

Распределение эритроцитов на разных уровнях артериального русла на основе их метаболических и структурных различий
На правах рукописи
Коваль Григорий Сергеевич
Распределение эритроцитов на разных уровнях артериального русла НА ОСНОВЕ их Метаболических и структурных РАЗЛИЧИЙ
03.00.13 – физиология
03.00.25 – гистология, цитология, клеточная биология
Автореферат
диссертации на соискание ученой степени
кандидата медицинских наук
Томск – 2008
Работа выполнена в Государственном образовательном учреждении высшего профессионального образования "Сибирский государственный медицинский университет Федерального агентства по здравоохранению и социальному развитию"
Научные руководители:
доктор медицинских наук,
профессор, академик РАМН,
Заслуженный деятель науки РФ Медведев Михаил Андреевич
доктор медицинских наук,
профессор Рязанцева Наталья Владимировна
Официальные оппоненты:
доктор медицинских наук,
профессор Байков Александр Николаевич
кандидат медицинских наук,
старший научный сотрудник Фомина Татьяна Ивановна
Ведущая организация:
ГУ НИИ физиологии СО РАМН
Защита состоится «___»_______________________2008 г. в ___часов на заседании диссертационного совета Д 208.096.01 при ГОУ ВПО СибГМУ Росздрава (634050, г. Томск, Московский тракт, 2)
С диссертацией можно ознакомиться в научно-медицинской библиотеке ГОУ ВПО СибГМУ Росздрава
Автореферат разослан «___»_______________________2008 г.

Ученый секретарь
диссертационного совета Суханова Г.А.
Общая характеристика работы
Актуальность проблемы. В настоящее время доказано существование механизма сепарации эритроцитов по признаку зрелости на уровне дуги аорты [Медведев М.А. и соавт., 1986]. Данный процесс рассматривается как часть общебиологического эволюционного процесса, способствующего адекватному энергетическому и кислородному обеспечению головного мозга [Шульговский В. В., 2003].
При интерпретации перераспределения кровотока в пользу жизненно важных органов, в первую очередь, – мозга, следует учитывать тот факт, что важна не столько абсолютная величина кровотока, сколько его соответствие кислородному запросу [Смирнов В.М. и соавт., 2007]. Характеристикой такого соответствия является соотношение между скоростью транспорта кислорода кровью, притекающей к головному мозгу, и величиной потребления им кислорода [Яковлев В.Н., 2006]. Обеспечить доставку кислорода к мозгу, адекватную его реальной потребности, позволяет преимущественное поступление в кровоток головного мозга молодых функционально полноценных форм эритроцитов. На основании исследования среднего диаметра эритроцитов, их сухой массы, кислотного гемолиза эритроцитов из сонной и бедренных артерий было показано существование адаптивного разделения эритроцитов на уровне дуги аорты на молодые (функционально полноценные) формы и на более зрелые формы [ Медведев М.А. и соавт., 1986]
Известно, что ключевую роль в определении структурной организации и функционирования эритроцита играет его мембрана. Являясь важными структурными и функциональными компонентами мембраны эритроцита, липидные молекулы регулируют подвижность и функциональную активность мембранных белков, тем самым обеспечивая в клетке селективную проницаемость и функционирование мембраноассоциированных ферментов, а также рецепторного аппарата [Камкин А.Г., Киселева И.С., 2008]. Помимо липидов, важнейшими компонентами эритроцитарной мембраны являются белки. Они выполняют множество функций: обеспечивают транспорт молекул внутрь клетки и из нее, в качестве ферментов катализируют ассоциированные с мембраной реакции, определяют морфологические и механические свойства клетки, служат рецепторами для получения и преобразования химических сигналов из окружающей среды, а также образуют антигенные детерминанты [Новицкий В.В. и соавт., 2004]. Свойства мембран эритроцитов, попадающих в мозговой кровоток, определяют их эластичность и способность проходить через мелкие капилляры головного мозга, являются важным для адекватного снабжения кислородом нервных клеток головного мозга.
Таким образом, учитывая важную роль мембраны в определении гомеостаза эритроцита, очевиден интерес к состоянию мембранных компонентов при изучении механизмов сепарации эритроцитов на уровне дуги аорты, что позволит более подробно изучить адаптационные механизмы организма, направленные на предотвращение гипоксических состояний мозга.
Цель исследования - провести сравнительный анализ распределения эритроцитов, поступающих в разные отделы артериального русла (восходящая аорта, общая сонная и бедренная артерии), на основе оценки их физико-химических свойств и морфологического статуса.
Задачи исследования:
- Дать комплексную сравнительную характеристику распределения эритроцитов, основываясь на данных исследования поверхностной архитектоники и формы эритроцитов в восходящей аорте, общей сонной и бедренной артериях у кроликов.
- Выявить особенности распределения эритроцитов на разных уровнях артериального русла по данным оценки липидного состава, активности Na+,K+-АТФазы и микровязкости липидной фазы эритроцитов.
- Оценить роль дуги аорты в распределении эритроцитов в различные отделы сосудистого русла на основе физико-химических особенностей их мембран.
Научная новизна. Впервые в экспериментах на животных проведено изучение особенностей метаболических и структурных свойств эритроцитов, поступающих в разные отделы артериального русла (дуга аорты, общая сонная и бедренная артерии). Получены приоритетные данные о перераспределении эритроцитов на уровне дуги аорты по физико-химическим характеристикам их мембран, направленном на преобладание в общей сонной артерии функционально полноценных эритроцитов, участвующих в транспорте кислорода к нервным клеткам головного мозга.
Выявлено преобладание морфологически трансформированных эритроцитов в бедренной артерии (переходные, предгемолитические и дегенеративные формы) по сравнению с морфологическими свойствами эритроцитов в общей сонной артерии. В бедренной артерии по сравнению с аортой и общей сонной артерией установлены выраженные изменения поверхностной архитектоники эритроцитов, приводящие к изменению их функциональной активности. Показано, что активность Na+,K+-АТФазы в мембранах эритроцитов в общей сонной артерии выше, чем в красных кровяных клетках из бедренной артерии. Установлены различия структуры липидного бислоя мембран эритроцитов на разных уровнях артериальной системы: в общую сонную артерию преимущественно поступают эритроциты с нормальным соотношением фракций фосфолипидов и микровязкостью липидной фазы мембран.
Практическая значимость работы. Данные о перераспределении эритроцитов на разных уровнях артериальной системы имеют важное значение для формирования фундаментальных знаний о механизмах кровообращения. Результаты исследования могут быть использованы как в области сердечно-сосудистой хирургии при разработке искусственных клапанов сердца и новых способов трансплантации сосудов, так и в области фармакологии – для целевой доставки лекарственных средств на основе знаний о функциональном и морфологическом различиях эритроцитов на разных уровнях артериального русла.
Положения, выносимые на защиту:
- На уровне дуги аорты имеет место распределение эритроцитов в зависимости от морфологических особенностей: количество трансформированных эритроцитов в бедренной артерии (переходные, предгемолитические и дегенеративные формы) увеличено по сравнению с их числом в общей сонной артерии.
2. Активность Na+,K+-АТФазы в мембранах эритроцитов, полученных из общей сонной артерии, выше, чем в эритроцитах из бедренной артерии.
3. Особенности липидной фазы мембран эритроцитов на разных уровнях артериальной системы характеризуются нарушением соотношения фосфолипидных фракций (увеличение содержания лизофосфатидилхолина и сфингомиелина, снижение доли фосфатидилхолина) и увеличением микровязкости в клетках, поступающих в бедренную артерию; липидный бислой мембран красных клеток крови в общей сонной артерии характеризуется нормальным соотношением липидных молекул и микровязкостью.
Внедрение результатов исследования. Основные положения исполь-зуются в курсе лекционных и практических занятий на кафедре нормальной физиологии ГОУ ВПО СибГМУ Росздрава, кафедре физиологии человека и животных ГОУ ВПО ТГУ, кафедре физиологии и валеологии ГОУ ВПО ТГПУ в разделах «Кровь» и «Физиология сердечно-сосудистой системы».
Апробация работы. Основные материалы доложены и обсуждены на XV Российско-японском медицинском симпозиуме (Благовещенск, 2007 г.), на ХХ съезде физиологического общества имени И.П. Павлова (Москва, 2007 г.), на VI съезде физиологов Казахстана с международным участием (Караганда, 2007 г.) и на VI Сибирском физиологическом съезде (Барнаул, 2008 г.).
Публикации
По материалам диссертации опубликовано 7 печатных работ, из них 1 в журнале из перечня ВАК.
Структура и объем диссертации
Диссертация изложена на 88 страницах, иллюстрирована 8 рисунками и состоит из введения, 4 глав, обсуждения, выводов, указателя литературы из 195 источников (73 отечественных и 122 иностранных авторов).
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Эксперименты проводили на 24 кроликах обоих полов породы «Шиншилла». Экспериментальные животные содержались в виварии ГОУ ВПО СибГМУ Росздрава, в рацион питания входили зерновые культуры и сухая трава. Для исследования использовали кровь, полученную с помощью катетеризации общей сонной артерии, дуги аорты и бедренной артерии. Кровь забирали в объеме 5 мл в стерильную пробирку, стабилизировали гепарином (50 ЕД/мл крови). Все вмешательства осуществляли с соблюдением принципов Хельсинской декларации Всемирной медицинской ассоциации (1989 г.), Европейской конвенции по охране позвоночных животных, Директивами-86/609/ECC [Council Directive 86/609/ECC (24 November 1986) on the approximation of laws, regulations and administrative provisions of the Member States regarding the protection of animals used for experimental and other scientific purposes].
Оценку поверхностной архитектоники эритроцитов осуществляли методом сканирующей электронной микроскопии. Для этого биоптаты периферической крови обрабатывали в соответствии с методикой, предложенной Г.И. Козинцом и соавт. [1977]. В образцах у 50-ти произвольно выбранных эритроцитов измеряли внешний диаметр клетки и размер центральной впадины, вычисляли процентное соотношение между ними. Фотографирование эритроцитарных клеток производили на электронном микроскопе “JEМ-100” при увеличении 1000-1500.
Мембраны эритроцитов выделяли по методу J.T. Dodge [1963], основанному на феномене гипоосмотического гемолиза клеток. Для этого 10 мл венозной гепаринизированной крови (50 ЕД/мл) центрифугировали при 3000 об/мин в течение 10 мин. Пастеровской пипеткой удаляли плазму. Оставшуюся клеточную взвесь трижды отмывали 10 мМ трис-НСl-буфером (рН 7,4) и 0,145 М NaCl, центрифугируя каждый раз в течение 10 мин при 3000 об/мин и удаляя надосадок.
Определение активности Nа+,K+-АТФазы в мембранах эритроцитов проводили методом, разработанным А.М. Казенновым и соавт. [1984] и основанном на накоплении неорганического фосфора (Pi) в среде, содержащей АТФ, в результате его гидролиза под действием АТФазы. Уровень Pi определяли по методу P.S. Chen et al. [1956].
Липиды мембран эритроцитов экстрагировали хлороформ-метаноловой смесью по методу J. Folch et al. [1957], в основу которого положен принцип первичного разрушения липид-белковых связей полярным растворителем (метанолом) с последующим экстрагированием липидов неполярным растворителем (хлороформом). Общее содержание липидов в мембранах эритроцитов определяли по методу W. Bloor в модификации J. Bragdon, основанному на окислении липидов хромовой кислотой.
Препаративное разделение нейтральных липидов проводили методом тонкослойной хроматографии [Финдлей Дж.Б., Эванз У.Г., 1990] в системе растворителей гептан : диэтиловый эфир : этилацетат (в соотношении 80:20:1,5) на пластинках “Silufol UV 254” (Чехия). Полученный липидный экстракт мембран выпаривали на водяной бане. Липидный остаток, разведенный в гептане, наносили на линию старта, расположенную на расстоянии 1,5 см от края пластин, которые помещали в камеру с вышеописанной смесью на 15-20 мин. Пятна обнаруживали нагреванием пластин до 1000С в сушильном шкафу после предварительного опрыскивания 2% спиртовым раствором фосфорномолибденовой кислоты.
Для разделения фракций фосфолипидов мембран эритроцитов методом тонкослойной хроматографии [Прохорова М.И., Тупикова З.Н., 1965; Прохорова М.И., 1982] сухой липидный остаток разводили гептаном и наносили на пластинки “Silufol UV 254” (Чехия). Разделение осуществляли в системе хлороформ : метанол : вода (в соотношении 32:12,5:2). Пластинки высушивали, опрыскивали раствором фосфорномолибденовой кислоты, проявляли в сушильном шкафу.
Спектрофлуориметрическое исследование мембран эритроцитов выполняли с использованием флуоресцентного зонда пирена. Измерение собственной флуоресценции теней эритроцитов и определение спектральных характеристик взаимодействия мембран с флуорофорами проводили на спектрофлуориметре «СМ-2203» (Беларусь). Для определения собственной флуоресценции теней эритроцитов, основанного на способности триптофановых остатков белковых компонентов мембран флуоресцировать под воздействием возбуждающего излучения, матричную мембранную взвесь разводили 10 мМ трис-НСl буфером (рН 7,4) и 0,145 М раствором NaCl до конечной концентрации белка 0,3 мг/мл. 2 мл полученного раствора мембран эритроцитов помещали в стандартную кварцевую кювету с длиной оптического пути 1 см в 0,9 % растворе NaCl и измеряли флуоресценцию при длине волны возбуждающего света 285 нм, щелях 2/3 нм. Максимум флуоресценции регистрировали при 340 нм [Владимиров Ю.А., Добрецов Г.Е., 1980].
Статистический анализ проводили с помощью пакета программ “Statistica for Windows 6.0”. Вычисляли среднее арифметическое значение (M), стандартное отклонение (s). Оценку нормальности распределения полученных результатов по каждой величине проводили с помощью критерия Колмогорова-Смирнова. Так как распределение признаков не соответствовало нормальному, был использован непараметрический Т-критерий Вилкоксона для зависимых выборок. Критическое значение уровня значимости принималось равным 0,05.
Для выявления однородных структур (кластеров) в исследуемых данных был применен метод древовидной кластеризации, растояние между кластерами измеряли с помощью евклидового расcтояния с использованием одиночной связи.
РЕЗУЛЬТАТЫ исследования и их обсуждение
Особенности морфологических свойств эритроцитов
на разных уровнях артериального русла
Исследование поверхностной архитектоники эритроцитов выявило изменение процентного соотношения их морфологических форм у кроликов в сосудах разного уровня.
В качестве образца крови до сепарации эритроцитов в дуге аорты мы использовали кровь, взятую из нижней части восходящей аорты, до отхождения плечеголовного ствола и других крупных сосудов. Проведенное исследование морфологических свойств эритроцитов показало, что эритроцитарный пул в этом отделе, в основном, представлен функционально полноценными двояковогнутыми дискоцитами с симметричными углублениями и гладкой клеточной поверхностью, суммарное содержание которых составило 87,30±0,81 %. На долю переходных форм эритроцитов, способных при благоприятных физиологических условиях к обратной трансформации в дискоциты, приходилось 7,60±0,06 %. К их числу были отнесены эллипсовидные клетки (0,56±0,05%), эритроциты в виде плоского диска (1,12±0,06%), дискоциты с единичными (2,36±0,06%) и множественными (1,38±0,04%) выростами конической формы с округлой вершиной, дискоциты с гребнем (1,37±0,04%), а также крайне редко встречающиеся в кровеносном русле эритроциты в виде «тутовой ягоды» со множеством тонких и длинных конусообразных выростов в виде шипов (0,78±0,05%). Лишь незначительная часть эритроцитарной популяции была представлена необратимо измененными предгемолитическими эритроцитами с гладкой поверхностью, принимающими куполообразную (1,02±0,03%) и сферическую (1,91±0,04%) форму, а также клетками в виде «спущенного мяча» (1,21±0,07%). Количество необратимо трансформированных и дегенеративно измененных форм клеток составило 4,10±0,06% и 1,01±0,05%, соответственно (табл. 1).
Содержание нормальных двояковогнутых эритроцитов в сонной артерии составило 88,70±1,33%, количество обратимо трансформированных форм эритроцитов - 7,20±0,38%. В сонной артерии реже, чем в восходящей аорте, встречались необратимо трансформированные предгемолитические формы (на 13 % меньше). Содержание дегенеративно измененных форм эритроцитов в сонной артерии было почти в 2 раза меньше их количества в аорте.
При оценке морфологического состава эритроцитов в бедренной артерии обращало на себя внимание достоверно значимое (p<0,01) снижение содержания двояковогнутых дискоцитов по сравнению с их показателями в восходящей аорте. Выявлено увеличение численности переходных (на 20%), необратимо трансформированных (на 21%) и дегенеративных форм (на 62%) в бедренной артерии по сравнению с аортой (табл. 1, рис. 1).
Сепарация преимущественно в бедренную артерию трансформированных эритроцитов (эллипсовидные клетки, эритроциты в виде “плоского диска” и “тутовой ягоды”, эхиноциты, стоматоциты, сфероциты, клетки в виде “спущенного мяча”), на наш взгляд, физиологически обоснована, поскольку их наличие является, с точки зрения микроциркуляции в головном мозге, неблагоприятным признаком, так как способствует снижению кислородтранспортной функции, критичной для нормальной работы нервных клеток. Видоизмененные эритроциты менее полноценны и стойки, чем двояковогнутые дискоциты и не соответствуют критериям физиологической полноценности интактных клеток красной крови [Berling C. et al., 1988; Reinhart W.H. et al, 1992; Степовая Е.А., 1998; Новицкий В.В. и соавт., 2000; Рязанцева Н.В. и соавт., 2005; Huabing L. et al., 2007].
Результаты кластерного анализа показали, что различия в распределении эритроцитов между аортой и бедренной артерией более выражены, чем между аортой и общей сонной артерией. Предположительно, это связано с тем, что бедренная артерия отходит после бифуркации брюшной аорты, где также возможно определенное перераспределение эритроцитов.
Таблица 1
Распределение морфологических форм эритроцитов (в %) на разных уровнях артериального русла у кроликов
(по данным сканирующей электронной микроскопии) (![]() ±m)
±m)
| Морфологические формы эритроцитов | Восходящая аорта (n=10) | Общая сонная артерия (n=10) | Бедренная артерия (n=10) |
| 87,30±0,81 | 88,70±1,33 p1>0,05 | 82,80±2,23 p1<0,01, p2 < 0,01 |
| Эллипсовидные эритроциты | 0,56±0,05 | 0,52±0,06 p1>0,05 | 0,76±0,06 p1<0,05, р2<0,05 |
| Эритроциты в виде плоского диска | 1,12±0,06 | 1,26±0,08 p1>0,05 | 1,87±0,02 p1<0,01, р2<0,01 |
| Дискоциты с выростом | 2,36±0,06 | 2,27±0,05 p1>0,05 | 1,87±0,02 p1<0,001,р2<0,001 |
| Дискоциты с гребнем | 1,37±0,04 | 1,54±0,04 p1<0,01 | 1,18±0,05 p1<0,01, р2<0,01 |
| Дискоциты с множественными выростами | 1,38±0,04 | 1,22±0,05 p1<0,05 | 2,54±0,08 p1<0,01, p2<0,01 |
| Эритроциты в виде тутовой ягоды | 0,78±0,05 | 0,39±0,03 p1<0,05 | 1,27±0,08 p1<0,01, р2<0,01 |
| Куполообразные эритроциты | 1,02±0,03 | 1,09±0,05 p1>0,05 | 1,32±0,05 p1<0,01, p2<0,05 |
| Сферические эритроциты | 1,91±0,04 | 1,46±0,05 p1<0,05 | 2,61±0,08 p1<0,01, р2<0,01 |
| Эритроциты в виде спущенного мяча | 1,21±0,07 | 1,01±0,03 p1<0,05 | 1,27±0,03 p1>0,05, р2>0,05 |
| 1,01 ± 0,05 | 0,57±0,02 p1<0,01 | 1,63±0,04 p1<0,01, р2<0,001 |
Примечание: здесь и в табл. 2-5 р1 - достоверность различий параметров по сравнению с аналогичными показателями в восходящей аорте; р2 - в сонной артерии.
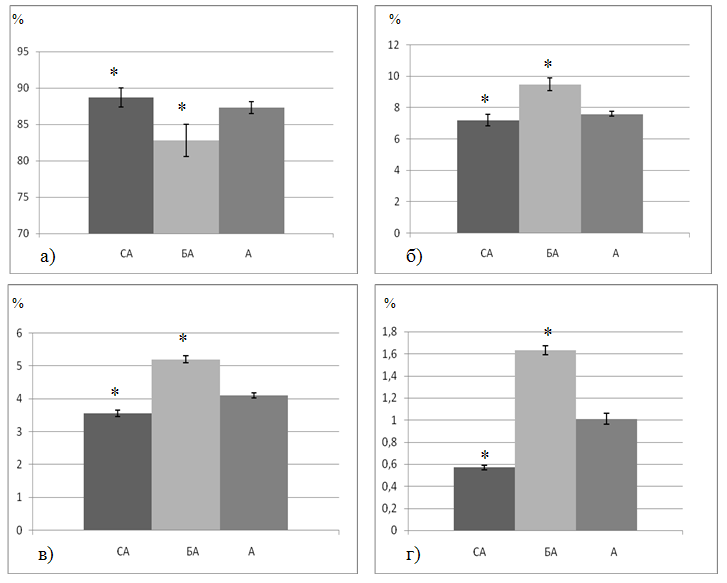
Рис. 1. Перераспределение двояковогнутых дискоцитов (а), переходных (б), предгемолитических (в) и дегенеративных форм эритроцитов (г) на уровне дуги аорты у экспериментальных животных (по данным сканирующей электронной микроскопии). А – восходящая аорта, БА – бедренная артерия, СА – общая сонная артерия.
По результатам цитометрического исследования эритроцитов можно сделать вывод, что значения соотношения внутреннего и внешнего диаметров дискоцитов, полученных из аорты, составляли 52,71±0,71%. В сонной артерии эти значения достоверно (p<0,05) превышали соответствующий показатель в аорте, составив 54,86±1,22%; в бедренной артерии данная величина не отличалась от характеристик эритроцитов, полученных при катетеризации восходящей аорты (52,31±1,12%) (рис. 2).
Таким образом, результаты анализа распределения морфологических форм эритроцитов на разных уровнях артериального русла выявили преобладание функционально-полноценных форм в общей сонной артерии. В бедренной артерии повышено содержание трансформированных эритроцитов, характеризующихся нарушением функциональной активности.
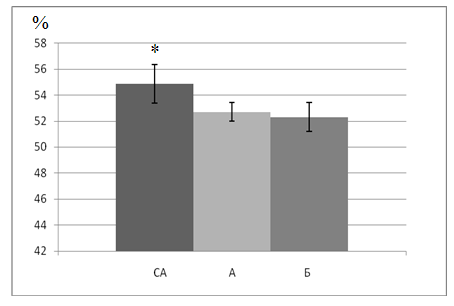
Рис. 2. Перераспределение эритроцитов на уровне дуги аорты у экспериментальных животных (СА – общая сонная артерия, А – аорта, Б – бедренная артерия).
Особенности активности Na+,K+-АТФазы в мембранах эритроцитов на разных уровнях артериального русла
Поскольку активность и свойства транспортных ATФаз плазматических мембран в значительной степени определяются свойствами липидного матрикса, в который погружены молекулы ферментов, при старении эритроцитов нарушается активность ионтранспортирующих систем. К их числу относится Na+,K+-АТФаза, играющая важную роль в поддержании ионного гомеостаза и сохранении функциональной активности эритроцитов. Активность Na+,K+-АТФазы в мембранах эритроцитов, выделенных из восходящей аорты у кроликов, составила 0,097±0,006 мкМ Pi/ч*мг белка, из бедренной артерии - 0,089±0,005 мкМ Pi/ч*мг белка, из общей сонной артерии - 0,115±0,005 мкМ Pi/ч*мг белка (табл. 2). Сравнительный анализ изменения активности Na+,K+-АТФазы в эритроцитарных мембранах продемонстрировал достоверные различия: активность данного фермента в мембранах эритроцитов в сонной артерии была выше его активности в эритроцитах аорты на 9,3 %, в то время как в эритроцитах, полученных из бедренной артерии, активность Na+,K+-АТФазы была на 18,5 % ниже значений активности этого фермента в мембранах эритроцитов из восходящей аорты.
Таблица 2
Активность Na+,K+-АТФазы в мембранах эритроцитов у кроликов на разных уровнях артериального русла (![]() ±m)
±m)
| Место забора крови | Активность Na+,K+-АТФазы, ммоль Pi /часмг белка |
| Восходящая аорта (n = 14) | 0,097±0,006 |
| Общая соонная артерия (n = 14) | 0,115±0,005 p1<0,01 |
| Бедренная артерия (n = 14) | 0,089±0,005 p1>0,05 p2<0,05 |
Активность Na+,K+-АТФазы в мембранах эритроцитов играет существенную роль в определении их функциональных свойств [Vermeulen W.P. et al., 1996; Schulpis K. H., 2006]. Кроме того, установлено, что активность Na+,K+-АТФазы изменяется с возрастом эритроцитов и может рассматриваться как показатель их старения [Pillips M. et all., 2000].
На основании полученных результатов можно заключить, что активность Na+,K+-АТФазы в мембранах эритроцитов на разных уровнях кровеносного русла существенно различается: в клетках, полученных из общей сонной артерии, активность этого фермента выше, чем в клетках из бедренной артерии, что является подтверждением факта перераспределения эритроцитов на уровне дуги аорты - эритроциты с большей активностью Na+,K+-АТФазы поступают в мозговой кровоток, в то время как эритроциты с низкой активностью Na+,K+-АТФазы - в другие органы и системы организма.
Особенности структуры липидной фазы мембран эритроцитов
на разных уровнях артериального русла
Путем регистрации степени эксимеризации неполярного зонда пирена, диффундирующего в гидрофобном компартменте мембраны, с использованием флуоресцентного зондирования нами было проведено исследование микровязкостных свойств липидной фазы мембран эритроцитов как в области белок-липидных контактов (при длине волны возбуждающего света (в), равной 285 нм), так и всего липидного бислоя мембран эритроцитов (при в=340 нм).
Выявлено, что средние величины отношения интенсивностей флуоресценции димерной и мономерной форм липотропного зонда пирена (I470/I340) при длине волны возбуждающего света 340 нм в мембранах эритроцитов, полученных из разных отделов артериальной системы, различны: в восходящей аорте – 0,347±0,008 усл.ед., в сонной и бедренной артериях – 0,386±0,006 и 0,306±0,007 усл.ед., соответственно (рис. 3). Вместе с тем, эксимеризация пирена в области анулярной липидной фракции осуществлялась с гораздо большими ограничениями, на что указывали несколько сниженные средние значения I470/I340 при в=285 нм в мембранах эритроцитов, составлявшие в восходящей аорте 0,305±0,011 усл.ед., в сонной и бедренной артериях – 0,343±0,009 и 0,284±0,008 усл.ед., соответственно. Соотношение интенсивностей флуоресценции мономеров пирена (I370/I390) при в=340 нм, отражающее полярность микроокружения зонда в интегральной липидной фазе эритроцитарных мембран, в восходящей аорте оказалось равным 1,010±0,014 усл.ед. и не отличалось от исследуемой величины в общей сонной (1,010±0,010 усл.ед.) и бедренной артериях (0,995±0,011 усл.ед.) (p>0,05) (табл. 3).
Наряду с этим, процент индуктивно-резонансного переноса энергии с триптофана на пирен, позволяющий судить о белково-липидных взаимодействиях в мембранах, составил в восходящей аорте – 69,16±1,17%, в сонной артерии – 71,83±1,35%, в бедренной – 69,16±2,15% (статистический анализ показал, что данный показатель в исследуемых выборках значимо не отличался) (табл. 3).
Таблица 3
Показатели флуоресценции зонда пирена в мембранах эритроцитов у кроликов на разных уровнях артериального русла (![]() ± m)
± m)
| Место забора крови | Параметры флуоресценции, усл. ед. | Величина миграции энергии с триптофана на пирен, % | ||
| в=285 нм | в=340 нм | |||
| I470/I340 | I470/I340 | I370/I390 | ||
| Восходящая аорта (n=14) | 0,305±0,011 | 0,347±0,008 | 1,010±0,014 | 69,16±1,17 |
| Общая сонная артерия (n=14) | 0,343±0,009 p1<0,01 | 0,386±0,006 p1<0,01 | 1,010±0,010 p1>0,05 | 71,83±1,35 p1<0,05 |
| Бедренная артерия (n=14) | 0,284±0,008 p1<0,01, p2<0,01 | 0,306±0,007 p1<0,01, p2<0,01 | 0,995±0,011 p1>0,05, p2>0,05 | 69,16±2,15 p1 >0,05, p2>0,05 |
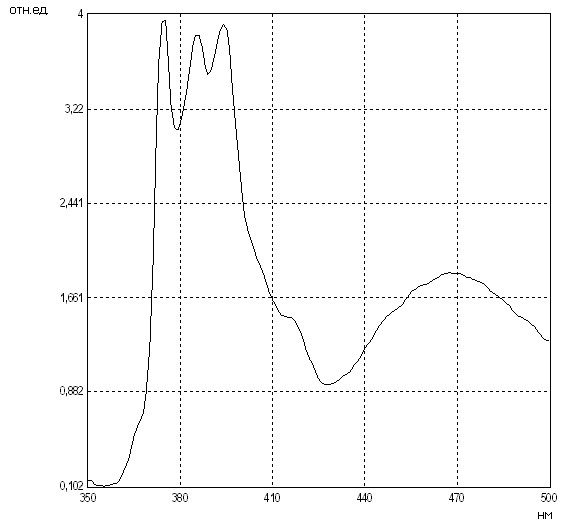
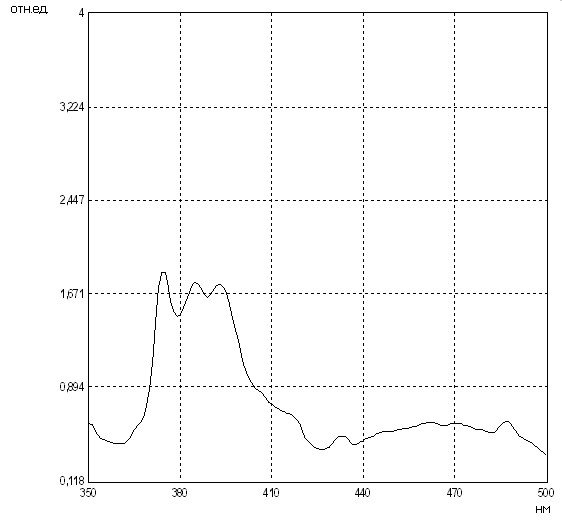
Рис. 3. Особенности флуоресценции мембран эритроцитов из восходящей аорты кроликов (Ex: 285.0 нм; SEx: 0.5 нм; SEm: 1.0 нм) (a), (Ex: 340.0 нм; SEx: 0.5 нм; SEm: 1.0 нм) (б)
Особенности липидного состава мембран эритроцитов
на разных уровнях артериального русла
В ходе проведенного нами исследования было установлено, что содержание общих липидов в мембранах эритроцитов у кроликов из аорты составляет 0,93±0,02 мг/мг белка. Препаративное разделение нейтральных липидов эритроцитарных мембран, полученных из восходящей аорты кроликов, проводимое методом тонкослойной хроматографии, выявило преобладание фракции общих фосфолипидов, составившей 44,69±0,81%. На долю фракций холестерина и эфиров холестерина приходилось 36,20±1,14 и 19,11±0,65%, соответственно (табл. 4).
Среди фракций фосфолипидов выделяли лизофосфатидилхолин (4,01±0,11%), фосфатидилинозитолы (5,25±0,09%), сфингомиелин (18,72±0,13%), фосфатидилхолин (33,88±0,17%), фосфатидилсерин (6,54±0,07%), а также фосфатидилэтаноламин (31,59±0,26%) (табл. 5). При этом каких-либо различий показателей, характеризующих липидный спектр мембран эритроцитов у кроликов разных полов, зарегистрировано не было.
Таблица 4
Относительное содержание фракций фосфолипидов, холестерина и эфиров холестерина в мембранах эритроцитов у кроликов (в %), полученных на разных уровнях артериальной системы (![]() ±m)
±m)
| Фракции липидов | Место забора крови | ||
| Восходящая аорта (n=14) | Общая сонная артерия (n=14) | Бедренная артерия (n=14) | |
| Фосфолипиды | 44,69±0,81 | 44,83±0,82 p1>0,05 | 44,84±0,95 p1>0,05, p2>0,05 |
| Холестерин | 36,20±1,14 | 36,81±0,97 p1>0,05 | 35,65±0,96 p1>0,05 p2>0,05 |
| Эфиры холестерина | 19,11±0,65 | 18,34±0,87 p1>0,05 | 19,51±0,64 p1>0,05 p2>0,05 |
- Исследование особенностей липидного спектра мембран эритроцитов, полученных из общей сонной артерии, показало, что содержание общих липидов в мембранах составиляло 0,95±0,03 мг/мг белка. Доля общих фосфолипидов, холестерина и эфиров холестерина соответствовала таковой в мембранах эритроцитов в восходящей аорте (табл. 4), также не отличалось содержание фракций фосфатидилинозитолов, фосфатидилсерина и фосфатидилэтаноламина. Уровень лизофосфатидилхолина и сфингомиелина был снижен, а содержание фракции фосфатидилхолина повышено по сравнению с содержанием в мембранах эритроцитов из аорты (табл. 5).
- В мембранах эритроцитов, полученных из бедренной артерии у кроликов (табл. 5), содержание общих липидов составило 0,91±0,02 мг/мг белка. При анализе содержания фракций фосфолипидов в мембранах эритроцитов из бедренной артерии было отмечены следующие отличия от мембран клеток из восходящей аорты и общей сонной артерии: снижение содержания фракции фосфатидилхолина, повышение содержания фракций сфингомиелина и лизофосфолипидов. Фракции фосфатидилинозитолов, фосфатидилсерина и фосфатидилэтаноламина не отличались по своему процентному содержанию от таковых в мембранах эритроцитов из восходящей аорты и общей сонной артерии (табл. 5).
Таблица 5
Относительное содержание фракций фосфолипидов в мембранах эритроцитов у кроликов (в %) на разных уровнях артериального русла (![]() ±m)
±m)
| Фракции фосфолипидов | Место забора крови | ||
| Восходящая аорта (n=14) | Сонная артерия (n=14) | Бедренная артерия (n=14) | |
| Лизофосфатидилхолин | 4,01±0,11 | 3,51±0,06 p1<0,05 | 4,98±0,17 p1<0,05 p2<0,01 |
| Фосфатидилинозитолы | 5,25±0,09 | 5,07±0,16 p1>0,05 | 4,99±0,10 p1>0,05 p2>0,05 |
| Сфингомиелин | 18,72±0,13 | 16,95±0,14 p1<0,01 | 21,17±0,17 p1<0,01 p2<0,01 |
| Фосфатидилхолин | 33,88±0,17 | 36,50±0,21 p1<0,01 | 31,29±0,16 p1<0,01 p2<0,01 |
| Фосфатидилсерин | 6,54±0,07 | 6,48±0,06 p1>0,05 | 6,35±0,07 p1>0,05 p2>0,05 |
| Фосфатидилэтаноламин | 31,59±0,26 | 31,48±0,28 p1>0,05 | 31,20±0,22 p1>0,05 p2>0,05 |
Примечательно, что результаты кластерного анализа фракций фосфолипидов, полученных из мембран эритроцитов на разных уровнях артериальной системы, оказались созвучны результатам кластерного анализа распределения морфологических форм эритроцитов, описанных выше: фракции липидов мембран эритроцитов из общей сонной артерии и восходящей аорты объединялись в один кластер, в отличие от мембран эритроцитов из бедренной артерии, что доказывает возможность распределения эритроцитов на уровне дуги аорты.
Поддержание соотношения между фракциями фосфолипидов мембраны на физиологическом уровне служит непременным условием нормального функционирования эритроцита. Дезорганизация липидов, являющихся субстратом как перекисного окисления липидов, так и ферментативного гидролиза, является причиной утраты способности клеток регулировать ионный и антиоксидантный гомеостаз, нарушения работы мембраносвязанных энзимов, что, в конечном итоге, способствует изменению метаболизма клетки, а также приводит к необратимым нарушениям ее структурно-функционального статуса [Величко Л.Н., Федорова В.М., 1981; Backman L. et. al., 1998]. Увеличение среднего уровня лизофосфолипидов в эритроцитах, полученных из бедренной артерии, может приводить к изменению проницаемости мембран для органических молекул и ионов, солюбилизации мембранассоциированных ферментов, усилению экзоцитоза [Condrea E., 1980; Rogausch H., 1984; Грибанов Г.А., 1991; Сороковой В.И. и соавт., 1994; Zavodnik I.B. et al., 1997].
Нарушение распределения эритроцитов по качественному признаку между общей сонной артерией и другими крупными сосудами, отходящими от дуги аорты, могут играть существенную роль в механизмах формирования гипоксии головного мозга при различных патологических изменениях дуги аорты и аортального клапана сердца. Результатом этого является нарушение адекватного участия красных кровяных клеток в газообмене, обусловленного их уникальной способностью к деформации в мелких сосудах головного мозга. Тем более, что, согласно данным литературы, ишемия мозга и деформируемость красных кровяных клеток, взаимосвязанные явления: в случаях, когда у крыс моделируют процесс ишемии мозга, имеют место процессы нарушения деформируемости эритроцитов [Плотников М.Б. и соавт., 1995]. Справедливо и обратное утверждение – при нарушениях, связанных с изменением деформируемости эритроцитов и их трансформацией, при различных патологических состояниях (поражение клапанного аппарата сердца, изменение эластичности аорты и ее формы), развивается ишемия головного мозга [Барбараш Л.С. и соавт., 1995]. Эти данные, на наш взгляд, отражают важность процесса перераспределения эритроцитов по их функциональной активности в дуге аорты.
В целом, по итогам проведенного исследования были получены приоритетные данные, свидетельствующие о выраженном распределении эритроцитов в дуге аорты и за ее пределами, что указывает на важность процессов турбулентного течения крови в артериях крупного калибра. Физико-химические характеристики мембран эритроцитов в восходящем отделе аорты, общей сонной и бедренных артериях различаются между собой: в общую сонную артерию поступают клетки с «оптимальными» для адекватного газообмена функциональными свойствами.
Выводы
- Эритроциты в разных отделах артериального русла характеризуются морфологической неоднородностью: в общей сонной артерии по сравнению с восходящей частью аорты снижено содержание необратимо трансформированных эритроцитов и преобладают нормальные двояковогнутые дискоциты; в бедренной артерии на фоне снижения числа нормальных двояковогнутых дискоцитов количество переходных форм (эллипсовидные эритроциты, клетки в виде плоского диска, дискоциты с одним и множественными выростами, эритроциты в виде тутовой ягоды), предгемолитических форм (куполообразные и сферические эритроциты) и дегенеративно измененных форм эритроцитов значительно повышено.
- В мембранах эритроцитов, поступающих после перераспределения на уровне дуги аорты в общую сонную артерию, активность Na+,K+-АТФазы в 1,3 раза выше активности этого ионтранспортирующего фермента, в мембранах эритроцитов, выделенных из бедренной артерии у экспериментальных животных.
- Мембраны эритроцитов, полученных из бедренной артерии у кроликов, отличаются по своим физико-химическим свойствам от мембран эритроцитов из общей сонной артерии и характеризуются: а) возрастанием микровязкости как в области интегральной липидной фазы при в=340 нм, так и в области белок-липидных контактов при в=280 нм; б) изменением фосфолипидного спектра мембран эритроцитов (увеличение содержания лизофосфатидилхолина и сфингомиелина, снижение доли фосфатидилхолина).
- Реализация феномена перераспределения эритроцитов на уровне дуги аорты обусловливает поступление функционально полноценных клеток с оптимальными физико-химическими и структурными характеристиками мембраны преимущественно в общую сонную артерию, что может закономерно приводить к улучшению снабжения кислородом нервных клеток головного мозга.
Список работ, опубликованных по теме диссертации
- Коваль, Г.С. Изменение поверхностной архитектоники и диаметра эритроцитов после их распределения на уровне дуги аорты / Г.С. Коваль, М.А. Медведев // Материалы XV Российско-японского медицинского симпозиума. – Благовещенск, 2007. – С. 114.
- Морфологический профиль эритроцитов после их распределения на уровне дуги аорты / Г.С. Коваль, М.А. Медведев, Н.В. Рязанцева и др. // Тезисы докладов ХХ съезда физиологического общества имени И.П. Павлова. – Москва, 2007. – С.266.
- Характеристика структурных свойств мембран эритроцитов с использованием флуоресцентного зондирования после их распределения на уровне дуги аорты / Г.С. Коваль, М.А. Медведев, Н.В. Рязанцева, М.А. Чурбанова // Материалы VI съезда физиологов Казахстана с международным участием. – Караганда : Изд-во КГМА, 2007. – С.42-43.
- Медведев, М.А. Физиологическая сепарация эритроцитов на уровне дуги аорты / М.А. Медведев, Г.С. Коваль, Н.В. Рязанцева // Бюллетень Сибирской медицины: научно-практический журнал. - 2007. – Т.6. – № 4. - С. 37-40.
- Физиологическое распределние эритроцитов на уровне дуги аорты по данным цитометрического и спектрофлуометрического исследований / М.А. Медведев, Г.С. Коваль, Н.В. Рязанцева, М.А. и др. // Вестник Томского государственного университета. – 2007. – № 300 (II). – С.170-171.
- Юрьева, В.Д. Структурный профиль и активность Na+/K+-АТФазы эритроцитов после их распределения на уровне дуги аорты / В.Д. Юрьева, Г.С. Коваль // Материалы Всероссийской 66-й итоговой студенческой научной конференции им. Н.И. Пирогова. – Томск, 2007. – С.373-374.
- Коваль, Г.С. Морфофункциональная характеристика распределения эритроцитов на разных уровнях артериального русла / Г.С. Коваль, М.А. Медведев, Н.В. Рязанцева // Материалы VI Сибирского физиологического съезда. – Барнаул, 2008.–Т.1.– С.94–95.