) | 3,27 * |
| Сумма | 32 (18,0) | 33 (18,5) | 99 (55,6) | 14 (7,9) | 7,48* |
| Примечания. 1). 2=52,8 (р=0,00); К=0,28; 2).здесь и далее в скобках после абсолютной численности класса приведена его частота, %; 3). t-критерий вычислен по результатам сравнения наиболее частой в семье морфы со следующей за ней по частоте. | |||||
Вычисленное по данным таблицы фактическое значение Хи-квадрат превысило стандартное для соответствующего числа степеней свободы. Сила связи «семья – частота морфы», оцененная по показателю взаимной сопряженности признаков (Чупрова) определена как средняя: К=0,28.
Детально различия распределений морф в разных семьях проявляются при сравнении их частот (% от объема семьи). Сравнение проведено с использованием преобразования по Фишеру (![]() ). В четырех семьях из шести выявлена морфа, обладающая максимальной в сравнении с другими частотой – модальная. Во всех случаях это морфа III.
). В четырех семьях из шести выявлена морфа, обладающая максимальной в сравнении с другими частотой – модальная. Во всех случаях это морфа III.
Аналогичный в ряде отношений результат получен при группировке кластеров рыб, выращенных в условиях КЗРП (2=62,2 р=0,00; К=0,32). Однако есть и существенное отличие. Если в условиях КрасНИИРХ модальная морфа, по-видимому, наиболее адаптивная определилась в большинстве семей уже на первом году выращивания, то в условиях КЗРП этого не произошло. Модальная морфа найдена лишь в одной семье (18.21).
Напомним, что условия выращивания рыб в прудах КрасНИИРХ были более благоприятными в сравнении с КЗРП. С этих позиций и можно, с нашей точки зрения, объяснить полученный результат. Благоприятные условия выращивания в КрасНИИРХ обеспечивают реализацию генотипически обусловленных различий морф по адаптивности. Суммарные по данному хозяйству частоты четырех морф: 18,0%, 18,5%, 55,6% и 7,9%, - варьируют в широких пределах. Варьирование таких же частот четырех морф на КЗРП не столь широко: 24,7%, 33,8%, 21,4% и 20,1%. По-видимому, по той же причине в семейных выборках КЗРП не встречаются морфы столь редкие, как морфа четыре в КрасНИИРХ (7,9%).
Оставалось объединить результаты кластерного анализа рыб, полученные в раздельном анализе выборок из двух хозяйств. Необходимость этого очевидна. Общее число выделенных при раздельном анализе групп кластеров составляет восемь: (I – IV) – в выборках из КрасНИИРХ, (V – VIII) – из КЗРП. Действительное их число должно быть заведомо меньшим, поскольку, как уже было показано, большинство кластеров, выделенных в семейных выборках из КрасНИИРХ, имеют своих гомологов в выборках КЗРП (см. рис. 6,7). Методом решения вопроса о гомологии морф, выделенных в различных хозяйствах, мог быть все тот же кластерный анализ групп. Но интерпретация его результатов столкнулась с некоторыми трудностями: статус группы VIII КЗРП оказался не вполне определенным. Выбор оптимального кластерного решения должен был основываться на каком то ином подходе. В его качестве был избран анализ матрицы парных расстояний Махаланобиса при сравнении всех восьми выявленных групп (табл. 9).
Таблица 9 – Матрица парных расстояний Махаланобиса для восьми групп кластеров рыб, выращенных в двух хозяйствах
| Группа | I | II | III | IV | V | VI | VII | VIII |
| I | 9,70 | 19,16 | 10,49 | 0,84 | 13,59 | 8,13 | 14,54 | |
| II | 0,00 | 6,33 | 22,18 | 15,94 | 2,30 | 0,09 | 6,58 | |
| III | 0,00 | 0,00 | 16,06 | 24,79 | 1,00 | 7,28 | 0,62 | |
| IV | 0,00 | 0,00 | 0,00 | 8,85 | 16,57 | 21,22 | 10,40 | |
| V | 0,38 | 0,00 | 0,00 | 0,00 | 19,36 | 13,97 | 18,77 | |
| VI | 0,00 | 0,04 | 0,20 | 0,00 | 0,00 | 2,89 | 1,39 | |
| VII | 0,00 | 0,92 | 0,00 | 0,00 | 0,00 | 0,04 | 7,14 | |
| VIII | 0,00 | 0,00 | 0,37 | 0,00 | 0,00 | 0,14 | 0,00 |
Приведенные в таблице оценки вероятности нуль-гипотезы об отсутствии различий между группами (вниз от главной диагонали) подсказывают единственно корректное решение разделения групп на морфы – объединении группы VIII КЗРП с группам III КрасНИИРХ и VI КЗРП. Во всех трех парных сравнениях в новой группе достоверных различий не обнаружено (рН0 : 0,14; 0,37; 0,20). Естественно недостоверными оказались различия в парах: VII КЗРП - II КрасНИИРХ (0,92) и V КЗРП – I КрасНИИРХ (0,38). В этом варианте разделения групп выполнялось и другое условие: расстояния между членами разных морф во всех возможных парных сочетаниях должны быть достоверными (p<0,05). Анализ данных таблицы показывает, что в 23 таких парных сравнениях вероятность нуль-гипотезы действительно колебалась в пределах 0,00 – 0,04.
Итогом всей совокупности кластерных анализов и проверки корректности кластерных решений стало заключение о сосуществовании в семьях белого амура четырех различных морф (обозначены: А, В, С, D). Частоты этих морф в выборках из КрасНИИРХ и КЗРП представлены в табл. 10.
Таблица 10 - Двумерное распределение частот частот сеголетков белого амура по принадлежности к морфе и месту выращивания
| Место выращивания | Морфа | |||
| A | B | C | D | |
| КрасНИИРХ | 32 (17,9) | 33 (18,5) | 99 (55,6) | 14 (8,0) |
| КЗРП | 38 (24,7) | 33 (21,4) | 83 (53,9) | 0 (0,0) |
| t-критерий | 0,15 | 0,65 | 0,30 | 5,20 * |
| Примечания. 1). 2=14,3 (р=0,00); К=0,16; 2). в скобках после абсолютной численности класса приведена его частота, % | ||||
Выше показано (см. табл. 8), что распределения частот морф в разных семьях различны. Причина кроется, безусловно, в генетических различиях между соответствующими парами производителей, от которых получены семьи. Данные табл. 10 свидетельствуют о том, что другой причиной различия распределений частот морф могут стать и различия условий выращивания. Другими словами, в формировании морфизма участвует естественный отбор в форме дифференциальной выживаемости разных генотипов.
Выявленная в экспериментах на семьях белого амура зависимость внутригрупповой структуры – (соотношения частот морф) от генотипов родоначальников семей и условий выращивания потомства (естественного отбора) направило дальнейшие исследования в русло проблем популяционной генетики. Однако, совершенно очевидно, что потомства от одних и тех же скрещиваний, выращенные в различных хозяйствах, в течение одного сезона, популяциями не являются. Для корректного решения вопроса о роли внутрипопуляционной изменчивости в формировании межпопуляционных различий был нужен адекватный экспериментальный материал. В целом ряде важных отношений соответствующим требованиям удовлетворяли исследования, выполненные на черноморской кумже.
Материал для исследования получен в результате морфометрического и остеологического (структуры осевого скелета) описания выборок из различных природных и искусственных популяций: рек Пшада и Небуг и двух рыборазводных заводов (Адлерский производственно-экспериментальный рыбоводный лососёвый завод (АПЭРЛЗ) и ФГУП форелевое хозяйство «Адлер».
Изученные выборки и их численность представлены в таблице 11.
Таблица 11 – Структура объединённой выборки черноморской кумжи
| Местообитание | Возраст особей | Численность выборки | Обозначение группы |
| ФХ "Адлер" | Второй год жизни | 85 | Адлер-2 |
| ФХ "Адлер" | Третий год жизни | 35 | Адлер-3 |
| ФХ "Адлер" | Четвёртый год жизни | 30 | Адлер-4 |
| АПЭРЛЗ | Второй год жизни | 100 | АПЭРЛЗ-2 |
| АПЭРЛЗ | Третий год жизни | 25 | АПЭРЛЗ-3 |
| АПЭРЛЗ | Четвёртый год жизни | 29 | АПЭРЛЗ-4 |
| р. Пшада | Разновозрастные | 36 | Пшада |
| р. Небуг | Разновозрастные | 42 | Небуг |
| Всего | 382 | ||
С нашей точки зрения, по всем основным критериям стада рыборазводных заводов отвечают понятию «искусственная популяция». Действительно, в пределах таких стад осуществляется панмиксия в том смысле, что при выборе производителей для искусственного воспроизводства не отдаётся предпочтение каким-либо морфометрическим вариантам рыб. Условия развития и жизни рыб на различных заводах различны. Изоляция стад имеет место. Вполне ожидаемы, особенно при сравнении искусственных и природных популяций, и различия в направлениях отбора. Наиболее важная характеристика популяций заключается в их генетической гетерогенности. Гетерогенны, как будет показано ниже, не только природные популяции кумжи, но и искусственные популяции рыборазводных заводов.
Обратимся непосредственно к итогам дискриминантного анализа. Качество разделения групп оценено по расстояниям Махаланобиса (таблице 12).
Таблица 12 – Расстояния (Махаланобиса) между центроидами дискриминируемых групп
| Группы | АПЭРЛЗ 2 | Адлер-2 | АПЭРЛЗ3 | Адлер3 | АПЭРЛЗ4 | Адлер 4 | Пшада | Небуг |
| АПЭРЛЗ-2 | – | 3,01 | 79,20 | 78,11 | 296,64 | 331,24 | 51,07 | 36,33 |
| Адлер-2 | 0,08 | – | 83,99 | 79,81 | 300,65 | 332,62 | 54,27 | 31,67 |
| АПЭРЛЗ-3 | 0,00 | 0,00 | – | 8,41 | 123,94 | 125,66 | 70,47 | 69,65 |
| Адлер-3 | 0,00 | 0,00 | 0,00 | – | 137,97 | 111,85 | 62,52 | 53,28 |
| АПЭРЛЗ-4 | 0,00 | 0,00 | 0,00 | 0,00 | – | 68,24 | 205,94 | 201,66 |
| Адлер-4 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | – | 201,01 | 228,21 |
| Пшада | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | – | 20,49 |
| Небуг | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | – |
| Примечание: выше главной диагонали указаны расстояния Махаланобиса, ниже – вероятность ноль-гипотезы об отсутствии различий. | ||||||||
Из таблицы следует, что достоверно различаются все сравниваемые группы, кроме выборок из двух маточных стад, исследованных на втором году жизни (3,01; p=0,08).
Далее расстояние увеличилось и стало достоверным к третьему году
(8,41 усл. ед.) и существенно увеличилось четвертому году жизни (68,24 усл. ед.). Таким образом, различные условия выращивания рыб в двух рыборазводных хозяйствах приводят с течением времени к формированию специфического соотношения морфотипов кумжи, что, несомненно, происходит под действием разнонаправленного естественного отбора.
Следует отметить и то, что наибольшим сходством с природными популяциями обладают выборки из рыборазводных хозяйств, описанные на втором году жизни. Так, наименьшие расстояния выборки из
р. Пшада выявлены до групп АПЭРЛЗ-2 (51,07 усл. ед.) и Адлер-2
(54,27 усл. ед.). Далее, в зависимости от возраста групп, эти расстояния увеличились до 70,47 усл. ед. и 60,52 усл. ед. для третьего года жизни и
205,94 усл. ед. и 201,01 усл. ед. – для четвёртого. Та же закономерность выявлена и для выборки из р. Небуг.
Основную задачу всего последующего анализа экспериментальных данных мы видим в проверке положения, по которому все фиксируемые при измерении расстояний Махаланобиса, межпопуляционные различия, связанные с происхождением и, следовательно, генофондом популяций или с различием условий их существования основаны на различии частот элементов внутрипопуляционного разнообразия - морф.
Поиск морф был выполнен по тому же алгоритму, который был использован при анализе внутрисемейной структуры. В пределах каждой из восьми выборок методом кластерного анализа были выделены группы фенотипически сходных особей. Далее среди них были выявлены гомологи и была проведена оценка частоты каждой из морф в разных выборках. Однако, особое место в данном разделе работы было отведено определению биологического статуса выделенных групп.
Проверка кластерного решения проведена с помощью дисперсионного и дискриминантного анализов. Первый позволил оценить межкластерные различия по средним значениям признаков, второй – по их комплексу и, одновременно по системе их корреляций. Данное исследование направлено на доказательство генетически обусловленных различий между выделенными кластерами, которое и может стать решающим основанием для присвоения им статуса морф – элементов внутрипопуляционной гетерогенности.
По результатам дисперсионного анализа установлено, что межкластерные различия выявляются по всем признакам без исключения, их вклад достаточно высок и варьирует от 27,7% до 73,6% от общей дисперсии. Причем для подавляющего числа признаков доля межкластерных различий превышает 60%.
Если различия средних значений признаков не вызывают сомнений, то различия структуры их связей нуждаются в специальном доказательстве. Анализ корреляционных структур ориентирован на выявление в системе связей признаков так называемых корреляционных плеяд – групп тесно коррелированных признаков, отделенных от других таких же групп слабыми или нулевыми связями (Терентьев, 1959). Различный состав корреляционных плеяд и позволяет считать доказанным факт различия корреляционных структур.
Одним из подходов к выявлению плеяд являтся сопоставление распределений вкладов признаков в их линейные комбинации, полученные с использованием метода главных компонент (Андреев, Решетников, 1978; Андреев, 1980).
В качестве примера в рамках данного подхода был проведен анализ корреляционных матриц, полученных для трех кластерах выборки АПЭРЛЗ-2 (рис. 10).

Рис. 10 – Распределение признаков по их вкладам в первую и вторую главные компоненты для первого кластера группы «АПЭРЛЗ-2»
На рисунке обозначены следующие морфометрические признаки: L - длина тела; С - длина головы; HC - высота головы у затылка; LC - длина средней части головы; R - длина рыла; HН - наибольшая высота тела; H - наименьшая высота тела; AD - антедорсальное расстояние; PD - постдорсальное расстояние; AA - антеанальное расстояние; AV -антевентральное расстояние; PV - пектовентральное расстояние; LP - длина грудного плавника; VA - вентроанальное расстояние; LV - длина брюшного плавника; LD - длина спинного плавника; HD - высота спинного плавника; LA - длина анального плавника; HA - высота анального плавника; AC - длина хвостового стебля.
Коэффициенты – вклады, с которыми признаки включаются в главные компоненты, вычисляются на основе анализа корреляционной матрицы. Это дает основание считать, что близкие по значению и знаку вклады отражают принадлежность признаков к одной корреляционной плеяде и, наоборот. На данном распределении отчетливо выделяется пять групп близко расположенных признаков. Первую группу – корреляционную плеяду образуют признаки AA, AD, AV, H и C. Во вторую вошли признаки L, LT, PD, VA, PV, HC, HН, LD. Третья плеяда содержит три признака: LA, LC и LV, а четвертая – R и LP. Последняя, пятая плеяда включает только один признак – AC. Анализ распределения для кластеров два и три позволил выделить по шесть корреляционных плеяд. Количество признаков в них и, самое главное, их разный качественный состав, позволяет считать, что корреляционная структура второго и третьего кластера выборки АПЭРЛЗ-2 отличается от таковой для кластера один.
Таким образом, можно заключить, что выделенные в рамках кластерного анализа группы особей различаются как по средним значениям признаков, так и по их корреляционной структуре. Данный результат в совокупности с доказанными различиями средних значений признаков позволяет интерпретировать межкластерные различия как генотипически обусловленные, а сами кластеры считать морфами.
Таким образом, в совокупности, полученные нами результаты сравнения кластеров находятся в полном соответствии с фундаментальным тезисом И.И. Шмальгаузена (1968), согласно которому признаки фенотипа не пассивный передатчик информации, а активный авторегулятор индивидуального развития. Генотип полностью детерминирует определенные пути – «каналы» развития. Это выражается в конечном числе схем роста, различимых по значениям признаков и системе их корреляций. Внешние факторы, по И.И. Шмальгаузену, не имеют здесь формообразующего значения, исполняя роль освобождающих причин, специфичность ответа на которые определяется унаследованными нормами реакции. С этих позиций решение задачи выявления искомой структуры через анализ комплекса коррелированных признаков кажется не только возможным, но и наиболее естественным.
Рассмотрим частоту трех выделенных морф в различных по происхождению и возрасту выборках черноморской кумжи (таблица 13).
Таблица 13 – Частоты морф, выделенных в пределах изученных групп особей черноморской кумжи
| Группа | Морфа | |||||
| I | II | III | ||||
| Частота | Метка | Частота | Метка | Частота | Метка | |
| АПЭРЛЗ-2 | 37 37,0% | A | 39 39,0% | B | 24 24,0% | C |
| АПЭРЛЗ-3 | 4 16,0% | H | 10 40,0% | I | 11 44,0% | G |
| АПЭРЛЗ-4 | 7 24,1% | N | 7 24,1% | O | 15 51,8% | M |
| Адлер-2 | 21 24,7% | D | 36 42,4% | E | 28 32,9% | F |
| Адлер-3 | 7 20,6% | K | 12 35,3% | L | 15 44,1% | J |
| Адлер-4 | 7 23,3% | Q | 8 26,7% | R | 15 50,0% | P |
| Пшада | 18 50,0% | V | 12 33,3% | T+U | 6 16,7% | S |
Окончание табл. 13
| Небуг | 18 42,9% | Y | 16 38,1% | W+X | 8 19,0% | Z |
| Примечание: ниже абсолютной численности каждого класса двумерного распределения приведена его частота, % | ||||||
Анализ данного двумерного распределения позволил сделать два важных заключения:
– распределения частот трёх морф в группах (выборках из различных популяций) достоверно различаются;
– достоверно различаются и частоты разных морф в пределах одной группы.
Первое из заключений статистически подтверждено с использованием критерия 2. Фактическое его значение – 31,5 превысило стандартное для 5%-го уровня значимости равное 23,7. Этот результат означает, что характер распределения, то есть соотношение частоты трёх морф связан с условиями существования популяции и возрастом рыб.
Второе заключение подтверждают данные таблицы 14, где приведены результаты сравнения частот трёх морф в двух искусственных и двух естественных популяциях. Основанием для объединения данных по двум искусственным популяциям из двух рыбоводных заводов и двух естественных из рек Пшада и Небуг послужили результаты детального анализа динамики частот трёх морф, связанной с возрастом рыб. Детали этого анализа мы опускаем на том основании, что его итоги установили принципиальное сходство динамики в двух различных искусственных популяциях и её отличие от таковой в естественных популяциях.
Таблица 14 – Динамика частот трёх морф в искусственных и естественных популяциях черноморской кумжи, связанная с возрастом рыб
| Тип популяции и возраст рыб | Частота морф, % | ||
| I | II | III | |
| Искусственные, 2-й год жизни | 30,9 | 40,7 | 28,4 |
| Искусственные, 3-й год жизни | 18,3 | 37,7 | 44,0 |
| Искусственные, 4-й год жизни | 23,7 | 25,4 | 50,9 |
| Естественные, разновозрастные | 46,5 | 35,7 | 17,8 |
Из данных таблицы следует, во-первых, что в искусственных популяциях модальная морфа, то есть характеризующаяся максимальной частотой, не устанавливается ни для второго, ни для третьего года жизни рыб. Под действием естественного отбора в этих поколениях идёт процесс преобразования частот морф. Модальная морфа выявляется лишь к четвёртому году жизни - только к этому возрасту её частота становится достоверно более высокой, чем частоты двух других морф. В обоих искусственных популяциях модальной становится морфа III, частота которой (50,9%) достоверно превышает частоту других морф.
В контексте предпринятого обсуждения генетически обусловленной гетерогенности популяций термин модальная морфа означает наиболее адаптивная к данным конкретным условиям существования.
Судя по данным таблицы 14, в естественных популяциях морфа III, очевидно, не является модальной. На это место претендует морфа I, хотя её численное преобладание над морфой II (46,5% против 35,7%) и не доказывается достоверным, безусловно, в силу ограниченности объёма изученной выборки из природных популяций.
В целом выполненный анализ изменчивости комплекса морфометрических признаков в двух искусственных и двух естественных популяциях приводит к трём основным заключениям:
- элементами структуры популяции кумжи, отражающими её гетерогенность по адаптивности, являются морфы, которые и служат точками приложения естественного отбора;
- морфы успешно выявляются в кластерном анализе особей популяции, выполненном на основе евклидовых расстояний в пространстве главных компонент;
- результат отбора состоит в преобразовании относительных частот морф.
С целью усилить аргументацию в пользу генетической детерминированности различий между морфами, выявляемыми в системном анализе фенотипической изменчивости, мы обратились к исследованию меж- и внутрипопуляционной изменчивости некоторых остеологических признаков, конкретно - структуре осевого скелета рыб: соотношению числа позвонков в грудном (G), переходном (P) и хвостовом (Н) отделах позвоночника. Следует отметить в главную контексте данного исследования особенность этой категории признаков. Они не испытывают модификационной изменчивости на постэмбриональных стадиях развития. Следовательно, выявленные различий между выборками разного происхождения или разного возраста можно достаточно уверенно интерпретировать как генотипические.
В выполненных специальных исследованиях кафедры генетики и микробиологии Кубанского госуниверситета (Ганченко, 1989; Тюрин В.В., 2009) было доказано, что различия между генетически различными группами рыб, например семьями от индивидуальных скрещиваний, заключаются не в различии средних значений отдельных параметрам осевого скелета, а частот формул осевого скелета (О:G:Р:H). Описать формулу можно, объединив значения числа позвонков в разных отделах в интегральный показатель – их линейную комбинацию (главную компоненту). Анализ совокупности полученных данных позволил объяснить наблюдаемое фенотипическое разнообразие в семьях расщеплением по двум генам с функцией, регулирующей соотношения: (G+P)/H и G/P.
При остеологическом исследовании 388 особей черноморской кумжи были встречены 86 вариантов формул осевого скелета (см. абсциссу рис.). Количественной характеристикой каждой из этих формул служили значения главных компонент, на основе которых и была проведена кластеризация формул. Например, значение главной компоненты -0,086 усл. ед. соответствует формуле О = 58; G = 25; P = 7; H = 26, а значение -1,788 усл. ед. формуле О = 53; G = 23; P = 6; H = 24.
Кластерное решение (рис. 11) представлялось единственно верным: встреченные формулы относятся к 4 разным кластерам (обозначены А, В, С, D).

Рисунок 11 – Дендрограмма кластеризации формул осевого скелета черноморской кумжи
Абсцисса – варианты структуры осевого скелета
Двумерное распределение особей кумжи по принадлежности к одному из четырёх генотипических классов по структуре осевого скелета (А, В, С, D) и одной из восьми сопоставляемых групп приведено в табл. 15.
Таблица 15 – Частоты генотипических классов по структуре осевого скелета в изучаемых группах черноморской кумжи
| Генотипический класс | АПЭРЛЗ-2 | АПЭРЛЗ-3 | АПЭРЛЗ-4 | Адлер-2 | Адлер-3 | Адлер-4 | Пшада | Небуг |
| A | 30 (30%) | 0 (0%) | 0 (0%) | 38 (45%) | 2 (6%) | 1 (3%) | 3 (8%) | 4 (10%) |
| B | 9 (9%) | 7 (28%) | 5 (17%) | 15 (18%) | 17 (49%) | 9 (30%) | 3 (8%) | 14 (33%) |
| C | 53 (53%) | 14 (56%) | 13 (45%) | 21 (25%) | 11 (31%) | 12 (40%) | 9 (25%) | 12 (29%) |
| D | 8 (8%) | 4 (16%) | 11 (38%) | 11 (13%) | 5 (14%) | 8 (27%) | 27 (58%) | 12 (29%) |
| 2=153,3; (p<0,01); К=0,30 | ||||||||
Полная биологическая интерпретация найденной связи пока затруднена в связи с множественностью факторов, способных ее определить. В контексте обсуждения наиболее важным являются различия распределения морф в искусственных и естественных популяциях (табл. 16). В искусственных модальная морфа D, в естественных – С, что можно связать с действием отбора.
Таблица 16 – Частоты генотипических классов в выборках черноморской кумжи, выращенных в условиях искусственных и природных популяций
| Генотипический класс | Популяция | Сумма | |
| искусственная | природная | ||
| A | 71 (23,4%) | 7 (8,3%) | 78 |
| B | 62 (20,4%) | 17 (20,2%) | 79 |
| C | 124 (40,8%) | 21 (25,0%) | 145 |
| D | 47 (15,5%) | 39 (46,4%) | 86 |
| Сумма | 304 | 84 | 388 |
| 2=40,2; (p<0,01); К=0,24 | |||
Как было уже отмечено, в качестве специальных, дополнительных выполнены исследования на черноморской мидии (Mytilus galloprovincialis Lam.). Их цель – убедиться в возможности расширения сферы применения методов изучения меж- и внутрипопуляционной изменчивости, разработанных на рыбах, и на другие объекты аквакультуры.
Материал для исследования получен в результате изучения выборок черноморской мидии из различных местообитаний. Район сбора охватил восточную часть Чёрного моря от Сочи до Тамани. Всего было исследовано 9 местообитаний. Общий объём проанализированного материала составил 1545 особей. Морфометрическое описание створок раковины было выполнено по разработанной нами методике, включающей измерение 23 показателей (Тюрин и др., 2002).
Анализ внутрипопуляционной структуры был выполнен по обычной схеме: кластеризация особей в пределах каждой выборки, выявление элементов внутрипопуляционного разнообразия – морф и сопоставление их частот с целью определения модальных (Тюрин, 2008).
В результате указанной последовательности процедур для каждого местообитания были получены следующие кластерные решения: в Аше и Озереевке было выделено по четыре, а в Сочи, Магри, Новомихайловском, Геленджике, Утрише, Анапе и Тамани - по три кластера. Общее количество выделенных кластеров равно 29. Практически в каждом местообитании, кроме Анапы, были обнаружены кластеры (один - два), обладающие достоверно более высокой частотой. Этот результат выявляет те морфы, адаптивность которых выше в сравнении с другими элементами внутрипопуляционной структуры в конкретном местообитании.
Была получена количественная оценка гетерогенности мидий в пределах каждого местообитания в виде коэффициента внутрипопуляционного разнообразия по Л.А. Животовскому (1982) (табл. 17).
Таблица 17 - Значения коэффициента внутрипопуляционного разнообразия ()
| Местообитание | Коэффициент () | Ошибка | Число морф | Число особей |
| Сочи | 2,97 | 0,0021 | 3 | 64 |
| Аше | 3,92 | 0,0010 | 4 | 299 |
| Магри | 2,92 | 0,0008 | 3 | 280 |
| Новомихайловка | 2,92 | 0,0009 | 3 | 262 |
| Геленджик | 2,97 | 0,0005 | 3 | 174 |
| Утриш | 2,97 | 0,0024 | 3 | 50 |
| Озереевка | 3,93 | 0,0030 | 4 | 116 |
| Анапа | 2,99 | 0,0001 | 3 | 100 |
| Тамань | 2,93 | 0,0014 | 3 | 200 |
Показатель внутрипопуляционного разнообразия, вычисленный для выборок из каждого местообитания, варьировал от 2,92 до 3,93. Если учесть, что его максимальное значение может быть равно либо 3, либо 4 (в зависимости от числа выделенных морф), уровень генотипической гетерогенности местообитаний должен быть признан очень высоким.
Заключение
Понимание необходимости перейти от исследования изменчивости отдельных признаков к их коррелированным комплексам в этой работе пришло уже при решении первой из поставленных задач: разработке эколого-генетических основ семейной селекции рыб.
Для вскрытия и последующей реализации в селекции гетерогенности исходного материала – маточных стад рыборазводных предприятий – был использован классический генетический метод скрещивания и сравнения потомств индивидуальных производителей. Сразу же сложилась весьма интригующая ситуация: при несомненных генетических различиях потомств от разных пар производителей межсемейные различия по продуктивности (выходу рыбы с единицы площади пруда), равно как и ее компонентам: выживаемости, скорости роста, приросту, конечной массе тела рыб, - не выявлялись даже в дисперсионном анализе. В «лучших» случаях вклад межсемейной дисперсии в общую достигал немногих единиц процента, а нередко вообще не выявлялся как достоверный.
Остаточная дисперсия, обусловленная различием условий прудов, где выращивались потомства, поглощала основную долю изменчивости (80% и более). Даже совместное, в одном пруду, выращивание разных семей не решало проблемы в силу естественного взаимодействия «семья – пруд». Попытки разделить семьи по продуктивности за счет увеличения числа повторностей опыта (прудов) изначально представлялись несостоятельными в статистическом, а, главное, технологическом отношении. Требовался принципиальной иной способ анализа данных.
Им стал анализ системы «зарыбленный пруд». Система характеризовалась по комплексу из 8 характеристик, отражающих одновременно биологические особенности выращиваемых групп рыб и их реакцию на изменения рыбоводных условий выращивания. Все 8 характеризующих систему параметров естественно коррелировали. Однако факторный анализ выделил в них три плеяды, отделенные друг от друга относительно слабыми связями. Плеяду «массонакопления» составили: составляют скорость роста, прирост и конечная масса тела рыб. Плеяду «качества посадочного материала»: начальная масса тела и выживаемость. Плеяду «условий выращивания»: плотность посадки, плотность по выходу и рыбопродуктивность. Относительная независимость варьирования показателей трех плеяд позволила сконструировать новые, интегральные характеристики семей за счет дискриминантного анализа групп с одинаковой семейной принадлежностью. Различия «между прудами» при такой организации материала оказывались внутригрупповыми и апостериорно и, главное, эффективно минимизировались.
Селекционная информативность дискриминантных функций гарантировалась тем, что показатели плеяды массонакопления вошли в них с вкладами на порядок более высокими, чем показатели двух других плеяд. Выбор лучшей по продуктивности семьи был построен центроидов семей, четко разграниченных в пространстве дискриминантных функций, с точкой «модели» - некоторым идеальным в условиях конкретного эксперимента объектом.
Обращение к изменчивости морфотипа рыб как в меж-, так и внутрипопуляционном аспектах уже не оставляло выбора основного принципа анализа экспериментальных данных. Он мог состоять только в исследовании изменчивости комплекса коррелированных признаков, поскольку сам морфотип был изначально определен как система взаимосвязанных промеров тела рыбы и его частей.
Возможность группировки (кластеризации) особей по комплексу традиционно измеряемых признаков в выборках каждого из сравнительных экспериментов; последующее выявление гомологичных кластеров в других выборках, например, из разных семей или хозяйств где они выращивались, дали основания рассматривать внутри- и межпопуляционную изменчивость как проявление морфизма, формируемого на базе генетической гетерогенности и регулируемого естественным отбором.
Автором термина «морфизм» (Huxley, 1955) предусмотрен и такой его тип как «непрерывный», когда различия между морфами, определяемые многими генами, размыты, и могут быть выявлены только с помощью специальных методов. О нем в данном случае и идет речь.
Итог морфометрических исследований на полученном материале, с нашей точки зрения, позволил вплотную приблизиться к определению «точек приложения» (по Н.В. Глотову, 1988) естественного отбора. Возникает прямая ассоциация с экоэлементами Е.Н. Синской (1948) – первичными групповыми образованиями в пределах популяции, с которыми оперирует естественный отбор. И, добавим, может, по видимому, оперировать искусственный. Согласно определению Л. Бербанка (1955) селекция- это область промышленности, где знание рабочих методов природы перестает быть просто теорией, а становится подлинным фактором деятельности.
Решение ряда весьма различных в теоретическом и практическом планах вопросов из области эколого-генетического изучения селекционируемых и естественных популяций рыб с единых позиций анализа изменчивости комплекса количественных признаков показалось нам достаточным для включение в заглавие работы термина «методология», которым традиционно называют учение о принципах построения, формы и способах научного познания.
Основные выводы
- Классический метод раскрытия генетической гетерогенности в исходном материале и последующей ее реализации в селекции, который состоит в разложении исходного материала на потомства от индивидуальных скрещиваний, в семейной селекции рыб при прудовом их выращивании практически не реализуем. Причина – абсолютное преобладание изменчивости, связанной с различий условий прудов, на фоне которой выбор лучших семей по продуктивности неосуществим. Разработка метода апостериорного (после опыта) «снятия» модифицирующих условий среды становиться непременным условием решения проблемы.
- Все показатели системы зарыбленного пруда, в совокупности характеризующие как объект – рыбу, так и условия его выращивания, сплетены в единый коррелированный комплекс. Однако факторный анализ выделяет в их составе три корреляционные плеяды, отделенные друг от друга менее сильными связями. По составу вошедших в них показателей они названы: плеядой «массонакопления» (средняя конечная масса тела рыб, скорость роста, прирост за сезон); плеядой «качества посадочного материала» (средняя начальная масса, выживаемость) и плеядой «условий выращивания» (плотность посадки, плотность по выходу, рыбопродуктивность).
- Апостериорная минимизация становится возможной при вовлечении групп, сформированных по семейной принадлежности в дискриминантный анализ, минимизирующий ставшую теперь внутригрупповой изменчивость по показателям плеяд «качества посадочного материал» и «условий выращивания». Селекционная информативность получаемых дискриминатных функций обеспечивается тем, что показатели плеяды «массонакопления» входят в них с вкладами на порядок более высокими, чем показатели двух других плеяд.
- Доказательство различия семей в дискриминантном пространстве еще не решает проблему выбора лучших, поскольку значения дискриминантных функций (усл. ед.) не могут быть переведены в масштаб селекционно значимых признаков. Предложен простой способ решения проблемы: введение в пространство дискриминантных функций, где распределены семьи, некоторой точки – модели, координаты которой вычисляются по максимальным в данном эксперименте значениям показателей массонакопления и средним значениям прочих показателей. Лучшие семьи выбираются по наименьшим расстояниям до модели. Данный подход был эффективен не только при сравнении семей растительноядных рыб, но и пород карпа и гибридов синтетической селекции.
- Доказана возможность использования системного морфометрического анализа в качестве оперативной оценки степени генетических различий между группами (семьями, гибридами, получаемыми на этапах синтетической селекции). Тем самым открывается возможность сокращения числа групп, вовлекаемых далее в трудоемкие эксперименты по сравнительной оценке продуктивности.
- В сравнительных экспериментах показано, что полный список признаков, традиционно рекомендуемых для описания рыб, может быть весьма существенно сокращен без потери качества разделения групп в пошаговом дискриминантном анализе. Хотя получаемый «информативный» список однозначно связан со спецификой конкретного эксперимента, целесообразность его определения не умаляется. Особенно при значительном количестве сопоставляемых групп рыб и больших выборок.
- Установлена высокая корреляция (более 0,7) морфометрических межсемейных расстояний и их расстояний по продуктивности. Поэтому генетически и технологически оправдано ограничить число оцениваемых семей теми представителями морфометрически разных групп, между которыми можно ожидать различий и по селекционно значимым признакам.
- Кластерный анализ вскрывает генетическую гетерогенность групп рыб изученных в работе категорий: семей растительноядных рыб, искусственных и естественных популяций черноморской кумжи. В каждой группе с его помощью выделяется небольшое число (3-5) вариантов морфотипа. Как правило, эти варианты имеют своих гомологов в семьях, происходящих от других производителей. Особенно важно наличие одних и тех же вариантов морфотипа в разных искусственных и природных популяциях. Вся совокупность подобных данных позволяет классифицировать этот феномен как морфизм – сбалансированное сосуществование различных генетических морф. Межсемейные или межпопуляционные различия с этих позиций естественно связывать с относительными частотами морф. Различия частот морф в группах, выращиваемых в разных условиях свидетельствует об участии естественного отбора в формировании морфизма.
Список опубликованных работ по теме диссертации
I. Статьи в изданиях, в которых рекомендуется публикация основных результатов диссертации на соискание ученой степени доктора наук
1. Тюрин В.В., Алехин А.С., Волчков Ю.А. Эколого-генетический аспект проблемы восполнения численности природных популяций черноморского лосося за счет заводского воспроизводства // Проблемы региональной экологии. 2007. №3. – С. 53-56.
2. Тюрин В.В., Алехин А.С., Зубарев А.Н., Волчков Ю.А. Различия природных и искусственных популяций черноморского лосося по комплексу морфометрических признаков // Вестник Российской академии сельскохозяйственных наук. 2007. №6 – С. 74-76.
3. Тюрин В.В., Волчков Ю.А. Динамика генетической структуры семей рыб и проблема выбора оптимального возраста их селекционной оценки // Вестник Российского государственного университета им. Канта. 2008. №7 – С. 43-48.
4. Тюрин В.В. Экоэлементная структура популяций черноморской мидии (Mytilus galloprovincialis Lam.) восточной части Черного моря // Проблемы региональной экологии. 2008. №3 – С. 62-67.
5. Тюрин В.В. Принцип апостериорной минимизации эффекта модификационной изменчивости на примере оценки продуктивности в семейной селекции белого толстолобика (Hypophtalmicthis molitrics. Val.) // Труды Кубанского государственного аграрного университета. 2008. № 5 (14) – С. 118-121.
6. Тюрин В.В. Система генетического контроля структуры осевого скелета рыб и перспективы использования этого признака в популяционно-генетических исследованиях // Известия высших учебных заведений Северо-Кавказский регион. Естественные науки. №1. 2009 – С. 79-83.
7. Тюрин В.В., Бурло А.М. Динамика продуктивности в ряду поколений синтетической селекции местного карпа с использованием интродуцированных пород // Труды Кубанского государственного аграрного университета. 2009 № 1 (16) – С. 93-98.
8. Тюрин В.В., Волчков Ю.А. Определение минимального числа признаков, разделяющих генетически различные группы рыб в системном морфометрическом анализе // Труды Кубанского государственного аграрного университета. 2009. № 5 (20) – С. 132-136.
9. Тюрин В.В., Алехин А.С., Москул Г.А., Волчков Ю.А., Принцип и метод оптимизации генотипической структуры потомства от искусственного воспроизводства рыб // Труды Кубанского государственного аграрного университета. 2010. № 2 (23) – С.. 112-117.
II. Статьи в аналитических сборниках и материалах конференций
10. Сержант Л.А., Хромов Л.В., Тюрин В.В., Краснянский С.Ф., Стецко В.Г. Усовершенствование методов племенной работы с карпом и разработка основ селекции белого толстолобика и канального сома в условиях Северного Кавказа (депонированный отчет о НИР). Отчет о НИР, 1985, № гос. регистрации 01840029496. 120 с.
11. Ганченко И.В., Шарт Л.А., Тюрин В.В. Производственная проверка рыбохозяйственных качеств краснухоустойчивого карпа // Сб. науч. трудов ВНИИПРХ. 1986. № 48 С 23 – 27.
12. Тюрин В.В., Радецкий В.П., Волчков Ю.А. Изменчивость комплекса признаков продуктивности в семьях растительноядных рыб // Рыбохозяйственное освоение растительноядных рыб: Сб. докл. 11 Всесоюзного совещания. Кишинев, 1988. С. 34-36.
13. Тюрин В.В., Синченко С.Л., Радецкий В.П., Волчков Ю.А. Разработка генетических основ селекции растительноядных рыб в прудовых хозяйствах Кубани // Актуальные вопросы изучения экосистемы бассейна Кубани: Сб. докл. Региональной конференции. Краснодар: Кубанский гос. ун-т, 1988. С. 43-46.
14. Решетников С.И., Радецктй В.П., Тюрин В.В., Ганченко М.В. Устойчивость растительноядных рыб к дефициту кислорода как селекционный признак // Актуальные вопросы экологии и охраны природы Азовского моря и Восточного Приазовья: Сб. докл. Региональной конференции. Краснодар: Кубанский гос. ун-т, 1989. С.139-140
15. Тюрин В.В., Волчков Ю.А., Ганченко М.В. Селекционная оценка продуктивности семей краснухоустойчивого карпа Актуальные вопросы экологии и охраны природы Азовского моря и Восточного Приазовья: Сб. докл. Региональной конференции. Краснодар: Кубанский гос. ун-т, 1989. С.141-142
16. Тюрин В.В. Радецкий В.П., Волчков Ю.А. Системный подход к оценке продуктивности пород рыб // Актуальные вопросы экологии и охраны природы экосистем малых рек: Сб. докл. Региональной конференции. Краснодар: Кубанский гос. ун-т, 1990. С. 55-57.
17. Тюрин В.В., Радецкий В.П., Сержант Л.А., Волчков Ю.А. Анализ комплекса признаков продуктивности у импортированных пород карпа // Сборник научных трудов ВНИИПРХ, вып.68, М.,1992. С. 45 – 51.
18. Тюрин В.В., Радецкий В.П., Сержант Л.А., Волчков Ю.А. Информативная селекционная оценка пород карпа по признакам продуктивности при прудовом выращивании // Сборник научных трудов ВНИИПРХ, вып.68, М.,1992. С. 52-59.
19. Илясов Ю.И., Волчков Ю.А., Тюрин В.В., Ганченко М.В., Михайлова С.Ш. Альбинизм канального сома и возможности его использования в двухлинейном разведении // Сборник научных трудов ВНИИПРХ, вып.68, М.,1992. С. 32-38.
20. Тюрин В.В., Волчков Ю.А. Оценка продуктивности при прудовом выращивании рыб по принципу апостериорной минимизации эффекта факторов среды. Деп. в ВИНИТИ, 1993 № 1973-В93. 12 с.
21. Тюрин В.В., Ганченко М.В., Сержант Л.А., Волчков Ю.А. Морфометрический и остеологический анализ белого толстолобика, полученного на геотермальных водах // Сборник науч. трудов ГосНИОРХ, Вып. 324, Спб., 1993. С. 56-61.
22. Kirpichnikov V.S., Ilyasov Ju.I., Vikhman A.A., Ganchenko M.V., Tyurin V.V.,Simonov V.M. Selection of Krasnodar common carp (Cyprinus carpio L.) for resistance to dropsy: principal results and prospects // Acuaculture, 111, 1993.Р. 7-20.
23. Тюрин В.В., Волчков Ю.А., Ганченко М.В., Радецкий В.П. Перспективы использования остеологических и морфометрических признаков в организации эколого-генетического мониторинга популяций рыб // «Актуальные вопросы экологии и охраны природы экосистем южных и центральных регионов России» Сб. материалов межреспубликанской конференции Краснодар, 1996. С. 67-70.
24. Тюрин В.В., Волчков Ю.А., Радецкий В.П. Эколого-генетический мониторинг в развитии методической базы селекции прудовых рыб // Тез. докладов региональной научной конференции «Современные проблемы экологии» Анапа, 1996. С. 43-45.
25. Тюрин В.В., Волчков Ю.А. Экоэлементная структура популяций черноморской мидии // «Актуальные вопросы экологии и охраны природных экосистем южных регионов России и сопредельных территорий». - материалы Х Межреспубликанской научно-практической конф. - Краснодар, 1997. С. 12-13.
26. Морев И.А., Тюрин В.В. Системный анализ изменчивости морфотипа в оценке закономерностей формообразования при отдаленной гибридизации осетровых // «Актуальные вопросы экологии и охраны природных экосистем южных регионов России и сопредельных территорий.» Краснодар, 1999 Материалы XII межреспубликанской научно-практической конференции. С. 41-43.
27. Волчков Ю.А., Морев И.А., Чмырь Ю.Н., Ловчикова Я.Б., Тюрин В.В. Структура искусственных популяций стерляди выявляемая в системном морфометрическом анализе // Сб. «Ресурсосберегающие технологии в аквакультуре». Краснодар, 1999. С. 52-54.
28. Чебанов М.С., Чмырь Ю.Н., Ловчикова Я.Б., Морев И.А., Тюрин В.В. Сравнительный морфометрический анализ искусственных популяций стерляди различного происхождения // Сб. «Ресурсосберегающие технологии в аквакультуре». Краснодар, 1999. С. 55-58.
29. Тюрин В.В. Корреляционная структура размерных показателей раковины черноморской мидии // «Актуальные вопросы экологии и охраны природы экосистем южных регионов России и сопредельных территорий». Краснодар 2000. Материалы XIII межреспубликанской научно-практической конференции. С. 68-69.
30. Тюрин В.В. Подход к разработке критерия отбора семей растительноядных рыб по продуктивности с точки зрения многомерной статистики // Международная научно-практическая конференцию «Проблемы воспроизводства растительноядных рыб и их роль в аквакультуре» Адлер, сентябрь 2000. С. 49-51.
31. Тюрин В.В., Ступин В.А., Алпеева И.Г. Структура популяций черноморской мидии выявляемая в анализе изменчивости показателей размера раковины // Актуальные вопросы экологии и охраны природы экосистем южных регионов России и сопредельных территорий.
Краснодар, 2001. С. 56-57.
32. Тюрин В.В., Ступин В.А., Круглов И.А., Алпеева И.Г. Межпопуляционная изменчивость комплекса морфометрических признаков раковины черноморской мидии (Мytilus galloprovincialis) // Актуальные вопросы экологии и охраны природы экосистем южных регионов России и сопредельных территорий. Краснодар, 2001. С. 61-62.
33. Тюрин В.В., Волчков Ю.А., Ступин В.А. Гетерогенность пород карпа, выявляемая в анализе изменчивости морфотипа // «Проблемы и перспективы развития аквакультуры в России». Материалы докладов всероссийской конференции, Краснодар 2001. С. 112-113.
34. Тюрин В.В., Волчков Ю.А., Алпеева И.Г., Ступин В.А. «Гетерогенность популяций черноморской мидии и перспективы селекции при организации искусственного воспроизводства и товарного выращивания » В соответствии с заданием региональной научно-технической программы «Марикультура Кубани». (депонированный отчет) Отчёт о НИР (заключительный) / КубГУ; Руководитель Тюрин В.В. – № ГР 01.20.0013039; Инв. № 02.200.203183. – Краснодар, 2001. 24 с.
35. Тюрин В.В., Волчков Ю.А., Ступин В.А., Пенкин Р.И. Идентификация пород карпа по комплексу морфометрических признаков // «Актуальные вопросы экологии и охраны природных экосистем южных регионов России и сопредельных территорий.» Краснодар, 2002 Материалы XV межреспубликанской научно-практической конференции С. 153-155
36. Тюрин В.В., Волчков Ю.А., Ступин В.А., Пенкин Р.И. Перспектива использования морфометрических индексов для анализа внутрисемейной гетерогенности сеголеток белого амура // «Актуальные вопросы экологии и охраны природных экосистем южных регионов России и сопредельных территорий.» Краснодар, 2002 Материалы XV межреспубликанской научно-практической конференции С. 155-157
37. Тюрин В.В., Алпеева И.Г. Изменчивость размерных показателей раковины черноморской мидии из популяций восточной части Черного моря // Актуальные вопросы экологии и охраны природных экосистем южных регионов России и сопредельных территорий." Краснодар, 2002 Материалы XV межреспубликанской научно-практической конференции С. 157-159
38. Морев И.А., Тюрин В.В. Анализ генетически обусловленной гетерогенности популяций стерляди, связанной с адаптивностью // «Актуальные вопросы экологии и охраны природных экосистем южных регионов России и сопредельных территорий.» Краснодар, 2002 Материалы XV межреспубликанской научно-практической конференции С. 159-160
39. Тюрин В.В., Волчков Ю.А., Ступин В.А., Бурло А.М. Оценка расстояний между породами карпа по комплексу морфометрических и биохимических признаков // «Актуальные вопросы экологии и охраны природных экосистем южных регионов России и сопредельных территорий.» Краснодар, 2003 Материалы XV межреспубликанской научно-практической конференции С. 140-142.
40. Тюрин В.В., Бурло А.М., Волчков Ю.А., Исследование формообразования при межпородной гибридизации карпа // «Актуальные вопросы экологии и охраны природных экосистем южных регионов России и сопредельных территорий.» Краснодар, 2032 Материалы XV межреспубликанской научно-практической конференции С. 143-145.
41. Тюрин В.В., Алехин А.С. Анализ изменчивости морфометрических признаков как адекватный подход к оценке гетерогенности маточных стад черноморского лосося // Проблемы естественного и искусственного воспроизводства рыб в морских и пресноводных водоемах. Тезисы международной научной конференции 9-10 июня 2004 Ростов-на-Дону. С. 10-12.
42. Тюрин В.В., Алёхин А.С., Волчков Ю.А. Разработка методов оценки генетической гетерогенности и принципов формирования ремонтно-маточного стада черноморского лосося на Адлерском производственно-экспериментальном лососёвом заводе (АПЭРЛЗ) Отчёт о НИР (закл.) / КубГУ; руководитель В.В. Тюрин – № ГР 01200411971; Инв. № 02200406109. – Краснодар, 2004. – 42 с.
43. Тюрин В.В., Алёхин А.С., Волчков Ю.А. Разработка комплекса информативных признаков для идентификации пород форели: Отчёт о НИР (закл.) / КубГУ; руководитель В.В. Тюрин – № ГР 01200509529; Инв. № 02200505664. – Краснодар, 2005. – 35 с.
44. Тюрин В.В., Алехин А.С. Изменчивость параметров осевого скелета черноморского лосося // Актуальные вопросы экологии и охраны природы экосистем южных регионов России и сопредельных территорий. Материалы XVIII межреспубликанской научно-практической конференции Краснодар. 2005. С. 166-167.
45. Тюрин В.В. Оценка генетической гетерогенности искусственных популяций черноморского лосося (Salmo trutta labrax) на основе анализа морфометрических признаков // Ekologia pogranicza EP’06 POLSKA AKADEMIA NAUK, Gorzow, 2006 Р. 438-444.
III. Учебно-методические работы
46. Тюрин В.В., Волчков Ю.А., Радецкий В.П., Илясов Ю.А. Методика сравнения групп рыб по продуктивности при прудовом выращивании. М., ВНИИПРХ, 1992. 10 с.
47. Тюрин В.В., Волчков Ю.А., Радецкий В.П., Илясов Ю.А. Приложение к «Методике сравнения групп рыб по продуктивности» (ЭВМ-программа). М., ВНИИПРХ, 1992. 18 с.
48. Тюрин В.В., Волчков Ю.А., Щеглов С.Н. Математические методы в биологии. Реализация с использованием пакета STATGRAPHICS 3.0. Кубанский государственный университет. 2000 г. 40 с.
49. Тюрин В.В., Морев И.А., Волчков Ю.А. Дискриминантный анализ в селекционно-генетических исследованиях (практикум). Кубанский государственный университет, 2002. 23 с.