

Биоразнообразие долгоносикообразных жуков (coleoptera, curculionoidea) агроэкосистем при адаптивном эколого-ландшафтном земледелии на севере лесостепи приволжской возвышенности
На правах рукописи

НИКОЛАЕВА Татьяна Геннадьевна
БИОРАЗНООБРАЗИЕ ДОЛГОНОСИКООБРАЗНЫХ ЖУКОВ (COLEOPTERA, CURCULIONOIDEA) АГРОЭКОСИСТЕМ ПРИ АДАПТИВНОМ ЭКОЛОГО-ЛАНДШАФТНОМ ЗЕМЛЕДЕЛИИ НА СЕВЕРЕ ЛЕСОСТЕПИ ПРИВОЛЖСКОЙ ВОЗВЫШЕННОСТИ
Специальность 03.02.08 – экология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Казань – 2012
Работа выполнена в лаборатории агроэкологических разработок ГБУ Институт проблем экологии и недропользования Академии наук Республики Татарстан
| Научный руководитель: | кандидат биологических наук, Григорьян Борис Рубенович |
| Научный консультант: | кандидат биологических наук, ведущий научный сотрудник Егоров Леонид Валентинович |
| Официальные оппоненты: | доктор сельскохозяйственных наук, профессор, заведующий кафедрой защиты растений и селекции ФГОУ ВПО «Казанский государственный аграрный университет» Сафин Радик Ильясович кандидат биологических наук, доцент кафедры зоологии беспозвоночных ФГАОУ ВПО «Казанский (Приволжский федеральный университет» Шулаев Николай Вячеславович |
| Ведущая организация: | ФГБОУ ВПО «Кубанский государственный аграрный университет», г. Краснодар |
Защита диссертации состоится «24» мая 2012г. в 1430 часов на заседании диссертационного совета ДМ 212.081.19 при ФГАОУ ВПО «Казанский (Приволжский) федеральный университет» по адресу: 420008, г. Казань, ул. Кремлевская, д. 18, главное здание, ауд. 211.
Отзыв на автореферат в двух экземплярах с подписями, заверенными гербовой печатью, просим направлять по адресу: 420008, Республика Татарстан, г. Казань, ул. Кремлевская, д. 18, К(П)ФУ, диссертационный совет ДМ 212.081.19.
Факс: (843) 238-71-21; (843) 231-52-40; e-mail: [email protected]
С диссертацией можно ознакомиться в Научной библиотеке им. Н.И.Лобачевского ФГАОУВПО «Казанский (Приволжский) федеральный университет» по адресу:
г. Казань, ул. Кремлевская, д. 35.
Автореферат разослан «24» апреля 2012 года

Ученый секретарь диссертационного совета,
кандидат биологических наук, доцент
Зелеев Равиль Муфазалович
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность. Одной из основных причин снижения биоразнообразия является интенсификация использования природных ресурсов при сельскохозяйственной деятельности (Одум, 1987; Букштынов, 1989; Миркин и др., 1996), поэтому особое внимание в последнее время уделяется вопросам внедрения принципов экологического земледелия и сохранения биоразнообразия в различных агроландшафтах (Сумароков, 2003; Пойрас, 2006; Шпанев, Голубев, 2010 и др.). Многовековой опыт ведения сельского хозяйства показал, что при формировании современных агроэкосистем необходимо ориентироваться на ландшафтно-экологическое земледелие (Каштанов, 1983; Ильина и др., 2001), направленное на устойчивое развитие агроценозов и рациональное использование природных ресурсов.
При адаптивном эколого-ландшафтном земледелии возделывание сельскохозяйственных культур проводится на беспестицидной основе, в связи с чем, для планирования природоохранных мероприятий, проведения экологических экспертиз, мониторинга и оптимизации фитосанитарного контроля агрокультур, необходимы сведения о состоянии энтомофауны. В наземной фауне умеренных широт среди жесткокрылых надсемейство Curculionoidea по численности особей занимает доминирующее положение (Ольшванг, Богачева, 1990). Это самая богатая видами группа высокоспециализированных растительноядных насекомых (Коротяев, 2000), являющаяся одним из постоянных и самых многочисленных компонентов любого наземного сообщества беспозвоночных животных. В качестве консументов первого порядка, Curculionoidea играют важную роль в трофических, энергетических и информационных цепях биоценозов. Большинство представителей Curculionoidea узкоспециализированные фитофаги, разнородные по преферендумам к абиотическим условиям среды, активные на протяжении сезона и в течение суток, отличающиеся длительными периодами питания на личиночной и имагинальной стадиях, что характеризует их в качестве удобной модельной группы для изучения биоразнообразия любой наземной территории.
В работах, посвященных долгоносикообразным жукам типичных агроценозов, исследователи концентрируют свое внимание в основном на отдельных вредителях различных культур (Бей-Биенко, 1946; Алимджанов, 1951; Храмушин, 1954; Рязанцев, 1960; Карабатов, 1969; Петруха, 1969; Тураев, 1969; Тураев, Карабатов, 1969а, б; Байтенов, 1975; Коротяев, 1983 и др.). Действительно, среди Curculionoidea есть виды, способные наносить вред овощным, зерновым, кормовым и древесно-кустарниковым растениям. Видимо, поэтому с сельскохозяйственной точки зрения значение куркулионоидных жесткокрылых до последнего времени рассматривалось довольно односторонне, лишь в качестве вредителей, в то время как среди представителей надсемейства существуют и полезные виды – энтомофаги, инквилины и фитофаги сорных и паразитических растений, которые выступают в качестве биологических агентов и могут быть использованы при ведении устойчивого сельского хозяйства (Де Бах, 1968; Пойрас, 2006; Коротяев, 2012). Оценка состояния фауны данной группы жуков важна при планировании природоохранных мероприятий и проведении экологического мониторинга в агроценозах (Чичков, Легалов, 2008).
Однако, составляя существенную долю в сообществах хортобионтных беспозвоночных в различных ценозах, малоизученными остаются эколого-фаунистические аспекты разнообразия долгоносикообразных жуков агроэкосистем при адаптивном эколого-ландшафтном земледелии. Обобщающие сведения по фауне и экологии Curculionoidea агроэкосистем при адаптивно-ландшафтном земледелии на исследуемой территории в литературе не представлены.
Цель и задачи исследования. Цель настоящей работы заключается в выявлении особенностей таксономического разнообразия и экологической структуры фауны долгоносикообразных жесткокрылых агроэкосистем при адаптивном эколого-ландшафтном земледелии на севере лесостепи Приволжской возвышенности.
Для достижения поставленной цели решались следующие задачи:
1) Выявить таксономическое разнообразие надсемейства Curculionoidea.
2) Изучить экологическую структуру надсемейства Curculionoidea в трофическом, пространственном и биотопическом аспектах.
3) Исследовать синэкологические аспекты биоразнообразия Curculionoidea.
4) Охарактеризовать комплекс экономически важных в хозяйственном отношении видов долгоносикообразных жуков.
Научная новизна. Впервые представлена эколого-фаунистическая характеристика долгоносикообразных жесткокрылых агроценозов при адаптивном эколого-ландшафтном земледелии на севере лесостепи Приволжской возвышенности. Впервые для фауны Чувашской Республики (ЧР) приведены сведения о 4 видах Curculionoidea: Ceratapion basicorne (Illiger, 1807), Squamapion cineraceum (Wencker, 1864), Bagous lutulentus Gyllenhal 1813, Ceutorhynchus roberti Gyllenhal, 1837. В фауне района исследования впервые выявлено 4 семейства (Anthribidae Billberg, Attelabidae Billberg, Rhynchitidae Gistel, Dryophthoridae Schoenherr), 8 подсемейств, 32 трибы, 84 рода и 189 видов куркулионоидных жесткокрылых. Охарактеризован комплекс видов, имеющих хозяйственное значение для исследованной агроэкосистемы. Выявлена зависимость видового богатства жуков и показателей биоразнообразия Curculionoidea от видового богатства фитоценозов, а также установлена количественная зависимость между представителями Curculionoidea со средним и широким трофическим спектром и типами адаптивных стратегий растений модельных биотопов.
Теоретическая и практическая значимость. Полученные сведения вносят вклад в изучение экологии и распространения долгоносикообразных жуков при адаптивно-ландшафтном земледелии. Результаты исследования значительно расширяют представление о куркулионоидных жесткокрылых и в дальнейшем могут быть использованы при разработке, планировании и проведении мероприятий по моделированию устойчивых агроэкосистем. Приведено научное обоснование эффективности внедрения адаптивно-ландшафтной беспестицидной системы земледелия для устойчивого развития агроэкосистемы и сохранения биоразнообразия.
Материалы исследований могут быть включены в Летописи природы особо охраняемых природных территорий (ООПТ), а также использованы при составлении энтомологического кадастра Чувашской Республики, в учебных курсах биологического, экологического и сельскохозяйственного направлений. Показано, что для оценки биоразнообразия агроэкосистем в условиях экологического земледелия в качестве модельной группы могут быть использованы долгоносикообразные жуки. Информация о вредоносных и полезных видах Curculionoidea является важной при организации мониторинговых исследований на территории изучаемой агроэкосистемы и региона в целом.
Положения, выносимые на защиту:
1) Видовое разнообразие и структура фауны долгоносикообразных жесткокрылых агроэкосистем при адаптивном эколого-ландшафтном земледелии соответствуют заповедным природно-естественным комплексам.
2) Наибольшим видовым богатством фауны долгоносикообразных жесткокрылых агроэкосистем при адаптивном эколого-ландшафтном земледелии на севере лесостепи Приволжской возвышенности характеризуются разнотравные мезофитные луга, экотонные околоводно-лугово-лесные и лесо-луговые участки.
3) Своеобразие фауны долгоносикообразных жуков при адаптивном эколого-ландшафтном земледелии по сравнению с таковой агроценозов с традиционной системой земледелия заключается в преобладании видов с узкой пищевой специализацией, наличии видов-энтомофагов, более широком спектре фитобионтных групп и биотопических группировок.
Апробация работы. Основные положения диссертации докладывались и обсуждались на конференциях по экологии, зоологии и сельскому хозяйству, ежегодных семинарах ИПЭН АН РТ. На Международных конференциях: «Экологическое равновесие и устойчивое развитие территории» (С.-Петербург, 2010), «Роль особо охраняемых природных территорий в сохранении биоразнообразия» (Чебоксары, 2010), «Экологическое нормирование, сертификация и паспортизация почв как научно-инновационная основа рационального землепользования» (Москва, 2010), Пущинских школах-конференциях молодых ученых (Пущино, 2010, 2011), «Ломоносов» (Москва, 2011, 2012), ZOOCENOSIS-2011: «Биоразнообразие и роль животных в экосистемах» (Днепропетровск, 2011); Всероссийских: «Полевые и экспериментальные исследования биологических систем» (Ишим, 2010), «Биоразнообразие: глобальные и региональные процессы» (Улан-Удэ, 2010), «Инновационные разработки молодых ученых – АПК России» (Казань, 2010), «Научное наследие В.И. Вернадского и современные проблемы науки» (Чебоксары, 2010), «Устойчивость экосистем: теория и практика» (Чебоксары, 2010), «Принципы и способы сохранения биоразнообразия» (Йошкар-Ола, 2010), «Симбиоз – Россия 2010» (Н. Новгород, 2010).
Публикации. По теме диссертации опубликовано 17 работ общим объемом 6.2 печатных листа (п.л.), в т.ч. 4 статьи в журналах, рекомендованных ВАК.
Обоснованность и достоверность результатов работы основаны на репрезентативности собранного материала, его камеральной обработке, применением современных методов математической статистики при обработке и анализе данных.
Личный вклад автора. Автором разработана программа исследований, осуществлен сбор материала, проведена его камеральная и статистическая обработка, интерпретация полученных данных, обобщение результатов и формулировка выводов. Идентификация большинства видов долгоносикообразных жуков проведена автором.
Структура и объем диссертации. Диссертационная работа состоит из введения, 9 глав, выводов, библиографического списка, включающего 354 источника, в том числе 49 на иностранных языках. Работа изложена на 229 страницах (из которых 168 страниц основного текста), содержит 29 таблиц, 42 рисунка, 3 приложения.
Благодарности. Работа выполнена в Институте проблем экологии и недропользования Академии наук РТ под руководством к.б.н., доцента Б.Р. Григорьяна, которому автор выражает благодарность за помощь, советы и научные консультации на всех этапах проведения исследования. Автор глубоко благодарен и признателен научному консультанту к.б.н. Л.В. Егорову (г. Чебоксары) за проверку правильности определения энтомологического материала и советы при окончательном оформлении работы, а также Б.А. Коротяеву (ЗИН РАН, г. С.-Петербург) за идентификацию некоторых видов. Особую благодарность автор выражает н.с., к.б.н. М.Ш. Сибгатуллиной (ГБУ ИПЭН АН РТ) за помощь в определении видовой принадлежности растений, к.б.н. Л.П. Тепловой (г. Чебоксары) за уточнения и проверку правильности определения некоторых видов растений; за ценные советы при статистической обработке данных с.н.с., к.б.н. Р.А. Суходольской (ГБУ ИПЭН АН РТ); к.б.н. В.Н. Подшивалиной (г. Чебоксары), с.н.с. Л.М. Сунгатуллиной (г. Казань) за внимание к работе и замечания по рукописи автореферата; работникам СХПК «Ленинская искра» Ядринского района ЧР Герасимову В.Г., Игнатьеву В.Я., Сапожникову Н.Л. за возможность проведения исследования на территории хозяйства. Автор благодарен родным и близким за всестороннюю помощь, поддержку и понимание.
СОДЕРЖАНИЕ РАБОТЫ
Глава 1. Экологическое земледелие и его роль в сохранении биоразнообразия
Представлено понятие об экологическом земледелии, рассмотрена история его развития в мире и России, показано положительное влияние данного вида земледелия на различные живые организмы. Приведена характеристика адаптивно-ландшафтного земледелия как одного из видов экологического земледелия; общая характеристика модельного хозяйства и обзор сведений о разнообразии различных групп организмов в изучаемом агроландшафте. Показано состояние изученности фауны и экологии долгоносикообразных жуков на территории исследования.
Глава 2. Условия обитания фауны
Представлено краткое физико-географическое описание территории исследования: охарактеризовано положение района в различных системах природного и хозяйственного районирования, приведены сведения об особенностях рельефа, климата, гидрологической сети, почвенного покрова (в т.ч. агрохимическая характеристика исследуемых участков), растительности. Приведены ботанические описания модельных групп биотопов, таксономическая и экологическая структуры растительного покрова агроэкосистемы.
Глава 3. Материал и методы исследования
3.1. Материал исследования
Материалом для диссертации послужили собственные сборы автора, выполненные в течение относительно умеренно-теплого (2009 г.) и засушливого (2010 г.) полевых сезонов на территории производственного участка СХПК «Ленинская искра» Ядринского района ЧР. Исследованы буферные участки и окраины агроценозов под зерновыми культурами (озимая пшеница, яровая пшеница, овес, ячмень), кормовыми травосмесями, многолетними и однолетними травами. Сбор материала выполнен в 15 пунктах с помощью традиционного набора общепринятых методов (Фасулати, 1971). Все изученные биотопы объединены в 5 групп: разнотравные луга с отдельными деревьями и различным режимом увлажнения на склонах (группа биотопов №1), сосново-рябиновые лесополосы вблизи мезофитных и гигрофитных лугов (№2), одиночные березово-липовые лесополосы (№3), околоводно-прибрежные биотопы (№4), зарастающие участки поля на окраине смешанного леса (№5). Учетная единица (проба) составила 100 взмахов. При проведении сборов отмечались показания температуры и влажности травянистого яруса. Всего собрано 15836 экземпляров долгоносикообразных жуков. Количество отобранных проб в каждом биотопе составило свыше 60. Пробы отбирались в 3-кратной повторности.
3.2. Математическая обработка результатов
Видовое богатство (ВБ) сообществ долгоносикообразных жуков и фитоценозов оценивали следующими параметрами: уровнем, концентрацией, потенциалом и оригинальностью (Мордкович и др., 2002; Мордкович, 2004). Для оценки биоразнообразия фауны Curculionoidea использовали индексы видового разнообразия фауны Маргалефа (DMg) и Менхиника (DMn), индекс биоразнообразия Шеннона (H*) и вычисленный на его основе показатель выравненности фауны (Е), меры доминирования: индекс Симпсона (1D) и индекс полидоминантности (1/D) (Мэгарран, 1992; Лебедева и др., 1999). Для выявления общности объектов по качественным данным позитивным совпадениям применяли индекс Чекановского-Съеренсена (ICS) (Песенко, 1982). При анализе сходства фаун модельных групп биотопов использовали кластерный анализ (Халафян, 2008). Доминантные группы в пределах биотопов выделены на основании общеевропейской шкалы численного обилия Ренконена (Renkonen, 1938). Кроме того, для оценки общего обилия использована логарифмическая шкала (Песенко, 1982). При изучении биотопической приуроченности видов Curculionoidea применяли ряд показателей: встречаемость, верность биотопу, участие биотопа в размещении популяции вида, степень относительной биотопической приуроченности (Песенко, 1982). При рассмотрении пространственного распределения Curculionoidea использовали показатель степени пространственной агрегированности (отношение дисперсии к среднему значению) – 2/m (Гиляров, 1990). При оценке численного обилия рассчитывались общепринятые показатели: среднее арифметическое значение (M) и ошибка средней (m), стандартное отклонение () и коэффициент вариации (V) (Лакин, 1990). Приведена ошибка выборочной доли в % (sp%) (Лакин, 1990). Корреляционный анализ данных проводили с использованием коэффициентов корреляции Пирсона (r) и Спирмена (rS) (Лакин, 1980), учитывали статистически значимые зависимости (p<0.05). При выявлении структурных различий между модельными группами биотопов применяли метод многомерной статистики дискриминантный анализ. Достоверность различий между значениями оценивали по критерию Стьюдента (tSt) и Краскела-Уоллиса (Лакин, 1980).
Номенклатура надвидовых таксонов долгоносикообразных жуков приведена преимущественно по работе Алонсо-Сарасаги и Лайаля (Alonso-Zarazaga, Lyal, 1999), для подсемейства Entiminae – в соответствии с работой Юнакова Н.Н. и др. (Yunakov, 2012). Идентификация типов адаптивных (эколого-фитоценотических) стратегий растений проводилась по работам J. Grime et. al (1988) и D. Frank, S. Klotz (1988).
Статистическая обработка данных и построение гистограмм проводились с помощью стандартных пакетов Microsoft Exel 2007 и Statistica 8.0 (StatSoft, Inc. 2007).
Глава 4. Таксономическое разнообразие фауны Curculionoidea агроэкосистемы
Целенаправленных исследований по изучению фауны долгоносикообразных жуков при адаптивно-ландшафтной системе земледелия в регионе не проводилось. Несмотря на наличие подробных сведений о фауне долгоносикообразных жуков севера лесостепи Приволжской возвышенности (Дмитриева, 2005, 2006) и хорошей изученности группы в регионе исследования (ЧР), до начала наших исследований существующие литературные источники содержали лишь отрывочные данные по некоторым единичным видам долгоносикообразных жуков агроценозов при адаптивной эколого-ландшафтной системе земледелия, полученные в ходе мониторинговых исследований по изучению почвенной мезофауны и герпетофауны полей (Мутиков и др., 1996; Кириллова и др., 2006; Егоров и др., 2007) и в ряде общих работ по жесткокрылым и насекомым Чувашской Республики (Егоров, Исаев, 2001; Егоров, Прокопьева, 2002; Егоров, 2004, 2006; Егоров, Хрисанова, 2006). К началу наших исследований в перечисленных работах для территории указано 30 видов Curculionoidea из 5 семейств. Кроме того, изучение опубликованных данных по проводимым зоологическим исследованиям в соседнем регионе в КП «Чулпан» Высокогорского района Республики Татарстан, в котором адаптивная эколого-ландшафтная система также внедрена в полном объеме, показало, что основное внимание исследователей уделено почвенной мезофауне (Пухачев и др., 2000; Шафигуллина, Гордиенко, 2008) и долгоносикообразные жуки рассмотрены лишь как представители группы насекомых-фитофагов.
По результатам наших исследований на территории изучаемой агроэкосистемы при адаптивной эколого-ландшафтной системе земледелия выявлено 215 видов долгоносикообразных жуков из 99 родов, принадлежащих 45 трибам, 15 подсемействам и 8 семействам (табл. 1), что составляет более 40% фауны Чувашской Республики. Нашими исследованиями не была охвачена обширная группа ксилобионтных долгоносиков, поскольку их экология и методы сбора принципиально отличаются от филлофагов (Падий, 1972; Олигер, 1989; Лабинов, Егоров, 1996; Юркина и др., 1997; Лабинов, Егоров 1998; Мозолевская, 1998; Федоренко, 1998; Егоров, Лабинов, 1999). Тем не менее, в ходе сборов обнаружено 2 вида из семейства короедов Scolytidae: Scolytus mali и S. rugulosus, информация о которых для исследуемого района ранее не приводилась.
Таблица 1
Таксономическое разнообразие надсемейства Curculionoidea агроэкосистемы при адаптивном эколого-ландшафтном земледелии
| Семейство | Число подсемейств | Число триб | Число родов | Число видов | ||||
| родов | % | sp% | видов | % | sp% | |||
| Anthribidae | 1 | 2 | 2 | 2.02 | 1.41 | 2 | 0.93 | 0.65 |
| Attelabidae | 1 | 1 | 1 | 1.01 | 1.00 | 1 | 0.47 | 0.47 |
| Rhynchitidae | 3 | 6 | 6.06 | 2.40 | 7 | 3.25 | 1.21 | |
| Apionidae | 1 | 7 | 21 | 21.21 | 4.11 | 51 | 23.72 | 2.9 |
| Nanophyidae | 1 | 1 | 1 | 1.01 | 1.00 | 1 | 0.47 | 0.47 |
| Dryophthoridae | 1 | 1 | 1 | 1.01 | 1.00 | 1 | 0.47 | 0.47 |
| Erirhinidae | 1 | 2 | 3 | 3.03 | 1.72 | 4 | 1.86 | 0.92 |
| Curculionidae | 9 | 28 | 64 | 64.65 | 4.80 | 148 | 68.83 | 3.16 |
| Всего: | 15 | 45 | 99 | 100.00 | 215 | 100.00 | ||
Помимо 4 новых для фауны ЧР видов долгоносикообразных жуков нами выявлено 11 редких видов из 4 семейств: Anthribus nebulosus, Cyanapion columbinum, Eutrichapion punctiger, Ischnopterapion loti, Pseudorchestes circumvistulanus, Ceutorhynchus sulcicollis, Otiorhynchus (Pseudocryphiphorus) conspersus, Sitona waterhousei, Larinus obtusus, Lixus albomarginatus, Scolytus rugulosus (Егоров, 2004, 2005; Егоров, Егорова, 2009).
В целом, с учетом литературных данных фауна долгоносикообразных жуков агроэкосистемы представлена 216 видами из 100 родов, принадлежащих к 45 трибам, 15 подсемействам и 8 семействам, фауна Ядринского района Чувашской Республики включает 218 видов долгоносикообразных жуков из 101 рода, принадлежащих к 46 трибам, 16 подсемействам и 8 семействам.
Наиболее крупными семействами, составляющие в сумме более 92% видового богатства изученной фауны Curculionoidea агроэкосистемы, являются Curculionidae (148 видов 68.83%) и Apionidae (51 вид – 23.72%) (табл. 1), составляющие ядро фауны и естественных биоценозов (Дмитриева, 2006; Хрисанова, 2006а и др.). При этом наибольший вклад в таксономическое разнообразие долгоносикообразных жуков на родовом (64 рода – 64.65%) и видовом уровнях вносит семейство Curculionidae. Доля остальных семейств значительно ниже и не превышает 3% на видовом уровне и 6% на родовом. Семейства Attelabidae, Nanophyidae и Dryophthoridae наименее богаты и представлены одним видом, что вполне закономерно, поскольку для данных семейств в природе изначально характерно небольшое число видов (Исаев, 2007).
Наиболее богаты видами подсемейства: Apioninae (51 вид, 21 род, 7 триб), Curculioninae (46 видов, 22 рода, 10 триб), Entiminae (38 видов, 14 родов, 8 триб), Ceutorhynchinae (34 вида, 17 родов, 4 трибы), Hyperinae (10 видов, 2 рода, 1 триба), Lixinae (9 видов, 4 рода, 2 трибы), Mesoptiliinae (5 видов, 1 род, 1 триба), Erirhininae (4 вида, 3 рода, 2 трибы). На родовом уровне наибольший вклад в разнообразие фауны Curculionoidea агроэкосистемы вносят: Sitona (12 видов), Сeutorhynchus (10), Hypera (9), Tychius (8), Phyllobius (7), Oxystoma, Protapion и Polydrusus (по 6 видов), Cyanapion, Eutrichapion и Magdalis (по 5 видов), Apion, Perapion, Gymnetron, Rhinoncus и Larinus (по 4 вида), Ceratapion, Squamapion, Curculio, Anthonomus, Bagous, Microplontus, Otiorhynchus и Lixus (по 3 вида), охватывающие около 60% фауны. Выявленная структура фауны Curculionoidea агроэкосистемы на родовом уровне во многом совпадает с аналогичной, характерной в целом для фауны Curculionoidea севера лесостепи Приволжской возвышенности (Дмитриева, 2006).
Сравнение видового богатства фауны Curculionoidea рассматриваемой агроэкосистемы при адаптивном эколого-ландшафтном земледелии с типичными агроценозами с традиционной системой земледелия и фауной долгоносикообразных жуков природных комплексов проводилось на основе собственных данных (Егоров, Николаева, 2004, 2005 а, б, 2006 а, б; Николаева, Егоров, 2007 а, б) и литературных источников (Исаев, 1994 [1996]; Егоров, Дмитриева, 2000 а, б, в; Дмитриева, 2002 а, б; Чичков, Легалов, 2008; Шпанев, Голубев, 2010). Для корректного сравнения нами учитывались лишь те литературные данные, которые содержали наиболее полную информацию о составе фауны Curculionoidea с высокой степенью изученности и сходным методом сбора. Выяснили, что по видовому богатству изучаемая фауна долгоносикообразных жесткокрылых при адаптивно-ландшафтной беспестицидной системе земледелия практически не уступает фауне природно-естественных комплексов (ГПЗ «Присурский» Чувашской Республики и Жигулевский заповедник Самарской области) и примерно в 2.5-3.5 раза превосходит фауну изучаемой группы жуков при традиционной системе земледелия. Как известно, большее видовое разнообразие обуславливает большую устойчивость сообщества (Лопатин, 1997), следовательно, можно утверждать, что агроэкосистема при адаптивно-ландшафтной беспестицидной системе земледелия более устойчива по сравнению с традиционными агроландшафтами и более приближена к природным биоценозам.
Глава 5. Количественные показатели фауны
5.1. Соотношение семейств по видовому богатству и численному обилию в модельных группах биотопов
Представители семейства Attelabidae обнаружены лишь на зарастающих лесо-полевых участках, Dryophthoridae – на разнотравных лугах, Anthribidae – преимущественно на зарастающих участках поля на окраине леса, а также в прибрежно-околоводных биотопах, Nanophyidae – в мезофитных и мезогигрофитных биотопах (в сосновых лесополосах вблизи мезофитных и гигрофитных лугов, прибрежно-околоводных комплексах и увлажненных понижениях вблизи сосново-широколиственного леса) (рис. 1). К увлажненным местообитаниям тяготеют и представители семейства Erirhinidae, встреченные нами в 4 модельных группах биотопов. Во всех группах биотопов отмечены представители трех семейств Rhynchitidae, Apionidae и Curculionidae, причем, высокая встречаемость и широкая представленность семейства Rhynchitidae не характерна для типичных агроценозов. По-видимому, создание полезащитных лесополос, искусственных древесных насаждений, лесов способствовало формированию благоприятных условий для развития фауны Rhynchitidae в агроэкосистеме.

Рис. 1. Представленность семейств Curculionoidea по числу видов в модельных группах биотопов агроэкосистемы (расшифровку номеров групп биотопов см. гл. 3).
По сравнению с умеренно-теплым сезоном представители семейств Anthribidae, Rhynchitidae и Erirhinidae имеют более широкий диапазон биотопического распространения в засушливый сезон, что, возможно, связано с поиском более благоприятных местообитаний с оптимальными микроклиматическими показателями. Представители семейств Apionidae и Curculionidae встречены во всех биотопах в течение двух сезонов и преобладают по численному обилию. Доля Apionidae по численному обилию снижается в засушливый сезон во всех биотопах кроме разнотравных лугов и опушки сосново-широколиственного леса, что связано с биологическими особенностями представителей семейства, чутко реагирующих на повышенные температуры и мигрирующих с травянистого на древесно-кустарниковый ярус. Для семейств Apionidae и Curculionidae в засушливый сезон характерно увеличение степени агрегированности пространственного распределения. По-видимому, большинство представителей семейств избегают биотопов с резким изменением абиотических условий среды, в первую очередь, температуры и влажности в пределах травянистого яруса, и предпочитают умеренно-влажные биотопы с небольшими колебаниями температуры.
5.2. Параметры видового богатства фауны Curculionoidea агроэкосистемы
Согласно критерию Краскела-Уоллиса модельные группы биотопов статистически значимо различаются по видовому богатству (H=69.27, p=0.00001). Согласно двухвыборочному Z-тесту статистически значимо по видовому богатству от всех модельных групп отличаются березово-липовые лесополосы (p<0.0007). За период исследований наибольшее число видов, родов, а также максимальные показатели концентрации видового богатства (64.65% и 63.72% соответственно) отмечены на разнотравных лугах и в комплексе прибрежно-околоводных биотопов (табл. 2). Кроме того, разнотравные луга на протяжении всех изученных сезонов отличаются высокими значениями потенциала видового богатства, что обусловлено наличием богатой кормовой базы для долгоносикообразных жуков, а также разнообразием абиотических условий среды (различным режимом увлажнения, уровнем инсоляции и скоростью ветра на склонах). Известно, что толща травостоя обладает весьма специфическими биотическими и абиотическими условиями и создает очень широкий спектр экологических ниш (Лагунов, 1998). Высокое значение потенциала видового богатства свидетельствует о потенциально большем видовом обилии фауны биотопа.
Наибольшее число оригинальных видов (21 вид) (встречающихся только в конкретной группе биотопов агроэкосистемы), а соответственно, и максимальные значения оригинальности видового богатства (15.33%) выявлены для фауны Curculionoidea прибрежно-околоводных биотопов. Причем, стоит отметить, что показатели оригинальности фауны данной группы биотопов сильно различаются между сезонами. В умеренно-теплый сезон число оригинальных видов (7 видов) и показатель оригинальности видового богатства (8.23) имеют довольно низкие значения по сравнению с аналогичными параметрами в засушливый сезон (22 вида и 18.03% соответственно). По-видимому, в засуху в прибрежно-водные биотопы из ближайших биоценозов мигрируют в поисках благоприятных микроклиматических условий для обитания луговые и лугово-лесные виды, о чем свидетельствуют высокие показатели концентрации видового богатства, число видов и родов. При этом, истинно околоводная фауна отличается бедным видовым составом, о чем свидетельствует низкое значение оригинальности биотопа в умеренно-теплый сезон. Околоводные и болотно-околоводные виды в сумме составляют всего 6.52%, в то время как доля видов, предпочитающих околоводные биотопы помимо основных, довольно существенна – 31.63%. Именно за счет таких видов фауна околоводных биотопов отличается высоким видовым богатством.
Таблица 2
Параметры видового богатства долгоносикообразных жуков модельных биотопов агроэкосистемы в условиях адаптивно-ландшафтного земледелия
| Параметры ВБ | Группы биотопов | ||||
| №1 | №2 | №3 | №4 | №5 | |
| Число видов | 139 | 117 | 86 | 137 | 126 |
| Число родов | 69 | 63 | 45 | 72 | 65 |
| Число оригинальных видов | 17 | 10 | 7 | 21 | 17 |
| Концентрация ВБ, % | 64.65 | 54.42 | 40.00 | 63.72 | 58.60 |
| Потенциал ВБ | 2.01 | 1.86 | 1.91 | 1.90 | 1.94 |
| Оригинальность ВБ, % | 12.23 | 8.55 | 8.14 | 15.33 | 13.49 |
В умеренно-теплый сезон наибольшее число видов, родов и оригинальных видов выявлено на зарастающих участках поля у леса, характеризующихся также высокой концентрацией (58.17%) и оригинальностью (16.85%) видового богатства, что объясняется выраженным эффектом экотона и участием мелколиственного молодняка в формировании второго яруса (Галата и др., 1969). В засушливый сезон происходит нарушение протекающих сукцессионных процессов, что негативно отражается на значениях концентрации видового богатства и оригинальности. Данный факт свидетельствует о неустойчивости и несформированности фауны данной группы биотопов. В целом, для зарастающих участков поля характерны средние показатели параметров видового богатства.
Средними значениями параметров видового богатства отличаются также сосновые лесополосы. Присутствие представителей луговых сообществ в фауне сосновых лесополос вследствие пограничного расположения с мезофитными и гигрофитными лугами способствовало повышению изначально бедного видового богатства фауны данной группы биотопов. Существенных различий в показателях видового богатства между сезонами не выявлено, что может свидетельствовать об относительно стабильном развитии фауны долгоносикообразных жуков данной группы биотопов.
Наименьшее значение концентрации видового богатства, а также число выявленных видов и родов зарегистрировано для одиночных березово-липовых лесополос в связи с бедным видовым составом произрастающих в данном биотопе растений. Наличие в видовом составе монофагов и узких олигофагов обусловило среднее значение оригинальности фауны данной группы биотопов по итогам каждого сезона, но в среднем характерно наименьшее значение оригинальности видового богатства фауны (табл. 2).
5.3. Оценка видового разнообразия и выравненности фаун долгоносикообразных жуков модельных групп биотопов
Видовое разнообразие отражает сложность строения и структуру сообщества. Мониторинговые исследования и контроль над уровнем биоразнообразия предполагает возможность его измерения. Оценивание биологического разнообразия имеет важное прикладное значение, поскольку позволяет контролировать сохранение генетического потенциала и дает представление о состоянии экосистем на определенной территории. Параметры видового богатства предоставляют информацию о качественной компоненте биоразнообразия, в то время как важными составляющими альфа-разнообразия являются: число видов и относительное обилие видов (Лебедева и др., 1999). Расчет критерия Краскела-Уоллиса указывает на значимость различий по индексу Шеннона (H=98.50, p=0.0001), Симпсона (H=87.54, p=0.00001), полидоминантности (H=87.58, p=0.00001), Маргалефа (H=91.18, p=0.00001), Менхиника (H=112.62, p=0.0001), показателю выравненности (H=89.96, p=0.00001) между модельными группами биотопов. Согласно двухвыборочному Z-тесту статистически значимых различий по всем параметрам не выявлено между разнотравными лугами и околоводно-прибрежными комплексами, а также околоводными биотопами и сосновыми лесополосами.
Высокие значения индексов биоразнообразия в течение всех периодов исследования отмечены на разнотравном лугу и в околоводно-прибрежных биотопах, что обусловлено высокими показателями выравненности населения сообщества долгоносикообразных жуков и его полидоминантной структурой (табл. 3).
Таблица 3
Средние значения индексов видового разнообразия фауны
Curculionoidea в модельных группах биотопов в течение двух сезонов (2009-2010 гг.)
| Сезон | показатели | Группы биотопов | ||||
| 1 | 2 | 3 | 4 | 5 | ||
| умеренно-теплый (2009 г.) | S | 46.00±2.08 | 48.00±2.65 | 31.33±2.19 | 52.00±4.51 | 49.33±1.20 |
| H* | 3.12±0.11 | 2.71±0.31 | 1.86±0.46 | 2.90±0.20 | 2.38±0.60 | |
| Е | 0.81±0.02 | 0.70±0.08 | 0.54±0.13 | 0.74±0.04 | 0.61±0.15 | |
| 1D | 0.93±0.02 | 0.83±0.08 | 0.66±0.15 | 0.88±0.02 | 0.71±0.13 | |
| 1/D | 14.70±2.62 | 8.53±3.11 | 4.61±2.07 | 9.68±2.62 | 9.71±7.29 | |
| DMg | 8.15±0.31 | 7.97±0.24 | 5.12±0.29 | 8.71±0.76 | 8.20±0.98 | |
| DMn | 2.92±0.06 | 2.52±0.14 | 1.63±0.12 | 2.79±0.25 | 2.70±0.83 | |
| засушливый (2010 г.) | S | 56.00±12.00 | 43.20±6.30 | 18.20±4.19 | 49.80±11.15 | 38.60±9.56 |
| H* | 3.02±0.25 | 2.82±0.19 | 1.47±0.36 | 3.02±0.21 | 1.86±0.28 | |
| Е | 0.77±0.04 | 0.75±0.03 | 0.50±0.09 | 0.79±0.02 | 0.52±0.07 | |
| 1D | 0.89±0.05 | 0.87±0.03 | 0.59±0.12 | 0.91±0.02 | 0.64±0.09 | |
| 1/D | 13.08±2.53 | 9.72±2.24 | 3.87±1.34 | 12.70±2.76 | 3.82±1.10 | |
| DMg | 8.81±1.32 | 7.53±0.93 | 3.17±0.70 | 8.46±1.46 | 6.09±1.23 | |
| DMn | 2.57±0.17 | 2.65±0.29 | 1.22±0.25 | 2.85±0.23 | 1.80±0.20 | |
Наличие же супердоминантных видов снижает видовое разнообразие. На зарастающем участке поля обнаружено максимальное число видов в раннелетний период, в дальнейшем же происходит резкий спад видового разнообразия, что объясняется не только биологическими особенностями жуков, но и присутствием супердоминантного вида Protapion fulvipes с высокой долей в сборах, являющегося достаточно многочисленным и политопным и в естественно-природных экосистемах (Дмитриева, 2005в), а также низкими параметрами полидоминантности сообщества. Вследствие происходящих сукцессионных процессов выравненность населения сообщества жуков также снижена. Наименее разнообразным и выравненным является сообщество долгоносикообразных жуков одиночных березово-липовых лесополос (табл. 3), что обусловлено численным преобладанием одного супердоминантного вида Betulapion simile – монофага на Betula pendula.
Экстремальные засушливые погодные условия существенно отразились на сообществах долгоносикообразных жуков с бедным видовым составом и монокультурой (березово-липовые лесополосы), а также на несформированной фауне Curculionoidea зарастающих участков поля у смешанного леса.

Рис.2. Распределение модельных групп биотопов в плоскости КДФ в зависимости от видового богатства, численного обилия и значений показателей биоразнообразия.
Дискриминантный анализ, проведенный по показателям биоразнообразия, видового богатства и численного обилия (рис. 2) показал статистически высоко значимые различия (Wilks' Lambda: 0.3153044, approx. F (28.906) = 12.21382, p < 0.00001) между сообществами модельных групп биотопов. В плоскости канонических дискриминантных функций (КДФ) отчетливо выделены две группы: разнотравные луга, сосновые лесополосы и околоводно-прибрежные биотопы составляют первую группу, березово-липовые лесополосы и зарастающие лесо-полевые участки объединены во вторую. Статистически значимо не отличаются между собой сообщества Curculionoidea сосновых лесополос и околоводно-прибрежных биотопов. Наибольшее сходство по изучаемым параметрам отмечено между разнотравными лугами и околоводно-прибрежными комплексами (расстояние Махаланобиса (MD2) в плоскости КДФ – 1.31, p=0.000008), наименьшее между березово-липовыми лесополосами и околоводно-прибрежными биотопами (MD2 = 10.05, p=0.0000001).
5.4. Численное обилие Curculionoidea
Согласно t-критерию Стьюдента наибольшие значения численного обилия в умеренно-теплом сезоне характерны для березово-липовых лесополос (115.80±20.64) и зарастающих участков поля у смешанного леса (93.06±11.63), что обусловлено массовым размножением супердоминантных видов. В засушливый период статистически значимо выше значения численного обилия на разнотравных лугах (52.43±8.08), что связано с высоким видовым богатством фауны Curculionoidea, и зарастающих участках поля у леса (58.48±9.57) в связи с массовостью супердоминантного вида P. fulvipes и его миграцией с травянистого на древесно-кустарниковый ярус. Наименьшие средние значения численного обилия отмечены для околоводно-прибрежных биотопов. В засушливый период статистически значимое снижение численного обилия отмечено на зарастающих участках поля у смешанного леса (tSt=2.20, p=0.046) и в березово-липовых лесополосах (tSt=3.20, p=0.009). Наибольшее варьирование численного обилия в умеренно-теплом сезоне выявлено в березово-липовых лесополосах (V=56.37%) и на опушке смешанного леса (V=49.98%), в которых изменение численности приурочено к циклу развития супердоминантного вида. Минимальные межгодовые значения коэффициента вариации численного обилия отмечены для разнотравных лугов (V=31.89%) и околоводно-прибрежных биотопов (V=32.33%) в силу полидоминантности и выравненности фауны сообщества жуков.



№1. №2. №3.


№4. №5.
Рис. 3. Соотношение средних значений численного обилия и видового богатства Curculionoidea в модельных группах биотопов в 2010 г.
Сезонная динамика видового богатства Curculionoidea характеризуется согласно t-критерию Стьюдента статистически значимо высокими значениями в весенне-раннелетний период. В августе наблюдается снижение видового богатства в силу биологических особенностей жуков и погодных условий. В сентябре в большинстве биотопов отмечено повышение числа видов, что связано с выходом нового поколения имаго, но данный пик значительно ниже весенне-летнего по причине того, что в конце сезона кошение и вылавливание насекомых по высокому травостою сильно затруднено (рис. 3).
В динамике численного обилия на разнотравных лугах отмечено 2 пика: максимальный в июне, связанный с активным питанием и размножением видов после зимовки, второй пик в конце вегетационного периода, обусловленный появлением особей нового поколения. В остальных исследованных группах биотопов, несмотря на осенний пик, наблюдаемый в видовом богатстве, пик численного обилия в конце сезона не отмечен. Выявлено, что изменение численного обилия на разнотравных лугах положительно коррелирует (r=0.99, p<0.05; r=0.98, p<0.05) с изменением видового богатства в данной группе биотопов в течение всех изученных сезонов, в остальных биотопах достоверной корреляции не выявлено.
Глава 6. Видовая структура и пространственное распределение Curculionoidea в модельных группах биотопов
1) Разнотравные луга. Супердоминант S. suturalis (VI баллов по логарифмической шкале численного обилия). Доминанты: Protapion apricans (VI баллов по логарифмической шкале численного обилия), P. fulvipes (VI), Zacladus geranii (V), Sitona inops (V), субдоминанты: Pseudoperapion brevirostre (IV), Eutrichapion viciae (V), Gymnetron pascuorum (V), Phyllobius (Pterygorrhynchus) contemptus (V), S. lineatus (V). К оригинальным видам относятся: Pseudomechoris aethiops, Squamapion samarense, Sitophilus oryzae, Notaris rhamni, G. labile, Tychius quinquepunctatus, T. sharpi, Bagous lutulentus, Ceutorhynchus rapae, Datonychus arquata, Rhinoncus inconspectus, Ph. (Pterygorrhynchus) maculicornis, Parafoucartia (Foucartia) squamulata, Sciaphobus rubi, S. puncticollis, Magdalis frontalis.
2) Сосновые лесополосы. Супердоминанты P. fulvipes (VI), преобладающий по численному обилию и в естественных биоценозах (Дмитриева, 2005в), а также Ph. (Pterygorrhynchus) contemptus (VI), составляющие 30% сборов. Доминанты: P. apricans (V), Z. geranii (V), субдоминанты: Ceratapion onopordi (IV), Taeniapion urticarium (IV), Nedyus quadrimaculatus (V), Ph. (Metaphyllobius) pomaceus pomaceus (IV), S. inops (IV), S. suturalis (IV), Limobius borealis (V). Оригинальные виды: Perapion oblongum, Pseudostenapion simum, N. acridulus, Cionus scrophulariae, Mecinus janthinus, C. barbareae, C. sisymbrii, Coryssomerus capucinus, Omiamima mollina, S. waterhousei, L. albomarginatus.
3) Березово-липовые лесополосы. Супердоминантами выступают Betulapion simile – монофаг на Betula pendula, численное преобладание которого резко выражено (45.60%, VII баллов), и S. inops (12.63%, VI). Доминанты: Stenopterapion tenue (V), Polydrusus (Polydrusus) fulvicornis (V), субдоминанты: Deporaus betulae (IV), P. filirostre (IV), P. fulvipes (V), Orchestes rusci (IV). Оригинальные виды: D. mannerheimii, E. vorax, Curculio rubidus, C. sulcicollis, Coeliodinus rubicundus, Polydrusus (Eustolus) flavipes, Magdalis carbonaria.
4) Околоводно-прибрежные биотопы. Супердоминант P. fulvipes, доля которого в сборах составляет 22.40% (VI). Доминант: P. apricans (V), субдоминанты: E. viciae (IV), Catapion seniculus (V), P. trifolii (V), P. varipes (V), R. bruchoides (V), S. inops (V), S. suturalis (V). Оригинальные виды: Tropideres dorsalis, Temnocerus tomentosus, Kalcapion pallipes, S. vicinum, Cyanapion gnarum, Acalyptus carpini, A. sericeus, Ellescus infirmus, Isochnus foliorum, Rhamphus pulicarius, B. glabrirostris, B. scanicus, Aulacobaris lepidii, C. ignitus, C. roberti, Microplontus millefolii, Trichosirocalus barnevillei, Pelenomus quadrituberculatus, R. bruchoides, Larinus sturnus, L. flavescens, Cyphocleonus dealbatus.
5) Зарастающие участки поля вблизи сосново-широколиственного леса. Супердоминант (VIII) P. fulvipes с очень высокой долей в сборах (60.30%). Доминант P. apricans (V); субдоминанты: P. trifolii (V), P. varipes (IV), T. stephensi (V), L. obtusus (V). Оригинальные виды: Anthribus nebulosus, Apoderus coryli, C. alcyoneum, Oxystoma opeticum, O. pomonae, Synapion ebeninum, C. nucum, C. venosus, Dorytomus taeniatus, O. signifer, C. puncticollis, Brachysomus (Brachysomus) echinatus, S. languidus, Cathormiocerus aristatus, Hypera plantaginis, H. viciae.
Виды доминантного комплекса распределены в пределах групп биотопов агрегированно. В целом, среди выявленных видов преобладает агрегированное и регулярное пространственное распределение, характерное преимущественно для малочисленных редких видов. Высокие показатели средней плотности и встречаемости отмечены для доминантных видов. В засушливый сезон структура доминантного комплекса модельных биотопов несколько отлична.

Рис. 4. Дендрограмма фаунистического сходства модельных групп биотопов по индексу Чекановского-Съеренсена.
Сравнение видового состава Curculionoidea модельных групп биотопов показало наибольшее фаунистическое сходство между сосновыми лесополосами и разнотравными лугами (рис. 4), для которых отмечено максимальное сходство флористического состава (ICS=0.65). Кластерный анализ позволил выявить 3 комплекса биотопов со сходным видовым составом Curculionoidea.
Глава 7. Экологическое разнообразие фауны Curculionoidea агроэкосистемы
Изучение экологической структуры фауны Curculionoidea при адаптивно-ландшафтном земледелии способствует пониманию процессов формирования устойчивых агроэкосистем, отражает специфику изменчивости агроландшафтных комплексов и их динамику в ходе антропогенной трансформации, что дает возможность использовать данную группу жесткокрылых при мониторинге и индикации современного состояния агроценоза.
7.1. Распределение по фитобионтным группам
В ходе анализа приуроченности к определенным жизненным формам растений использовалась классификация фитобионтных групп, предложенная Ануфриевым и Кирилловой (1998).
Нами выделено 5 основных и 8 переходных фитобионтных групп: 1) дендробионты – обитатели древесного яруса, 2) тамнобионты – обитатели кустарникого яруса, 3) хамебионты – обитатели полукустарников и кустарничков, 4) хортобионты – виды, приуроченные к травянистому ярусу, 5) гидатобионты – обитатели водных растений. Гидатобионты и гидатохортобионты практически не встречаются в типичных агроценозах. Создание системы искусственных водоемов и минимизация антропогенного воздействия на прибрежные участки способствовало формированию околоводной фауны Curculionoidea в агроэкосистеме при адаптивно-ландшафтном земледелии.
Преобладают хортобионты, значительно меньше дендробионтов и дендротамнобионтов (табл. 4). Среди хортобионтов 64 вида (43.24%) развиваются на многолетниках, 28 видов (18.92%) – на однолетниках и многолетниках, 19 видов (12.84%) – на двулетниках и многолетниках, 18 видов (12.16%) – на одно- дву- и многолетниках, 8 видов (5.41%) – на двулетниках, 7 видов (4.73%) – на однолетниках, 4 вида (2.70%) – на однолетниках и двулетниках. Следовательно, основу фауны куркулионоидных жуков составляют хортобионты, связанные с многолетниками, что свидетельствует о ее сформированности и стабильной структуре.
Полученные данные согласуются с известными литературными сведениями, приведенными для природных сообществ Curculionoidea (Дмитриева, 2006; Хрисанова, 2006а). Широкий спектр фитобионтных групп Curculionoidea агроэкосистемы обусловлен разнообразием жизненных форм произрастающих в агроэкосистеме растений и ландшафтными особенностями территории, способствующие формированию различных микроклиматических условий. В отличие от природных сообществ и агроэкосистемы в условиях адаптивно-ландшафтного земледелия в фауне долгоносикообразных жуков агроценозов при традиционном земледелии доля хортобионтов значительно выше, дендробионты и дендротамнобионты почти не представлены, наименее разнообразен фитобионтный комплекс.
Таблица 4
Распределение Curculionoidea агроэкосистемы по фитобионтным группам
| Фитобионтная группа | Число видов | % | sp% |
| Дендробионт хвойных | 5 | 2.36 | 1.04 |
| Дендробионт лиственных | 11 | 5.19 | 1.52 |
| Дендротамнобионт лиственных | 28 | 13.21 | 2.33 |
| Дендротамнохамебионт | 2 | 0.94 | 0.66 |
| Дендротамнохамехортобионт | 5 | 2.36 | 1.04 |
| Дендротамнохортобионт | 1 | 0.47 | 0.47 |
| Тамнобионт лиственных | 1 | 0.47 | 0.47 |
| Тамнохамехортобионт | 2 | 0.94 | 0.66 |
| Хамебионт | 1 | 0.47 | 0.47 |
| Хамехортобионт | 3 | 1.42 | 0.81 |
| Хортобионт | 148 | 69.81 | 3.15 |
| Гидатобионт | 1 | 0.47 | 0.47 |
| Гидатохортобионт | 4 | 1.89 | 0.94 |
| Всего: | 212 | 100.00 |
7.2. Трофическая структура фауны Curculionoidea
Пищевая специализация. В ходе изучения широты трофического спектра были использованы собственные наблюдения и литературные данные (Исаев, 1994, 2000; Исаев и др., 2004; Дмитриева, 2005в; Хрисанова, 2005, 2006 б; Дедюхин, 2007; Исаев, 2007; Дедюхин, 2009, 2010а, б; Legalov, 2010; Дедюхин, 2011 и др.). По характеру предпочитаемого пищевого субстрата среди выявленных нами видов долгоносикообразных жуков в пределах агроэкосистемы выделены следующие группы:
1) детритофаги (характерно питание мертвыми, разлагающимися растительными остатками) – малочисленная группа, к которой отнесено 3 вида: Omiamima mollina, Brachysomus (Brachysomus) echinatus, Cathormiocerus aristatus (Исаев, 2007);
2) зоомикофаги (кокцидомикофаги) – имаго Anthribus nebulosus охотится преимущественно на ложнощитовку еловую Physokermes piceae, личинки питаются щитовками, развивающимися на деревьях, также обнаружены на грибе Apiosporium pinophilum (Никитский и др., 1996);
3) зооцицидофаги – виды-инквилины Archarius crux, A. salicivorus, личинки которых развиваются в галлах пилильщиков из родов Pontania, Euura (Hymenoptera: Tenthredinidae) на листьях ив (Опанасенко, 1976). Личинки Melanapion minimum – инквилины в листовых галлах пилильщиков P. femoralis, P. proxima, P. bella, Amauronematus viduatus из семейства Tenthredinidae, личинки Polydrusus (Eurodrusus) cervinus являются инквилинами в листовых галлах, образуемых клещиками из рода Eriophyes;
4) фитофаги – многочисленная группа (211 видов среди имаго и 207 среди личинок), включающая растительноядных Curculionoidea.
Принимая за основу существующие классификации фитофагов по широте трофического спектра (Емельянов, 1964; Исаев, 1994) нами выделены следующие группы: монофаги – 39 видов, 18.48%; олигофаги: истинные узкие олигофаги (57 видов, 27.01%), узкодизъюнктивные олигофаги – виды, развивающиеся на растениях из разных родов одного семейства, часто не близких между собой, но не более чем на двух-трех (39 видов, 18.48%), широкие олигофаги – 41 вид, 19.43%, широкодизъюнктивные олигофаги – развиваются на немногих родах растений из разных не близких в систематическом отношении между собой семейств (10 видов, 4.74%); полифаги: узкие полифаги – виды, развивающиеся на растениях разных преимущественно близких семейств (7 видов, 3.32%), широкие полифаги – виды, развивающиеся на растениях из разных семейств, в том числе и на хвойных породах (18 видов, 8.53%).

Рис. 5. Варьирование числа видов Curculionoidea различного трофического спектра в агроэкосистеме.
В фауне долгоносикообразных жуков рассматриваемой агроэкосистемы среди фитофагов преобладают олигофаги (147 видов, 69.67%). Незначительна доля полифагов (25 видов, 11.85%). Основу исследуемой фауны Curculionoidea преимущественно составляют виды с узкой пищевой специализацией: 45.50% (96 видов) видов являются монофагами и узкими олигофагами. В наибольшей степени варьирует число узких и широких олигофагов (рис. 5).
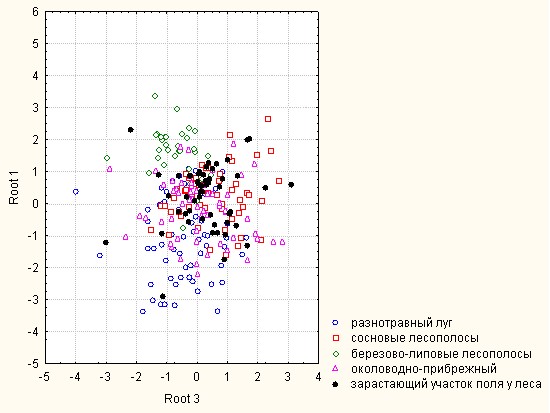
Рис. 6. Распределение модельных групп биотопов в плоскости КДФ в зависимости от трофического спектра видов сообществ долгоносикообразных жуков.
Модельные группы биотопов статистически значимо (Wilks' Lambda: 0.3991313, approx. F (28.906) = 9.391806, p<0.00001) различаются по трофической структуре сообществ долгоносикообразных жуков, особенно сообщества Curculionoidea березово-липовых лесополос и разнотравных лугов (p=0.0000001) (рис. 6).
Сложившаяся трофическая структура фауны Curculionoidea агроэкосистемы свидетельствует об ее устойчивости и зрелости. Интересно отметить, что для фауны долгоносикообразных жуков природных комплексов (Дмитриева, 2006; Хрисанова, 2006а) характерно сходное соотношение основных групп по пищевой специализации – монофагов, олигофагов и полифагов, что еще раз подчеркивает близость фауны Curculionoidea агроэкосистемы при адаптивно-ландшафтном земледелии с природными биоценозами.
Анализ приуроченности долгоносикообразных жуков-фитофагов к органам кормовых растений проводился с учетом классификации, предложенной Хрисановой М.А. (2006 а, б). Распределение имаго и личинок Curculionoidea по органам кормовых растений отличается и по широте трофического спектра и по предпочитаемым органам растений. По трофическим связям имаго Curculionoidea с органами растений нами выделено 22 группы, для личинок данный показатель значительно больше 35 трофических групп (табл. 5).
При этом для большинства имаго долгоносикообразных жуков, ведущих открытый образ жизни, в качестве пищевого объекта чаще всего выступают листья (45.02%), а также ткани цветков растений (7.11% антофагов и 18.96% антофиллофагов). В спектре приуроченности имаго Curculionoidea наиболее разнообразна группа видов, связанных с почками растений. Широта спектра питания имаго обусловлена особенностями жизненного цикла развития растения: весной с массовым выходом имаго (апрель-май) из мест зимовок большинство растений находится в стадии почкообразования и бутонизации, поэтому потенциальный пищевой ресурс имаго включает главным образом листья, почки и соцветия растений. На личиночной стадии Curculionoidea имеют более широкий трофический спектр приуроченности к органам растений. Среди личинок встречаются минеры листьев, галлообразователи (листовые, стеблевые, корневые).
Выявленные различия, существующие между имаго и личиночной стадией, способствуют расхождению экологических ниш, что положительно сказывается на видовом богатстве фауны Curculionoidea. В целом, трофическая структура долгоносикообразных жуков при адаптивной эколого-ландшафтной беспестицидной системе земледелия отличается многообразием, что обуславливает биологическое разнообразие Curculionoidea в агроэкосистеме.
Таблица 5
Распределение имаго и личинок Curculionoidea по трофической приуроченности к органам растений
| Распределение имаго по органам растений | Число видов | % | sp% | Распределение личинок по органам растений | Число видов | % | sp% |
| Поллиноантокарпофаг | 1 | 0.47 | 0.47 | Поллинофаг | 1 | 0.48 | 0.48 |
| Антофаг | 15 | 7.11 | 1.77 | Антофаг | 14 | 6.76 | 1.74 |
| Антосеменофиллофаг | 1 | 0.47 | 0.47 | Антосеменофаг | 8 | 3.86 | 1.34 |
| Антокарпофиллокаулофаг | 1 | 0.47 | 0.47 | Антокарпофаг | 2 | 0.97 | 0.68 |
| Антофиллофаг | 40 | 18.96 | 2.7 | Антокарпофиллофаг | 1 | 0.48 | 0.48 |
| Антофиллокаулофаг | 8 | 3.79 | 1.31 | Антофиллофаг | 7 | 3.38 | 1.26 |
| Семенофаг | 1 | 0.47 | 0.47 | Антокаулофаг | 1 | 0.48 | 0.48 |
| Карпофиллокаулофаг | 2 | 0.96 | 0.67 | Антофиллокаулофаг | 1 | 0.48 | 0.48 |
| Геммоантосеменофаг | 1 | 0.47 | 0.47 | Антокаулоцицидофаг | 1 | 0.48 | 0.48 |
| Геммоантосеменофиллофаг | 1 | 0.47 | 0.47 | Антоцицидофаг | 1 | 0.48 | 0.48 |
| Геммоантокарпофиллофаг | 2 | 0.96 | 0.67 | Анто-филлокаулоцицидофаг | 1 | 0.48 | 0.48 |
| Геммоантофиллофаг | 5 | 2.37 | 1.05 | Семенофаг | 26 | 12.56 | 2.3 |
| Геммокарпофиллофаг | 1 | 0.47 | 0.47 | Карпофаг | 5 | 2.42 | 1.07 |
| Геммофиллофаг | 11 | 5.21 | 1.53 | Карпосеменофаг | 3 | 1.45 | 0.83 |
| Геммофиллокаулофаг | 1 | 0.47 | 0.47 | Карпосеменокаулофаг | 1 | 0.48 | 0.48 |
| Геммофиллоксилофаг | 1 | 0.47 | 0.47 | Геммофаг | 2 | 0.97 | 0.68 |
| Филлофаг | 95 | 45.02 | 3.43 | Геммоантофаг | 1 | 0.48 | 0.48 |
| Филлокаулофаг | 17 | 8.06 | 1.87 | Геммоантофиллофаг | 2 | 0.97 | 0.68 |
| Филлоксилофаг | 1 | 0.47 | 0.47 | Геммоантофиллокаулофаг | 1 | 0.48 | 0.48 |
| Каулофаг | 2 | 0.96 | 0.67 | Геммофиллофаг | 2 | 0.97 | 0.68 |
| Ксилофаг | 3 | 1.43 | 0.82 | Филлофаг | 18 | 8.70 | 1.96 |
| Ризофаг | 1 | 0.47 | 0.47 | Филлокаулофаг | 1 | 0.48 | 0.48 |
| Всего: | 211 | 100.00 | Филлокаулоризофаг | 2 | 0.97 | 0.68 | |
| Филлоризофаг | 1 | 0.48 | 0.48 | ||||
| Филлоцицидокаулофаг | 1 | 0.48 | 0.48 | ||||
| Филлокаулоцицидофаг | 1 | 0.48 | 0.48 | ||||
| Филлокаулоризоцицидофаг | 2 | 0.97 | 0.68 | ||||
| Каулофаг | 23 | 11.12 | 2.19 | ||||
| Каулоризофаг | 9 | 4.35 | 1.42 | ||||
| Каулоцицидофаг | 6 | 2.90 | 1.17 | ||||
| Каулоризоцицидофаг | 2 | 0.97 | 0.68 | ||||
| Ксилофаг | 7 | 3.38 | 1.26 | ||||
| Ризофаг | 31 | 14.98 | 2.48 | ||||
| Ризофаг, клубеньки бобовых | 12 | 5.80 | 1.62 | ||||
| Ризоцицидофаг | 4 | 1.93 | 0.96 | ||||
| Не установлено | 6 | 2.90 | 1.17 | ||||
| Всего: | 207 | 100.00 | |||||
7.3. Биотопическое распределение Curculionoidea
Оценка биотопической приуроченности видов куркулионоидных жесткокрылых проводилась на основе показателя встречаемости. По преимущественной биотопической приуроченности имаго нами выделены следующие основные группы: 1) лесная; 2) луговая; 3) болотная; 4) околоводная. По спектру заселяемых стаций в фауне Curculionoidea рассматриваемой агроэкосистемы встречаются также переходные биотопические группы: лугово-лесные, лугово-степные, болотно-лесные, околоводно-лесные, околоводно-луговые и т.д. (табл. 6).
В целом, биотопический спектр изучаемой фауны Curculionoidea включает 15 групп. Заметно преобладание комплекса лугово-лесных, околоводно-лугово-лесных, лесных и луговых видов, которые суммарно составляют около 75% фауны Curculionoidea агроэкосистемы.
Таблица 6
Распределение долгоносикообразных жесткокрылых агроэкосистемы в условиях адаптивно-ландшафтного земледелия по биотопическим группам
| Биотопическая группа | Число видов | % | sp% |
| Лесная | 29 | 13.49 | 2.33 |
| Луговая | 23 | 10.70 | 2.11 |
| Лугово-лесная | 75 | 34.88 | 3.25 |
| Лугово-степная | 7 | 3.26 | 1.21 |
| Лугово-лесостепная | 1 | 0.46 | 0.46 |
| Болотно-лесная | 4 | 1.86 | 0.92 |
| Болотно-околоводно-лесная | 8 | 3.72 | 1.29 |
| Болотно-околоводная | 9 | 4.19 | 1.37 |
| Околоводная | 5 | 2.33 | 1.03 |
| Околоводно-лесная | 7 | 3.26 | 1.21 |
| Околоводно-луговая | 6 | 2.79 | 1.12 |
| Околоводно-лугово-лесная | 34 | 15.81 | 2.49 |
| Околоводно-болотно-луговая | 4 | 1.86 | 0.92 |
| Околоводно-болотно-лугово-лесная | 2 | 0.93 | 0.65 |
| Околоводно-лугово-лесостепная | 1 | 0.46 | 0.46 |
| Всего: | 215 | 100.00 |
Наиболее варьирует число лугово-лесных и околоводно-лугово-лесных видов, составляющих ядро фауны всех групп биотопов. Полученные данные объясняются особенностями физико-географического расположения территории исследования, а также ландшафтными особенностями (преобладание балок, мезопонижений и т.д.), внедренным комплексом лесомелиоративных и гидромелиоративных мероприятий в пределах агроэкосистемы, которые в свою очередь способствовали формированию различных условий обитания, разнообразных луговых ассоциаций и многочисленных экотонных участков.

Рис. 7. Распределение модельных участков в плоскости КДФ в зависимости от биотопического спектра видов сообществ долгоносикообразных жуков.
Дискриминантный анализ выявил статистически значимые различия (Wilks' Lambda: 0.16725, approx. F (60.950)=9.2081, p=0.00001) в биотопической структуре сообществ Curculionoidea модельных участков. Наиболее различна структура сообществ березово-липовых лесополос и разнотравных лугов (MD2 = 18.97, p=0.0000001), а также березово-липовых и сосновых лесополос (MD2 = 13.91, p=0.0000001) (рис. 7).
Глава 8. Синэкологические аспекты биоразнообразия Curculionoidea
Поскольку большинство видов долгоносикообразных жуков является узкоспециализированными облигатными фитофагами, то их распространение зависит прежде всего от видового состава и видового богатства растительного покрова модельных групп биотопов, о чем свидетельствуют результаты проведенного корреляционного анализа. Выявлена сильная линейная положительная корреляция (r=0.95, p<0.05) между числом видов Curculionoidea и числом произрастающих видов растений в исследуемых группах биотопов. Кроме того, зафиксирована сильная линейная положительная корреляция (p<0.05) между числом произрастающих видов растений и показателями биоразнообразия Curculionoidea в модельных группах биотопах: индексом Маргалефа (r=0.98), Менхиника (r=0.95) и Шеннона (r=0.94) в умеренно-теплом сезоне.
Анализ трофических связей изучаемой фауны долгоносикообразных жуков позволил выявить приуроченность к более 50 семействам растений из 3 отделов: Хвощеобразные (Equisetophyta), Соснообразные (Pinophyta), или Голосеменные (Gymnospermae) и Магнолиообразные (Magnoliophyta), или Покрытосеменные (Angiospermae). Наибольшее число видов Curculionoidea отмечено на растениях семейств Fabaceae (62 вида), Salicaceae (33), Asteraceae (32), Betulaceae (28), Rosaceae (24), Polygonaceae (22), Fagaceae (18), Brassicaceae (14), Lamiaceae (8), Tiliaceae, Scrophulariaceae (по 7 видов), Pinaceae, Chenopodiaceae, Ulmaceae и Urticaceae (по 6 видов), составляющих основу флористического комплекса агроэкосистемы. По приуроченности к родам растений наиболее богата фауна представителей Trifolium L. (36 видов), Vicia L. (32), Medicago L. (30), Salix L. (29), Lotus L. (27), Betula L. (26), Lathyrus L. (23), Populus L. (21), Amoria C. Presl (20), Rumex L. и Quercus L. (по 19), Corylus L. и Melilotus Mill. (по 18), Alnus Mill. (17), Pinus L. и Rubus L. (по 15), Rosa L. и Malus Mill. (по 14), Pyrus L., Padus Mill., Cirsium Mill., Centaurea L. (по 12). В целом, трофический спектр фауны Curculionoidea изучаемой агроэкосистемы при адаптивно-ландшафтном земледелии включает более 190 родов растений. В пределах модельных групп биотопов нами выявлено 203 вида сосудистых растений из 140 родов, 51 семейства, 42 порядков, 16 классов и 4 отделов, с учетом маршрутных сборов 233 вида из 158 родов. Три вида (яблоня лесная Malus sylvestris Mill., 1768, купальница европейская Trollius europaeus L., 1753, горечавка крестовидная Gentiana cruciata L., 1753) включены в Красную книгу ЧР (2001). Видовой состав Curculionoidea более чем на 80% отражает флористический состав изучаемой территории, что позволяет использовать долгоносикообразных жуков при проведении мониторинговых исследований.
В фауне Curculionoidea агроэкосистемы преобладают виды, развивающиеся преимущественно на растениях одного семейства (178 видов, 82.79%), значительно реже на двух (12 видов, 5.58%) и трех (7 видов, 3.26%), что скорее всего связано с биохимической коэволюцией растений и насекомых-фитофагов, приводящей к развитию защитных механизмов у кормового растения и противодействующих механизмов у насекомого (Фитофаги в растительных сообществах, 1980).
Кормовые растения характеризуются синтезом вторичных веществ, часто ядовитых для фитофагов, в организме которых возможна детоксикация лишь определенного ряда химических веществ, что приводит к ограничению потенциального пищевого спектра насекомых и выработке приспособлений к питанию на предпочитаемом ряде растений. В то же время известно, что химический состав растений зависит от условий и адаптивных способностей растений. В связи с этим мы предположили, что трофический спектр долгоносикообразных жуков-фитофагов в биоценозе определенным образом зависит от типа адаптивной стратегии произрастающих в нем растений.
Основу флоры агроэкосистемы составляют наземные, автотрофные, летнезеленые лугово-степные и водно-болотные травянистые поликарпики, гемикриптофиты. По отношению к влажности преобладают мезофиты, по отношению к питанию мезотрофы и мезоэвтрофы, что соответствует полученным результатам по агрохимическому исследованию почв. Преобладающим типом стратегии является конкурентный (виолент) С-тип (37.87%), второе место занимают виды со стратегией с сочетанием свойств первичных типов стратегий CSR (26.38%) и третье – виды с типом стратегии с сочетанием свойств конкурентности и стресс-толерантности CS (14.04%). Доминирование виолентов свидетельствует о благоприятных абиотических условиях и отсутствии (минимум присутствии) нарушений на изучаемой территории, а также об устойчивости фитоценоза.
Выявлена значимая положительная корреляционная связь между числом узких олигофагов (rS=0.90, p=0.04), широкодизъюнктивных олигофагов (rS=0.97, p=0.005), широких полифагов Curculionoidea (rS=0.90, p=0.04) и числом видов растений конкурентного и вторичного CSR типа адаптивной стратегии в модельных группах биотопов. К виолентам относятся многие растения- эдификаторы с широкой экологической нишей (деревья, кустарники, травы мезоморфного облика с широким простиранием в пространстве), в особенности в благоприятных, мезофильных местообитаниях, способные при достаточно высоком уровне обеспечения их светом и элементами минерального питания полностью использовать имеющиеся ресурсы (Раменский, 1971; Миркин, 1985; Миркин и др., 2000; Работнов, 1985; Усманов и др., 2001). Вторичный тип стратегии CSR составляют все луговые растения (Миркин и др., 2000). Следовательно, виды-эдификаторы с конкурентным типом эколого-фитоценотической стратегии и большинство луговых растений со вторичным CSR типом стратегии создают кормовую базу преимущественно для долгоносикообразных жуков-фитофагов со средним и широким трофическими спектрами.
Глава 9. Хозяйственное значение долгоносикобразных жуков агроэкосистемы
9.1. Основы взаимоотношений фитофага и кормового растения
Коэволюция растений и насекомых-фитофагов способствовала формированию комплекса адаптивных реакций, физиологических, биохимических и морфологических взаимоприспособлений на видовом уровне у каждого из партнеров по трофической связи, что привело к тому, что наносимый фитофагами вред компенсируется растением, вторичные вещества которого ограничивают размножение и питание насекомого.
9.2. Комплекс полезных видов Curculionoidea в агроэкосистеме
Наиболее перспективными биологическими агентами в борьбе со сорными растениями являются представители прогрессивных семейств Apionidae, Nanophyidae и Curculionidae (Коротяев, 2012). В мировой практике имеются примеры успешного использования ряда видов. В частности, Ceutorhynchus assimilis, способный нанести вред капустным в нашей зоне, в Северной Америке используется для борьбы с евразийским инвазивным сорняком Cardaria draba (Fumanal et al., 2004), являющимся массовым и в США, где в качестве потенциального биологического агента используют C. cardariae (Коротяев, 2012). C. roberti используется в С.Америке для сокращения распространения сорного растения Alliaria petiolata (Davis et al., 2006). Hadroplontus litura применяется в комбинированной биологической борьбе против канадского чертополоха (Cirsium arvense), широко распространенным сорным растением в США, Канаде, Северной Европе и Новой Зеландии (Sciegienka, 2011). Эффективность использования Mecinus janthinus, повреждающего и развивающегося в стеблях кормовых сорных растений из рода Linaria в качестве биологического агента в Канаде, научно доказана и подтверждена. Долгоносик был ввезен в Канаду в 1991 году из Южной Европы и юга России, с тех пор проводились неоднократные наблюдения и оценка эффективности его использования (Crawley, 1989; Syrett et al, 1996; McCiay, De Clerck-Floate, 2002). Ведутся активные исследования по изучению биологических особенностей Larinus obtusus, L. sturnus, L. planus, L. turbinatus с целью их использования в борьбе с сорными растениями из родов Carduus и Cirsium (McClay,1990; Groppe, 1992; Louda, O’Brien, 2002; Skuhrovec, Gosik, 2011). Ceratapion onopordi используется и официально утвержден в качестве биологического агента против различных видов чертополохов в Новой Зеландии (Import and release Ceratapion onopordi, 1999). C. basicorne, обитающий в Европе и Западной Азии, в настоящее время является перспективным классическим биологическим агентом в регуляции распространения Centaurea solstitialis в США (Smith, 2007).
9.3. Вредоносные виды Curculionoidea в агроэкосистеме
Среди экономически важных Curculionoidea агроэкосистемы 20 видов являются фактическими вредителями и 20 видов – потенциальными, что составляет 18.43% выявленной фауны. 1) Вредители зернобобовых культур и многолетних трав: Eutrichapion ervi, E. viciae, Oxystoma cerdo, O. craccae, Catapion seniculus, Protapion apricans, P. fulvipes, P. trifolii, Tychius aureolus, T. medicaginis, T. quinquepunctatus, T. picirostris, Otiorhynchus (Cryphiphorus) ligustici, Sitona cylindricollis, S. hispidulus, S. inops, S. lepidus, S. lineatus, S. longulus, S. macularius, Hypera nigrirostris. 2) Вредители плодовых и ягодных культур: Tatianaerhynchites aequatus, Anthonomus pomorum, A. rectirostris, A. rubi, Phyllobius (Phyllobius) pyri, к стволовым вредителям относятся Magdalis ruficornis, Scolytus mali, S. rugulosus. 3) Вредители гречишных культур: Rhinoncus bruchoides, R. pericarpius.4) Вредители крестоцветных культур: Ceutorhynchus assimilis, C. rapae, C. typhae. 5) Вредители зерновых запасов: Sitophilus oryzae. 6) Вредители древесных насаждений: Cryptorhynchus lapathi, Magdalis carbonaria, M. duplicata. 7) Многоядные вредители: Tanymecus palliatus – типичный полифаг.
Анализ литературных сведений показал, что регуляция численности вредоносных куркулионоидных жуков возможна не только агротехническими приемами, но главным образом благодаря действию естественных механизмов. Местная орнитофауна, энтомофаги, внутри- и межвидовая конкуренция за общий кормовой субстрат и места обитания способствуют снижению численности вредоносных Curculionoidea. Кроме того, некоторые из этих видов на всех стадиях развития поражаются грибами, различными болезнями, паразитами из различных таксонов.
При рассмотрении распределения выявленных видов вредоносных Curculionoidea по численному обилию согласно составленной логарифмической шкале, приходим к выводу, что 90% из них (36 видов) относятся к 1-5 классу по степени обилия, что соответствует редким, малочисленным и среднеобильным видам, причем преобладают единичные и малочисленные виды (47.50%). Среди многочисленных отмечено 4 вида. Супердоминантом является политопный, приуроченный к древесно-кустарниковому ярусу вид P. fulvipes. Исходя из полученных данных, можно заключить, что большинство видов Curculionoidea выявленного вредоносного комплекса не оказывают ощутимого вреда на культурные фитоценозы. Отмеченные многочисленные виды трофически связаны с бобовыми растениями, преобладающими по видовому разнообразию и проективному покрытию в изучаемых агроландшафтах.
ВЫВОДЫ
1) Таксономическое разнообразие долгоносикообразных жуков агроэкосистемы при адаптивно-ландшафтном земледелии сопоставимо с таковым природоохранных территорий и представлено 215 видами из 99 родов, 45 триб, 15 подсемейств и 8 семейств. 189 видов впервые указаны для фауны района исследования, 4 вида для Чувашской Республики. Видовое богатство фауны Curculionidae характеризуется следующим рядом: природно-естественные комплексы адаптивно-ландшафтное земледелие > традиционное земледелие.
2) Наибольшим видовым разнообразием, оригинальностью видового богатства, полидоминантной структурой и выравненностью сообщества долгоносикообразных жуков агроэкосистемы при адаптивно-ландшафтном земледелии характеризуются разнотравные мезофитные луга и экотонные прибрежно-околоводные и лесо-луговые участки. Высокий уровень видового богатства долгоносикообразных жуков в биотопах отмечен в весенне-раннелетний период.
3) По приуроченности к жизненным формам растений в фауне преобладают хортобионтные долгоносикообразные жесткокрылые; по широте трофического спектра олигофаги; большинство имаго Curculionoidea являются филлофагами, личинки преимущественно приурочены к генеративным органам и корням растений.
4) Ядро фауны составляют лугово-лесные, лесные и луговые виды долгоносикообразных жуков, распределение которых в пространстве преимущественно агрегированное.
5) Видовое разнообразие Curculionoidea при адаптивно-ландшафтном земледелии определяется видовым богатством флористического комплекса и широким набором абиотических микроклиматических условий. Видовое богатство долгоносикообразных жуков-фитофагов со средним и широким трофическим спектром обусловлено разнообразием растений с конкурентным типом эколого-фитоценотической стратегии, а также вторичным CSR типом, преобладающими среди фитоценозов агроэкосистемы.
6) Среди важных в хозяйственном отношении видов долгоносикообразных жуков выделен комплекс полезных представителей, включающий энтомофагов, инквилинов и фитофагов сорных растений, а также ряд вредоносных Curculionoidea, в том числе и зерновых культур, составляющих в целом 18.43% выявленной фауны. Преобладают малочисленные и единичные вредоносные виды, регуляция численности большинства которых осуществляется местными энтомофагами и рациональными агротехническими приемами.
ПУБЛИКАЦИИ ПО ТЕМЕ ДИССЕРТАЦИИ:
Публикации в изданиях, рекомендованных ВАК
- Григорьян Б.Р., Николаева Т.Г., Сунгатуллина Л.М. Изменение биологических параметров почвенной экосистемы в агробиоценозах в условиях различных систем земледелия // Георесурсы (научно-технический журнал). – 2011. – №2(38). – С. 9-13. (0.88 п.л., автора – 0.8)
- Николаева Т.Г. Экологическая характеристика долгоносикообразных жуков (Coleoptera, Curculionoidea) агроэкосистем при адаптивно-ландшафтном земледелии на севере лесостепи Приволжской возвышенности // Ученые записки Казанского университета. Сер. Естественные науки. 2012. Т. 154, кн. 2. (принята к печати).
- Николаева Т.Г., Григорьян Б.Р. Качество и экологическая безопасность зерна при биологическом и ресурсосберегающем земледелии в агроландшафтах лесостепи Приволжского региона // Вестник Саратовского госагроуниверситета им. Н.И. Вавилова. – 2011. – №6. – С. 25-30. (0.70 п.л., автора – 0.95)
- Николаева Т.Г., Григорьян Б.Р., Сунгатуллина Л.М. Сохранение биоразнообразия и почвенного плодородия – основа устойчивого развития органического сельского хозяйства // Ученые записки Казанского университета. Сер. Естественные науки. – 2011. – Т. 153, кн. 1. – С. 136-151. (1.36 п.л., автора – 0.95)
Публикации в тематических сборниках и трудах конференций
- Егоров Л.В., Николаева Т.Г. О составе фауны долгоносикообразных жуков (Coleoptera, Curculionoidea) агроэкосистем при адаптивно-ландшафтном земледелии на севере лесостепи Приволжской возвышенности / Роль особо охраняемых природных территорий в сохранении биоразнообразия: Матер. III Междунар. науч.-практ. конф. (Чебоксары, 25-26 ноября 2010 г.) // Научные труды государственного природного заповедника "Присурский". Чебоксары-Атрат: КЛИО, 2010. Т. 24. С. 41-45. (0.75 п.л., автора – 0.5)
- Николаева Т.Г. К изучению биоразнообразия долгоносикообразных жесткокрылых (Coleoptera, Curculionidea) в условиях органического земледелия // Полевые и экспериментальные исследования биологических систем: Матер. III Всеросс. науч.-практ. конф. студентов, аспирантов и молодых учёных (Ишим, 18 марта 2010 г.) – Ишим: ИГПИ им. П.П.Ершова, 2010. – С. 74-77. (0.19 п.л.)
- Николаева Т.Г. Состояние компонентов агроэкосистемы «почвенно-биотический комплекс – культурные растения» в условиях биологического и ресурсосберегающего земледелия // Экологическое равновесие и устойчивое развитие территории: Матер. Междунар. науч.-практ. конф. (Пушкин, 30-31 марта 2010 г.) – СПб.: ЛГУ, 2010. – С. 193-196. (0.20 п.л.)
- Николаева Т.Г. Изучение состояния некоторых компонентов полевых агроэкосистем в условиях биологического и ресурсосберегающего земледелия // Тез. 14 Междунар. Пущинской школы-конференции молодых ученых «Биология – наука XXI века» (Пущино, 19-23 апреля 2010 г.). – Пущино, 2010. – Т.2.– С. 63-64. (0.12 п.л.)
- Николаева Т.Г. Особенности некоторых компонентов полевых агроэкосистем в условиях биологического и ресурсосберегающего земледелия // Тез. III Всеросс. с междунар. участием конгресса студентов и аспирантов-биологов «Симбиоз-Россия 2010» (Ниж. Новгород, 24-29 мая 2010 г.). – Ниж. Новгород: ННГУ, 2010. – С. 26. (0.12 п.л.)
- Николаева Т.Г. Таксономический аспект биоразнообразия долгоносикообразных жуков (Coleoptera, Curculionoidea) в условиях адаптивно-ландшафтной системы земледелия на севере лесостепи Приволжской возвышенности // Биоразнообразие: глобальные и региональные процессы: Матер. Всеросс. конф. молодых ученых (Улан-Удэ, 14-17 сентября 2010 г.). – Улан-Удэ: БНЦ СО РАН, 2010. – С. 58-60. (0.08 п.л.)
- Николаева Т.Г. Видовое разнообразие долгоносикообразных жесткокрылых (Coleoptera, Curculionoidea) агроэкосистем лесостепи Приволжской возвышенности при адаптивно-ландшафтной системе земледелия // Принципы и способы сохранения биоразнообразия: Матер. IV Всеросс. науч. конф. с междунар. участием (Йошкар-Ола, 22-26 сентября 2010 г.) – Йошкар-Ола: МарГУ, 2010. – С. 138-140. (0.35 п.л.)
- Николаева Т.Г. Биотопическая приуроченность долгоносикообразных жуков (Coleoptera, Curculionoidea) в агроценозах севера лесостепи Приволжской возвышенности в условиях адаптивной эколого-ландшафтной системы земледелия // Труды Казанского отделения РЭО. – 2010. (в печати).
- Николаева Т.Г. Особенности некоторых биотических компонент агроэкосистемы в условиях адаптивно-ландшафтного земледелия // Биология – наука XXI века: Сборник тезисов 15-ой Международной Пущинской школы-конференции молодых ученых (Пущино, 18-22 апреля 2011 г.). – Пущино, 2011. – С.252. (0.12 п.л.)
- Николаева Т.Г. Анализ трофической приуроченности долгоносикообразных жуков (Coleoptera, Curculionoidea) агроэкосистем севера лесостепи Приволжской возвышенности при адаптивно-ландшафтном земледелии // Ломоносов-2011: Материалы XVIII Международной научной конференции студентов, аспирантов и молодых ученых (Москва, МГУ, 11-15 апреля 2011 г.) / Отв. ред. А.И. Андреев, А.В. Андриянов, Е.А. Антипов, М.В. Чистякова. – [Электронный ресурс] — М.: МАКС Пресс, 2011. — 1 электрон. опт. диск (DVD-ROM); 12 см. - Систем. требования: ПК с процессором 486+; Windows 95; дисковод DVD-ROM; Adobe Acrobat Reader. – С.118. (0.03 п.л.)
- Николаева Т.Г. Трофическая структура долгоносикообразных жуков (Coleoptera, Curculionoidea) агроэкосистем при адаптивно-ландшафтном земледелии на севере лесостепи Приволжской возвышенности // ZOOCENOSIS–2011: Материалы VI Международной научной конференции «Биоразнообразие и роль животных в экосистемах» (Украина, Днепропетровск, 4-6 октября 2011 г.). – Днепропетровск: Изд-во ДНУ, 2011. – С.199-201. (0.25 п.л.)
- Николаева Т.Г. Синэкологические аспекты биоразнообразия долгоносикообразных жуков (Coleoptera, Curculionoidea) агроэкосистем при адаптивно-ландшафтном земледелии на севере лесостепи Приволжской возвышенности // Ломоносов-2012: Тезисы докладов XIX Международной научной конференции студентов, аспирантов и молодых ученых: секция "Биология" (9-13 апреля 2012 г., Москва, МГУ имени М.В. Ломоносова, биологический факультет) / Отв. ред. Е.Н. Темерова, Сост. А.И. Исаченко. М.: МАКС Пресс, 2012. С. 116-117. (0.03 п.л.)
- Николаева Т.Г., Егоров Л.В. Видовое богатство и численное обилие долгоносикообразных жуков (Coleoptera, Curculionoidea) в агроценозах севера лесостепи Приволжской возвышенности при адаптивно-ландшафтной системе земледелия // Устойчивость экосистем: теория и практика: Матер. докладов Всеросс. науч. конф. с междунар. участием (Чебоксары, 23 октября 2010 г.). – Чебоксары: Изд-во ООО «Листок», 2010. – Т. 1. – С. 31-35. (0.26 п.л., автора – 0.8)