

Создание новой экспрессионной системы на основе расшифровки механ измов экспрессии генов протеаз бацилл
ФГАОУВПО «Казанский (Приволжский) федеральный университет»
На правах рукописи
ТОЙМЕНЦЕВА АННА АЛЕКСАНДРОВНА
Создание новой экспрессионной системы на основе расшифровки механизмов экспрессии генов протеаз бацилл
03.02.03 – микробиология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Казань – 2012
Работа выполнена в лаборатории биосинтеза и биоинженерии ферментов кафедры микробиологии Института фундаментальной медицины и биологии ФГАОУВПО «Казанский (Приволжский) федеральный университет».
Научный руководитель: Доктор биологических наук, профессор
Шарипова Маргарита Рашидовна
Официальные оппоненты: Доктор ветеринарных наук, профессор
Профессор кафедры Казанской государственной академии ветеринарной медицины им. Н.Э. Баумана
Госманов Рауис Госманович
Кандидат биологических наук
Старший научный сотрудник Федерального государственного бюджетного учреждения науки "Казанский институт биохимии и биофизики" Казанского научного центра РАН
Давыдова Марина Николаевна
Ведущая организация: Институт молекулярной генетики РАН, г. Москва
Защита диссертации состоится «13» декабря 2012 г. в 13.00ч в на заседании диссертационного совета Д.212.081.08 при ФГАОУВПО «Казанский (Приволжский) федеральный университет» по адресу: 420008, г. Казань, ул. Кремлевская, 18, главное здание, ауд. 211.
Факс: 8(843)238-71-21, 233-78-40. E-mail: [email protected]
С диссертацией можно ознакомиться в научной библиотеке им. Н.И. Лобачевского Казанского (Приволжского) федерального университета
Автореферат разослан «___» октября 2012 г.

Ученый секретарь
диссертационного совета,
д. б. н, З. И. Абрамова
Актуальность проблемы. Секретируемые белки составляют до 50% от общего количества белковых компонентов клеток бацилл. Многие из них участвуют в гидролизе природных полипептидов и синтезируются в ответ на изменения в окружающей среде. Среди них особое внимание привлекают протеазы. Эти ферменты вовлечены в важнейшие физиологические процессы клетки, например, участвуют в катаболизме субстратов, споруляции, расщеплении и круговороте белков [Gupta R., et al., 2002]. Бактериальные протеиназы широко используются в пищевой промышленности, для производства моющих средств и имеют большой потенциал применения в качестве фармакологических препаратов, например, для лечения расстройств пищеварения или болезней сердечнососудистой системы [Gupta R., et al., 2002]. Как правило, в качестве продуцентов протеиназ используются различные штаммы бацилл. Однако, наличие комплекса внеклеточных протеаз у бацилл создает большую проблему в генной инженерии и биотехнологии, поскольку снижает накопление гетерологичных белков в культуральной жидкости продуцентов [Westers L., et al., 2004]. Решение этой проблемы требует создания новых штаммов-продуцентов и экспрессионных систем, основанных на индуцибельных бациллярных промоторах.
Одной из наиболее интересных групп протеолитических ферментов является класс сериновых протеаз. Они обнаружены у прионов, бактерий, вирусов, простейших и высших эукариот [Rawlings N.D., et al., 2012]. Сериновые протеазы эукариот участвуют во многих процессах, в том числе, в эмбриогенезе и развитии патологических состояний. В связи с этим, исследование прокариотических аналогов этих ферментов является удобной моделью при разработке новых лекарственных средств для борьбы с заболеваниями человека. Расшифровка механизмов регуляции генов прокариотических сериновых протеаз может стать основой для новой стратегии получения гетерологичных белков и создания высокоэффективных систем их экспрессии, что является актуальным направлением современной биотехнологии (генной инженерии).
Несмотря на объём знаний, накопленных о сериновых протеазах бацилл, функция этих ферментов в клетке и механизмы их регуляции изучены недостаточно. Для выяснения физиологической функции белка часто используют метод направленной инактивации гена [Biswas I., et al., 1993]. Штаммы с инактивированными генами протеиназ служат для получения гетерологичных белков, поскольку инактивация внеклеточных протеиназ позволяет достичь выхода продукта в количествах, необходимых для применения в биотехнологии. Для разработки эффективных рекомбинантных штаммов и новых систем экспрессии необходимы знания о механизмах регуляции генов. Анализ регуляторных областей, выяснение промоторной специфичности и расшифровка механизмов экспрессии генов является основой клонирования генов в составе экспрессионных систем.
Целью работы являлось создание новой экспрессионной системы на основе расшифровки механизмов экспрессии генов протеаз.
Основные задачи исследования:
- Установить роль протеаз в физиологии бацилл путём инактивации генов сериновых протеаз;
- Определить минимальную длину промоторов генов субтилизиноподобной протеазы и глутамилэндопептидазы;
- Определить влияние транскрипционных факторов (Spo0A, DegU, SigD, плейотропных репрессоров) на функционирование минимальных промоторов;
- Разработать систему экспрессии для получения гетерологичных белков на основе антибиотико-индуцибельного промотора двухкомпонентной системы LiaRS Bacillus subtilis;
- Оценить эффективность экспрессии репортёрного гена gfp и генов сериновых протеиназ в составе новой экспрессионной системы.
Научная новизна. Все результаты, изложенные в диссертационной работе, получены впервые.
- Разработаны новые беспротеазные штаммы B. pumilus с инактивированными генами субтилизиноподобной протеазы и глутамилэндопептидазы. Получены приоритетные данные, свидетельствующие, что инактивация генов протеаз привела к изменениям в клеточной морфологии, ускорению лизиса клеток, частичному изменению в способности разлагать углеводы и изменению уровня секреции других гидролаз.
- Впервые изучена зависимость экспрессии генов сериновых протеаз от длины промоторного региона генов на основе репортёрных fusion-конструкций (PaprBp-gfp, PgseBp-gfp). Установлены минимальные области регуляции генов, свидетельствующие, что длина промотора обусловлена вкладом фермента в физиологию бацилл.
- Разработана новая эффективная система экспрессии генов на основе liaI промотора (PliaI) B. subtilis, который регулируется двухкомпонентной антибиотико-индуцибельной системой LiaRS. При клонировании гена зелёного флуоресцентного белка (gfp) в данной системе показано, что в течение 30 мин после индукции экспрессия гена gfp возрастает в 100 раз, что свидетельствует об эффективности экспрессии рекомбинантных белков. Установлено, что делеция на хромосоме liaIH-оперона приводит к увеличению экспрессии генов в составе сконструированных плазмид.
Практическая значимость результатов. Беспротеазные штаммы B. pumilus, полученные в работе, могут служить в качестве реципиентов для получения гетерологичных белков, повышения уровня их продукции и качества целевого белка за счет снижения протеолитической деградации. Эти штаммы могут применяться в биотехнологии, генной инженерии и, в частности, быть использованы для получения минорных компонентов спектра внеклеточных протеаз B. pumilus.
Данные о структуре промоторов генов сериновых протеаз B. pumilus 3-19 свидетельствуют о наличии репрессорных и/или энхансерных элементов, что необходимо учитывать при клонировании генов.
Разработанная новая экспрессионная система позволяет получить высокий выход гетерологичных белков в бациллах, что чрезвычайно важно при получении промышленно важных микробных препаратов.
Положения, выносимые на защиту:
- Гены гидролаз бацилл имеют различную длину промоторов, которая определяется их функцией в физиологии бактерий.
- Инактивация генов протеолитических ферментов бацилл приводит к плейотропному эффекту: изменяется спектр внеклеточных гидролаз, частично морфология и биохимические характеристики.
3. Новая антибиотико-индуцибельная система экспрессии на основе промотора PliaI B. subtilis эффективна для продукции гетерологичных белков, включая протеазы бацилл.
Связь работы с научными программами. Работа выполнена в соответствии с планом НИР Казанского (Приволжского) федерального университета (№ гос. регистрации 01:02.00 104982 «Биосинтез, биогенез, классификация, физиологические функции новых микробных ферментов и возможные области их практического применения»). Исследования выполнены при поддержке грантов РФФИ № 09-04-99044-р_офи, Германской службы академических обменов DAAD 2009-2011 гг.: № A/08/75088, № A/09/84065, Федеральной целевой программы «Научные и научно-педагогические кадры инновационной России» 2009-2013 гг.: ГК № П344, ГК № П406, ГК № П323, ГК № 815, ГК № 1053.
Апробация работы. Основные положения диссертации доложены и обсуждены на IV Российском симпозиуме "Белки и пептиды" (Казань, РФ, 2009), XIV Международной конференции "Ферменты микроорганизмов в биологии и медицине" посвященной 20-летию партнерства между Казанским государственным университетом и Гиссенским университетом им. Ю. Либиха (Казань, РФ, 2009), Всероссийской школе-конференции молодых ученых «Актуальные проблемы современной биохимии и молекулярной биологии», (Казань, РФ, 2009), 3-й совместной конференции Немецкого общества гигиены и микробиологии (DGHM) и Ассоциации по общей и прикладной микробиологии (VAAM) (Ганновер, Германия, 2009), X Научной конференции молодых учёных, аспирантов и студентов научно-образовательного центра Казанского (Приволжского) федерального университета (Казань, РФ, 2011), III Ежегодной Международной научно-практической конференции по биологии студентов, магистрантов и аспирантов Тбилисского государственного университета им. Иванэ Джавахишвили (Тбилиси, Грузия, 2011), Итоговой научно-образовательной конференции студентов Казанского (Приволжского) федерального университета (Казань, РФ, 2011), XIX Международной конференции студентов, аспирантов и молодых учёных «ЛОМОНОСОВ-2012», МГУ, (Москва, РФ, 2012), XI Научная конференция молодых учёных, аспирантов и студентов научно-образовательного центра Казанского (Приволжского) федерального университета «Материалы и технологии XXI века» (Казань, РФ, 2012).
Публикации. По материалам диссертации опубликовано 28 работ, из них 4 статьи в центральных отечественных рецензируемых журналах.
Структура и объём диссертации. Диссертация состоит из введения, обзора литературы, описания материалов и методов исследований, раздела экспериментальных исследований, обсуждения результатов, выводов и списка литературы. Работа изложена на 132 страницах машинописного текста, включает 2 таблицы, 28 рисунков. Библиография содержит 178 наименований российских и зарубежных авторов.
СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы
Штаммы бактерий, плазмидные вектора и условия культивирования. В работе использовали штаммы из музея лаборатории Биосинтеза и Биоинженерии Ферментов КФУ: стрептомицинустойчивый штамм ![]() B. pumilus 3-19, протеазо-дефицитный штамм B. subtilis BG2036 (предоставлен проф. Eugenio Ferrarri, Genencor Int. Inc., USA), штамм B. subtilis W168 (trpC2). Использовали штаммы и плазмиды, любезно предоставленные научной группой Dr. Prof. T. Mascher (Мюнхенский университет Людвига-Максимилиана, г. Мюнхен, Германия): degU::kan (TMB124), B. subtilis W168 spo0A::tet (TMB205), B. subtilis W168 sigD::spec (TMB225), B. subtilis W168 amyE:: pSJ5101 (PliaI-gfp) (TMB408), B. subtilis WT168 PliaI-liaIH clean deletion (TMB604), E. coli DH5 (recA1 endA1 gyrA96 thi hsdR17(rK- mK+) relA1 supE44 80lacZM15 (lacZYA-argF)U169), вектора - pDG1662, pGP380, pGFPamyE, pSG1151, pMAD.
B. pumilus 3-19, протеазо-дефицитный штамм B. subtilis BG2036 (предоставлен проф. Eugenio Ferrarri, Genencor Int. Inc., USA), штамм B. subtilis W168 (trpC2). Использовали штаммы и плазмиды, любезно предоставленные научной группой Dr. Prof. T. Mascher (Мюнхенский университет Людвига-Максимилиана, г. Мюнхен, Германия): degU::kan (TMB124), B. subtilis W168 spo0A::tet (TMB205), B. subtilis W168 sigD::spec (TMB225), B. subtilis W168 amyE:: pSJ5101 (PliaI-gfp) (TMB408), B. subtilis WT168 PliaI-liaIH clean deletion (TMB604), E. coli DH5 (recA1 endA1 gyrA96 thi hsdR17(rK- mK+) relA1 supE44 80lacZM15 (lacZYA-argF)U169), вектора - pDG1662, pGP380, pGFPamyE, pSG1151, pMAD.
Культивирование бактерий проводили на следующих средах: среда LB (%): триптон – 1,0; дрожжевой экстракт – 0,5; NaCl – 0,5; pH 8.5 [Sambrook et al., 1989]. Агаризованная среда LB включала дополнительно 2% агара. Среды для трансформации штаммов B. subtilis включали солевую основу cреды Спицайзена, (Спицайзена I/II) [Anagnostopolous, Spizizen, 1961]. Идентификационная агаризированная среда для отбора клонов, способных секретировать протеиназу, включала 30% обезжиренного молока и 2% агара. Среды стерилизовали при 1 атм. в течение 30 мин., рН доводили перед стерилизацией среды 40%-ным раствором NaOH до значения 8.5. В среду при необходимости добавляли антибиотики в конечных концентрациях (мкг/мл): стрептомицин - 10, ампициллин - 20, эритромицин – 20, линкомицин – 20, хлорамфеникол - 5, канамицин - 10, тетрациклин - 10, спекциномицин - 100.
Получение рекомбинантных конструкций и работа с ДНК. Процедуры по получению рекомбинантной плазмидной/хромосомной ДНК проводили в клетках E. coli, как описано в [Sambrook et al., 1989]. Праймеры конструировали с помощью программного обеспечения Oligo Calc (http://www.basic.northwestern.edu/biotools/oligocalc.html) на основании последовательностей промоторов и генов: aprBp B. pumilus 3-19 (AN AY754946.2), gseBp B. pumilus 3-19 (AN Y15136.1), liaIHGFSR-оперона (AN AL009126.3) B. subtilis 168. Синтез праймеров и секвенирование проводила фирма «Синтол» (Москва). Для клонирования использовали ферменты фирмы «Сибэнзим» (Москва). Выделение геномной ДНК, рекомбинантной плазмидной ДНК, очистку продуктов ПЦР реакции проводили с помощью наборов реактивов фирмы Fermentas (Латвия). При конструировании репортёрных fusion-конструкций (PaprBp/PgseBp+gfpmut3) различные фрагменты ДНК (промоторы генов aprBp, gseBp B. pumilus 3-19) клонировали в вектор pGFPamyE как описано в работе [Bisicchia P. et al., 2010]. Для изучения влияния транскрипционных факторов на экспрессию генов сериновых протеиназ B. pumilus, генные мутации по соответствующим факторам транскрипции совмещали с полученными репортёрными fusion-конструкциями. Трансформацию клеток B. subtilis плазмидной ДНК проводили, как описано в работе [Anagnostopolous et al., 1961].
Конструирование новой LIKE-системы экспрессии и определение активности рекомбинантного промотора PliaI. Для конструирования интегративного и репликативного векторов новой экспрессионной системы в промоторе liaIH-оперона B. subtilis оптимизировали область рибосом-связывающего сайта. Такой оптимизированный промотор, полученный с помощью ПЦР реакции, клонировали в векторы pDG1662, pGP380 с образованием плазмид pLia-int и pLia-rep соответственно. Амплификаты генов gfpmut1, aprBp, gseBp клонировали в полученные экспрессионные векторы. Промотор PliaI индуцировали добавлением в среду антибиотика бацитрацина (30 мкг/мл). Активность промотора (A=dGFP/dt/OD600) определяли для каждого времени измерения как описано в работе [Botella E., et al., 2010]. При определении активности рекомбинантного GFP белка, сумма естественной флуоресценции штаммов дикого типа (B. subtilis 168) вычиталась из суммы флуоресценции штаммов с репортёрными конструкциями (PliaI+gfpmut1; PaprBp/PgseBp+gfpmut3).
Определение протеолитической активности. Общую протеолитическую активность рекомбинантных штаммов определяли по расщеплению азоказеина (Sigma, США) [Charney J. et al., 1947]. Специфическую активность субтилизиноподобной протеазы определяли по расщеплению хромогенного субстрата Z-Ala-Ala-Leu-pNa по методу [Люблинская Л.А., с соав., 1987]. Специфическую активность глутамилэндопептидазы определяли по расщеплению хромогенного субстрата Z-Glu-pNa по методу [Houmard J., 1967]. Активность внеклеточной щелочной рибонуклеазы определяли по кислоторастворимым продуктам гидролиза РНК [Лещинская И. Б., с соавт., 1980]. Фосфомоноэстеразную активность определяли по действию культуральной жидкости (КЖ) на p-нитрофенилфосфат («Serva», Германия) [Лещинская И. Б., с соавт., 1980].
Секвенирование и геноинформатика. Анализ нуклеотидных последовательностей проводили с помощью программного пакета Clone Manager 6 version 6.00 (SciEdCentral). Выравнивание последовательностей проводили с помощью on-line программного обеспечения NCBI/BLAST.
Математическая обработка результатов. Для статистического анализа экспериментальных данных использовали программу Microsoft Excel, путём расчёта среднеквадратичного отклонения (). Результаты считали достоверными при 10%. При расчёте достоверности получаемых разностей использовали критерий Стьюдента, принимая P 0.05 за достоверный уровень значимости.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ
I. in silico характеристика промоторов генов сериновых протеаз B. pumilus. Поскольку структура и длина регуляторных цис-элементов генов протеаз B. pumilus может значительно различаться, промоторные регионы генов aprBp и gseBp проанализировали с использованием on-line программы NCBI/BLAST. Выравнивание промоторных областей размером 445 п.о. для гена aprBp и 1149 п.о. для гена gseBp, соответствующих ранее определенным межгенным регионам, показало высокую степень гомологии с нуклеотидными последовательностями штамма B. pumilus SAFR-032. Небольшая область (58 п.о.) в промоторе гена aprBp (-445…+1) имела слабую гомологию с последовательностью гена фермента фосфоглицератмутазы (yhfR, AN CP000813.1). Напротив, в регуляторном регионе гена gseBp (-1149…+1) обнаружили протяжённую последовательность (905 п.о.) с высокой степенью гомологии к гену NADPH-редуктазы (yrhJ, AN CP000813.1, 91% гомологии). Полученные результаты on-line выравнивания позволили предположить наличие открытой рамки считывания (ОРС) внутри 5'-межгенного региона гена gseBp (в положении -1094…-189 относительно сайта инициации транскрипции гена) (рис. 1).

Рис. 1. Создание репортёрных конструкций для определения минимальных регуляторных участков генов aprBp (а) и gseBp (б). Изогнутыми линиями обозначена область начала транскрипции генов. Фрагмент длиной 905 п.о. с 91% гомологией к гену NADPH-редуктазы штамма B. pumilus SAFR-032 найденный в 5'-межгенном регионе гена gseBp обозначен в виде серого пятиугольника. Линией обозначены участки 5'-межгенных регионов (потенциальных промоторов), использованные для создания fusion-конструкций.
Определение минимального участка промотора, необходимого для полноценной экспрессии генов сериновых протеаз. Для определения минимального регуляторного участка (промотора) генов aprBp и gseBp сконструировали репортёрные fusion-конструкции на основе гена зелёного флуоресцентного белка gfp и промоторных участков генов протеаз B. pumilus 3-19. Три варианта каждого промотора (включающие -445 п.о., -310 п.о., -280 п.о. для гена aprBp; -150 п.о., -122 п.о., -100 п.о. – для гена gseBp) были транскрипционно соединены с маркёрным геном gfpmut3 (рис. 1). Динамика роста и накопление флуоресценции полученных конструкций представлены на рисунке 2. Флуоресцентный сигнал появлялся во всех клетках в переходной фазе развития культуры (на 5-8 часы роста). Анализ PaprBp-gfpmut3 fusion-конструкций показал, что уменьшение 5'-регуляторной области гена субтилизиноподобной протеиназы AprBp (с -445 п.о. до -310 п.о.) приводит к значительному подавлению экспрессии репортёрного гена gfpmut3 (рис. 2а 1/2). Укорочение промоторного региона до -280 п.о. относительно сайта инициации транскрипции гена aprBp приводило к практически полному ингибированию активности белка gfpmut3.
(а1) (б1)


(а2) (б2)


Рис. 2. Динамика роста, накопление флуоресценции (а1, б1) и экспрессия гена gfpmut3 (а2, б2) в штаммах, содержащих fusion-конструкции PaprBp-gfpmut3 (а 1/2), PgseBp-gfpmut3 (б 1/2). Обозначения конструкций: промотор субтилизиноподобной протеазы r-PaprBp445, Ї-PaprBp310, Ј-PaprBp280; промотор глутамилэндопептидазы r-PgseBp150, Ї-PgseBp122, Ј-PgseBp100. Рост штаммов обозначен без символов.
Изучение fusion-конструкций с промотором гена глутамилэндопептидазы показало, что при длине регуляторной области (PgseBp) менее 150 п.о. экспрессия репортёрного гена резко снижается (рис. 2б 1/2).
Таким образом, были определены минимальные регуляторные фрагменты генов протеаз B. pumilus: 150 п.о. для гена глутамилэндопептидазы и 445 п.о. для гена субтилизиноподобной протеазы.
Экспрессия fusion-конструкций с минимальной областью промоторов протеаз в регуляторных мутантах. При лимитации источников питания в клетках почвенных бактерий формируется адаптивный ответ, направленный на выживание. В частности, клетки начинают активно синтезировать различные гидролитические белки (протеазы) [Lpez D., et al., 2010]. Такой процесс обусловлен синтезом в клетке определённых транскрипционных факторов (регуляторов), которые взаимодействуют с отдельными промоторами генов протеаз и активируют их экспрессию. Поэтому, мы исследовали репортёрные fusion-конструкции PaprBp-gfpmu3 и PgseBp-gfpmu3, содержащие минимально-необходимую область промоторов генов aprBp и gseBp в штаммах дефектных по регуляторным белкам. Показано, что белки DegU и Spo0A оказывают позитивный эффект на активность генов обеих протеаз, т.к. удаление этих регуляторов подавляет экспрессию в репортёрных конструкциях: делеция гена degU – полностью, делеция гена spo0A – частично (рис. 3, 4).
Полученные результаты согласуются с данными литературы о позитивном влиянии этих регуляторов на продукцию сериновых протеиназ. Более того, промоторы данных генов содержат потенциальные сайты взаимодействия с исследуемыми (DegU и Spo0A) транскрипционными факторами [Sharipova M.R., et al., 2007, 2008].
(а) (б)


Рис. 3. Экспрессия P445aprBp-gfpmut3 (а), P150gseBp-gfpmut3 (б) fusion-конструкций в рекомбинантном штамме с дефектом гена регуляторного белка DegU (degU::kan) - Ј; штамм без мутаций - r.
(а) (б)


Рис. 4. Экспрессия P445aprBp-gfpmut3 (а), P150gseBp-gfpmut3 (б) fusion-конструкций в рекомбинантном штамме с дефектом гена регуляторного белка Spo0A (spo0A::tet) - ™; штамм без мутаций - r.
Продукция гидролитических ферментов, в свою очередь, может конкурировать с другими адаптационными механизмами клетки (например, клеточной подвижностью). Чтобы проверить это предположение, изучали экспрессию репортёрных fusion-конструкций PaprBp-gfpmu3, PgseBp-gfpmu3 в мутантном штамме с делецией по регуляторному белку альтернативного сигма фактора - D. Функционирование sigD регулятора обеспечивает в клетках бацилл сборку жгутикового скелета и синтез автолизинов. В результате клетки способны к движению и не связаны между собой [West J.T., et al., 2000]. Данные, представленные на рисунке 5 доказывают, что в мутантном штамме (sigD::spec) сильная экспрессия репортёрного гена под контролем промоторов обеих сериновых протеаз B. pumilus начинается в начале переходной фазы.
(а) (б)


Рис. 5. Экспрессия P445aprBp-gfpmut3 (а), P150gseBp-gfpmut3 (б) fusion-конструкций в рекомбинантном штамме с дефектом гена регуляторного белка SigD - Ї; штамм без мутаций - r.
По-видимому, инактивация гена альтернативного сигма фактора (sigD), отвечающего за формирование клеточной подвижности, приводит к активной продукции клетками бацилл протеолитических ферментов.
Таким образом, характеристика промоторов генов сериновых протеиназ B. pumilus показала, что обе протеазы имеют сложный механизм регуляции. Промоторы доминирующей и минорной протеаз различны по длине и организации: промотор гена мажорной протеазы AprBp более протяженный (445 п.о.), вероятно, имеет большое количество консенсусных ДНК сайтов связывания регуляторных белков по сравнению с промотором (150 п.о.) гена минорной протеазы GseBp. Такая структура промоторов соответствует биохимическим характеристикам и функциям данных ферментов в клетках B. pumilus: доминирующая в протеолитическом пуле протеаза (70% активности) неспецифически расщепляет пептиды, белковые субстраты, минорная глутамилэндопептидаза (10% активности) гидролизует гидрофильные пептиды. Более того, активность данных ферментов разобщена по времени действия для оптимального поглощения питательных веществ из среды [Sharipova M.R., et al., 2007, 2008].
II. Получение и характеристика штаммов с нокаутированными генами сериновых протеаз. Для получения штаммов с инактивированными генами использовали подход, основанный на гомологичной рекомбинации между участками экзогенной и геномной ДНК, приводящей к нарушению ОРС генов aprBp и gseBp B. pumilus. Были созданы рекомбинирующие конструкции, в которых ген эритромицина (erm) встроен в гены сериновых протеаз. После трансформации полученными конструкциями клеток B. pumilus 3-19 проводили отбор мутантов на чашках с эритромицином. Колонии, выросшие на чашках с антибиотиком, тестировали на молочном агаре по уменьшению зон гидролиза казеина вокруг колоний. Выраженные зоны просветления наблюдали только у исходного штамма B. pumilus 3-19. У всех модифицированных штаммов зоны протеолиза отсутствовали, что указывало на инактивацию генов секретируемых протеолитических ферментов. Наличие остаточных зон гидролиза может объясняться активностью оставшихся секретируемых ферментов протеолитического спектра. В результате отобрали рекомбинантные штаммы B. pumilus MK10 (aprBp::erm) и B. pumilus 2A-5 (gseBp::erm), которые при тестировании на молочном агаре показали отсутствие протеолитической активности.
Морфология рекомбинантных штаммов (MK10 и 2A-5) по сравнению с диким штаммом (3-19) отличалась: длина клеток MK10 и 2A-5 составляла около 14 мкм (против 8 мкм для дикого штамма); поверхность колоний при росте на агаризованной среде была шероховатая, колонии имели неровные края (по сравнению с более гладкими, ровными краями и слизистыми колониями исходного штамма). На стандартной питательной среде LB показано, что экспоненциальный рост и накопление биомассы для всех трёх штаммов одинаковы. Штаммы одновременно переходили в стационарную фазу, однако склонность к лизису особенно сильно наблюдали у штамма B. pumilus 2A-5 (gseBp::erm) (рис. 6а). У обоих мутантных штаммов (по сравнению с исходным) нарушено спорообразование: появление свободных спор происходит к 36 часу, а на 45 час их количество составляет ~5% (рис. 6а).
При повышении температуры культивирования до 42°C в обоих штаммах сокращалась продолжительность стационарной фазы и происходил почти одновременный переход к лизису культуры (рис. 6б). Полученные данные свидетельствуют, что инактивация генов внеклеточных субтилизиноподобной протеиназы и глутамилэнодопептидазы оказывает эффект на спорообразование и продолжительность стационарной фазы роста. Усиление лизиса клеток у мутантных штаммов подтверждает участие сериновых протеинз в гидролизе автолизинов.
(а) (б)


Рис. 6. Динамика роста и спорообразования исследуемых штаммов, выращенных в жидкой среде LB. (а) Рост/спорообразование клеток: B. pumilus 3-19 (1/4); нокаут по гену aprBp::erm MK-10 (2/5), нокаут по гену gseBp::erm 2A-5 (3/6). (б) Рост клеток при температурах 42°C/15°C: B. pumilus 3-19 (1/4); нокаут по гену aprBp::erm MK-10 (2/5), нокаут по гену gseBp::erm 2A-5 (3/6).
Характеристика секретируемых гидролаз нокаутированных штаммов. Известно, что вклад протеаз, глутамилэндопептидазы и субтилизиноподобной протеазы, в общую протеолитическую активность штамма B. pumilus 3-19 различен – на AprBp протеазу приходится до 70%, на глутамилэндопептидазу GseBp до 10% [Sharipova M.R., et al., 2007, 2008]. Мы наблюдали эффект снижения активности различных протеаз в клетках, содержащих отдельные мутации по генам сериновых протеиназ aprBp и gseBp. По уровню общей протеолитической активности на азоказеине модифицированные штаммы МК10 и 2A-5 уступали исходному штамму 3-19 в ~2,5 раза (рис. 7а). У инактивированных штаммов активность на соответствующих нокауту субстратах отсутствовала. При этом, активность в отношении субстрата Z-Ala-Ala-Leu-pNА практически отсутствовала у обоих мутантных штаммов (рис. 7б), тогда как в отношении субстрата Z-Glu-pNА активность не выявлялась у мутанта 2A-5 (gseBp-), но определялась, хотя и на низком уровне у мутанта MK-10 (aprBp-) (рис. 7в). По-видимому, функционально-активная субтилизиноподобная протеиназа играет важную роль в формировании минорного фермента глутамилэндопептидазы. Глутамилэндопептидаза, в свою очередь, влияет на формирование пула внеклеточной субтилизиноподобной протеиназы.
Изучение активности РНКазы в штаммах с делециями протеаз показало, что инактивация протеазных генов не приводила к изменению её активности (рис. 7г), т.е. созревание фермента не зависит от присутствия субтилизиноподобной протеазы и глутамилэндопептидазы. В тоже время инактивация генов обеих сериновых протеиназ приводила к изменению уровня активности внеклеточной фосфатазы. По сравнению с исходным (3-19) штаммом активность фосфатазы в штамме 2А-5 повышалась в 3-6 раз на разных стадиях культивирования клеток (рис. 7д). Штамм с нокаутированным геном aprBp (МК-10) показал менее выраженное увеличение уровня фосфатазы, однако активность фермента была выше в ~2 раза в стационарной фазе по сравнению с исходным немодифицированным штаммом. Можно сделать заключение, что глутамилэндопептидаза участвует, по-видимому, в созревании фермента фосфогидролазы. В целом, полученные данные указывают на сбалансированность содержания протеаз в общем пуле внеклеточных ферментов и их корегуляцию.
(а) (б)

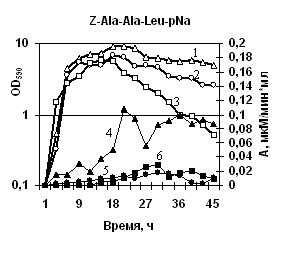
(в) (г)


(д)  | Рис. 7. Динамика роста и накопление активности ферментов в исследуемых беспротеазных штаммах и исходном штамме на различных субстратах. Рост клеток: B. pumilus 3-19 (1); нокаут по гену aprBp::erm MK-10 (2), нокаут по гену gseBp::erm 2A-5 (3). Активность ферментов: B. pumilus 3-19 (4); нокаут по гену aprBp::erm MK-10 (5), нокаут по гену gseBp::erm 2A-5 (6). |
Таким образом, полученные штаммы B. pumilus MK10 (aprBp::erm) и B. pumilus 2A-5 (gseBp::erm) могут быть рекомендованы для биотехнологических разработок в качестве штаммов-реципиентов для получения гетерологичных белков.
III. Разработка новой экспрессионной системы для получения гетерологичных белков в клетках B. subtilis. Известно, что присутствие в среде гликопептидных антибиотиков (например, бацитрацина, низина, ванкомицина и др.) нарушающих целостность клеточной мембраны, запускает в клетках бацилл работу четырёх сигнальных систем – альтернативного сигма фактора M, и трёх двухкомпонентных систем (LiaRS, BceRS, YvcPQ) [Jordan S., et al., 2008]. Гены liaIH-оперона имеют наибольший уровень экспрессии. В отсутствие индуктора, например в логарифмической фазе роста, промотор этого оперона (PliaI) является «молчащим» (не проявляет базальный уровень экспрессии). При его индукции определённым стимулом (антибиотиками, детергентами, органическими растворителями и др. веществами) может достигаться 100- или 1000-кратное увеличение уровня экспрессии генов в течение 5-10 мин. Строгая регуляция данного промотора определяется функционированием ответного регулятора LiaR. На основе характеристик промотора liaIH-оперона нами сконструирована новая экспрессионная система (LIKE-система, от нем. LIa-Kontrollierte Expression) для получения рекомбинантных белков в клетках B. subtilis. Получены два типа плазмид, несущих индуцируемый промотор PliaI (рис. 8).




Рис. 8. Схема плазмид pLIKE-int и pLIKE-rep, несущих индуцируемый PliaI промотор B. subtilis.
Вектор pLIKE-int является интегративным, т.е. при трансформации в клетки B. subtilis встраивается в геном. Таким образом, рекомбинантная конструкция (PliaI промотор + ген интересуемого белка) переносится в хромосому по участкам гена амилазы (amyE-back и amyE-front). Вектор pLIKE-rep, реплицирующийся в клетках как плазмида (шатл E. coli/B. subtilis), позволяет получить несколько копий рекомбинантной ДНК за счёт наличия сильной ori-последовательности – CoLE1. Длина регуляторного региона в составе обоих векторов составляет 101 п.о. и включает LiaR-связывающую последовательность для индукции промотора PliaI.
Для увеличения продукции интересуемых белков провели оптимизацию рибосом-связывающей области промотора PliaI до канонической последовательности (–taaggagg–), характерной для генов бациллярных белков (рис. 9).

Рис. 9. Часть последовательности промотора PliaI, входящего в состав разработанных экспрессионных векторов. PliaI содержит оптимизированный канонический сайт Шайна-Дальгарно (выделен жирным шрифтом) и полилинкер. В рамку выделена область из 7 нуклеотидов, расположенных до стартового кодона ATG.
Получение рекомбинантных штаммов B. subtilis для усиления активности PliaI промотора. На основе структурной организации liaIH-liaGFSR регулона и его функционирования предположили, что активация PliaI промотора приводит к усилению экспрессии генов liaGFSR локуса, за счёт «проскакивания» РНК-полимеразы через слабый терминатор [Mascher T., et al., 2004]. Тогда удаление терминатора будет способствовать усилению активности индуцируемого PliaI промотора и позволит добиться повышенной продукции рекомбинантных белков. С другой стороны показано, что гиперпродукция LiaH белка в клетках приводит к снижению уровня трансляции (сокращение аминокислот и рибосом), что нежелательно для производства рекомбинантных белков. Основываясь на данных знаниях, мы создали коллекцию реципиентных рекомбинантных штаммов B. subtilis, предполагая, что дополнительные регуляторные области на хромосоме (промотор, ген и терминатор) будут влиять на экспрессию рекомбинантного гена в составе LIKE системы экспрессии и, в конечном счёте, на выход целевого продукта. Схема liaIH-liaGFSR регулона полученных штаммов представлена на рисунке 10.

Рис. 10. Схема liaIH-liaGFSR регулона и генотип реципиентных рекомбинантных штаммов B. subtilis. Гены регулона обозначены сплошными стрелками. Промоторы представлены изогнутыми стрелками. Терминаторы представлены в виде шпилечной структуры.
Характеристика новой LIKE-системы на основе экспрессии генов рекомбинантных белков. В качестве индуктора LIKE-системы экспрессии нами выбран антибиотик бацитрацин. Бацитрацин оказывает наибольший индуцирующий эффект на PliaI промотор в клетках B. subtilis, в тоже время, максимальный уровень активности промотора достигается при низких концентрациях этого антибиотика [Rietktter E., et al., 2008]. В качестве репортёрного гена для изучения экспрессии в составе новой LIKE-системы выбрали ген gfpmut1. При сравнении активности PliaI промотора в составе pLIKE-int и pLIKE-rep плазмид во всех исследуемых рекомбинантных штаммах наблюдали быстрое (в течение 30 мин) накопление флуоресценции после добавлении бацитрацина (30 мкг/мл) в среду культивирования (рис. 11 A/B). Максимальный уровень активности PliaI промотора наблюдали в штаммах содержащих репликативный pLIKE-rep вектор по сравнению со штаммами несущими в клетках одну хромосомную копию рекомбинантной конструкции (т.е. в составе pLIKE-int вектора) (рис. 11 A/B).
Результат оптимизации рибосом-связывающего сайта наблюдали при сравнении продукции белка GFP в двух штаммах - W168 и TMB408, содержащих интегративную конструкцию - pLIKE-int+gfp (рис. 11 A/B). Показано, что оптимизация сайта Шайна-Дальгарно приводит к увеличению экспрессии рекомбинантного гена в ~6 раз.
Делеция нативного PliaI промотора на хромосоме штамма TMB604 приводила к снижению активности PliaI(opt) промотора в составе экспрессионного интегративного вектора в 1,5 раза, а в составе репликативного – 3 раза по сравнению со штаммом, имеющим генотип дикого типа (рис. 11 B). Делеция элементов liaIH-оперона, но сохранение нативного PliaI промотора на хромосоме (TMB1151/1152), почти не приводила к изменению активности рекомбинантного PliaI(opt) промотора при использовании pLIKE-int плазмиды. Однако, при использовании репликативного pLIKE-rep вектора, активность белка GFP в штаммах TMB1151/1152 увеличивалась в 1,2 раза (рис. 11 B).

Рис. 11. Рост, показания флуоресценции (А) и активность промотора (B) в штаммах несущих трансляционные конструкции PliaI-gfpmut1 и PliaI(opt)-gfpmut1. Кривые роста показаны без символов. Обозначения экспрессии в разных штамах: () (W168), () (TMB604), () (TMB1151), () (TMB1152) () (TMB408). Вертикальной пунктирной линией показано время добавление индуктора (OD600~0.4-0.4, бацитрацин в концентрации 30 мкг/мл).
(С) Western blot анализ цитоплазматической фракции клеток, содержащих полученные конструкции с использованием антител к белкам LiaH и GFP. Обозначения: 1,2 - экспрессия конструкций с нативного промотора PliaI, 3-10 – экспрессия конструкций PliaI(opt) в отсутствии (-) и присутствии (+) бацитрацина (концентрация антибиотика 30 мкг/мл).
Таким образом, результаты экспериментов показали положительный эффект оптимизации последовательности Шайна-Дальгарно в промоторе PliaI новой LIKE-системы для экспрессии рекомбиантных белков. Кроме того, наблюдали небольшой положительный эффект делетирования терминатора liaIH-оперона. Для подтверждения достоверности повышения выхода рекомбинантного белка GFP, провели Western-blot анализ с использованием антител к данному белку, а также к белку LiaH. (рис. 11 C). Показали, что оба белка не обнаруживаются в клетках до индукции бацитрацином, что свидетельствует о строгом контроле активности PliaI промотора в клетках B. subtilis. После добавления антибиотика в среду белки GFP и LiaH накапливались в клетках в различной концентрации, что определялось генотипом полученных конструкций (рис. 10, рис. 11).
Таким образом, нами разработан эффективный инструментарий – LIKE-система экспрессии B. subtilis, основанная на стресс индуцибельном промоторе liaIH-оперона и ряд штаммов с делециями liaIH-liaGFSR регулона на хромосоме. Разработанная система позволяет направленно экспрессировать интересуемые гены как в составе плазмидного элемента – репликативная конструкция (pLIKE-rep), так и в составе генома клеток – интегративная конструкция (pLIKE-int). Показано, что максимальный уровень экспрессии PliaI промотора в составе pLIKE-int вектора происходит через 20-30 мин после добавления антибиотика, в то время как в составе вектора pLIKE-rep – через 60 мин, при этом его уровень в 10 раз выше, что объясняется мультикопийностью репликативной плазмиды.
LIKE-система для продукции генов сериновых протеаз. Для тестирования новой системы экспрессии бацилл нами использованы гены сериновых протеаз B. pumilis 3-19. Известен потенциал этих ферментов в биотехнологии и медицине [Ицкович Е.Л., с совт., 1998], что определяет необходимость получения чистого продукта в большом количестве. Гены глутамилэндопептидазы и субтилизиноподобной протеиназы клонировали в репликативный вектор pLIKE-rep под контроль PliaI(opt) промотора. Полученными плазмидами трансформировали беспротеазный штамм B. subtilis BG2036 и исследовали активность протеаз на молочном агаре и с применением специфических субстратов. Как видно из рисунка 12 (а), после 16 ч инкубирования рекобинантных штаммов на молочном агаре с добавлением бацитрацина, клетки эффективно продуцируют протеазы в среду.


(б) (с)


Рис. 12. (а) Колонии исходного штамма B. pumilus 3-19 и рекомбинантных штаммов B. subtilis, содержащих конструкции PliaI-aprBp и PliaI-gseBp на молочном агаре с добавлением антибиотика бацитрацина (30 мкг/мл) после 16 ч инкубирования.
(б) Активность субтилизиноподобной протеиназы исходного B. pumilus 3-19 и рекомбинантного B. subtilis (PliaI-aprBp) штаммов.
(с) Активность глутамилэндопептидазы исходного B. pumilus 3-19 и рекомбинантного B. subtilis (PliaI-gseBp) штаммов.
Исходный штамм показан в белом цвете. Рекомбинантные штаммы при добавлении бацитрацина показаны в чёрном цвете, без добавления – в сером цвете. Стрелочкой указано время добавления индуктора.
Анализ рекомбинантных штаммов, содержащих гены сериновых протеаз под контролем индуцируемого PliaI(opt) промотора на специфических субстратах показал, что активность ферментов повышалась в 2 раза. Полученный результат по активности ферментов можно объяснить тем, что для продукции протеолитических ферментов требуется формирование Sec-каналов и белков для эффективной секреции протеаз в среду. На это указывают результаты культивирования штаммов на молочном агаре. Вероятно, для дальнейшего повышения уровня белка в КЖ необходима оптимизация системы. Таким образом, новая LIKE-система экспрессии бацилл эффективна для производства секреторных рекомбинантных белков.
Итак, основные характеристики новой разработанной LIKE экспрессионной системы - (1) отсутствие экспрессии PliaI промотора если в среду не добавлен индуктор, (2) быстрая и высокая (100-1000-кратная) экспрессия гена под контролем PliaI промотора, (3) возможность использования различных индукторов, (4) позитивное влияние разработанных рекомбинантных штаммов на экспрессию PliaI промотора, (5) возможность применения системы для продукции ферментов используемых в медицине и пищевой индустрии – предполагают её использование, как альтернативной и конкурентоспособной экспрессионной системы B. subtilis.
ВЫВОДЫ
- Установлена минимальная область регуляции генов протеаз штамма Bacillus pumilus: для гена субтилизиноподобной протеиназы промотор составляет 445 п.о.; для гена глутамилэндопептидазы - 150 п.о.
- Установлено, что для репортёрных конструкций с минимальной областью регуляции протеаз факторы транскрипции DegU и Spo0A оказывают позитивное влияние, тогда как альтернативный сигма фактор транскрипции SigD – негативное.
- Инактивация генов протеолитических ферментов приводит к плейотропному эффекту: функционально-активные протеиназы участвуют в формировании пула гидролитических ферментов.
- Сконструирована новая LIKE-система экспрессии на основе антибиотико-индуцибельного промотора LiaRS двухкомпонентной системы Bacillus subtilis.
- В составе новой экспрессионной системы экспрессия репортёрного гена gfp повышалась в среднем в 6 раз, а генов сериновых протеиназ в 2 раза.
Публикации по теме диссертации в изданиях, рекомендованных ВАК:
- Тойменцева А.А. Bacillus intermedius с нокаутированным геном субтилизиноподобной протеиназы / А.А. Тойменцева, М.Р. Каримова, А.А. Ризванов, Е.И. Шагимарданова, М.Р. Шарапова // Учен. Зап. Казан. ун-та. Сер. Естеств. науки. -2010. -Т. 152, кн. 4. -С. 143-155. (перечень ВАК), автора – 0,5 п.л.
- Тойменцева А.А. Влияние делений в промоторе на экспрессию гена глутамилэндопептидазы Bacillus intermedins / А.А. Тойменцева, Е.И. Шагимарданова, А.Р. Каюмов, М.Р. Шарипова // Учен. зап. Казан. ун-та. Сер. Естеств. науки. -2010.-Т. 152. кн. 3. -С. 149-158. (перечень ВАК), автора – 0,5 п.л.
- Черёмин A.M. Выделение регуляторных белков, активирующихся в условиях ограниченного роста бацилл / A.M. Черёмин, А.А. Тойменцева, А.Р. Сабирова, А.Р. Каюмов, А.Б. Маргулис, М.Р. Шарипова // Учен. зап. Казан. ун-та. Сер. Естеств. науки. -2010. -Т. 152. кн. 4. -С. 156-168. (перечень ВАК), автора – 0,3 п.л.
- Шарипова М.Р. Новое филогенетическое положение штамма Bacillus intermedius 3-19 / М.Р. Шарипова, А.А. Тойменцева, А.Р. Сабирова, А.Д. Мухаметзянова, А.И. Ахметова, А.М. Марданова, Н.П. Балабан // Микробиология. -2011. -Т. 80. -№ 3. -С. 424-426. (перечень ВАК) автора – 0,05 п.л.
Другие публикации по теме диссертации:
- Тойменцева А.А. Экспрессия гена глутамилэндопептидазы Bacillus intermedius модифицированного в области промотора в клетках Bacillus subtilis / А.А. Тойменцева, М.Р. Каримова, Е.И. Шагиморданова, М.Р. Шарипова // Материалы докладов XIV международной научной конференции студентов, аспирантов и молодых учёных «Ломоносов» Международный молодёжный научный форум «Ломоносов -2007». -Москва, 2007. –С.45.
- Тойменцева А.А. Получение гена глутамилэндопептидазы Bacillus intermedius с модифицированной областью регуляции и изучение его экспрессии в клетках Bacillus subtilis / Е.И. Шагиморданова, А.А. Тойменцева, М.Р. Каримова, М.Р. Шарипова // Тезисы докладов и стендовых сообщений VI симпозиума «Химия портеолитических ферментов». –Москва, 2007.
- Toymentseva A.A. Specificity of an expression of the modified genes Bacillus intermedius in Bacillus subtilis cells / A.A. Toymentseva, M.R. Karimova, M.R. Sharipova // International scientific and technical conference “SCIENCE AND EDUCATION-2008”. –Murmansk, 2008. -P.585-586.
- Тойменцева А.А. Экспрессия гена глутамилэндопептидазы Bacillus intermedius, модифицированного в области промотора в клетках Bacillus subtilis / А.А. Тойменцева, М.Р. Каримова, М.Р. Шарипова // XV Международная конференция студентов, аспирантов и молодых учёных «ЛОМОНОСОВ-2008». – Москва, 2008.
- Toymentseva A.A. Construction genes of glutamyl endopeptidase Bacillus intermedius with deletion in the regulation of area and analysis their expression in Bacillus subtilis cells / M.R. Karimova, A.A. Toymentseva // Materials of XLVI international scientific conference of students “Scientific-and-technological advance”. –Novosibirsk, 2008. -P.38-39.
- Toymentseva A.A. Identification promoter area of gseBi required for potential expression of this enzyme / A.I. Akhmetova, A.A. Toymentseva, A.R. Kayumov, M.R. Sharipova // Materials of IV Russian symposium called “Protein and Peptides”. -Kazan, 2009. -P.329.
- Toymentseva A.A. Characteristic of functional regulation activity and proteases’ genes expression in stress conditions of bacillus / M.R. Sharipova, A.A. Toymentseva, A.R. Kayumov, A.R. Sabirova, N.L. Rudakova, N.P. Balaban // Materials of IV Russian symposium called “Protein and Peptides”. –Kazan, 2009. -P.96.
- Sharipova M.R. Characteristic of functional regulation activity and proteases’ genes expression in stress conditions of bacillus / M.R. Sharipova, A.A. Toymentseva, A.R. Kayumov, A.R. Sabirova, N.L. Rudakova, N.P. Balaban // Materials of IV Russian symposium called “Protein and Peptides”. –Kazan, 2009. -P.96.
- Toymentseva A.A. Genetic modification of Bacillus intermedius glutamyl endopeptidase gene promoter / A.I. Akhmetova, A.A. Toymentseva, A.R. Kayumov, M.R. Sharipova // XIV international conference “microbial enzymes in biotechnology and medicine” devoted to the 20th anniversary of partnership between Kazan State University and Justus-Liebig Giessen University. –Kazan, 2009. -P.18.
- Sharipova M.R. The role of microbial proteinases in stress response of Gram-positive bacteria: from signaling to regulatory networks / M.R. Sharipova, A.A. Toymentseva, A.R. Kayumov, A.A. Rizvanov, E.I. Shagimardanova // XIV international conference “Microbial enzymes in biotechnology and medicine” devoted to the 20th anniversary of partnership between Kazan State University and Justus-Liebig Giessen University. -Kazan, 2009. -P.58.
- Toymentseva A.A. Inactivation of serine proteinase genes from Bacillus intermedius / A.A. Toymentseva, A.A. Akhmetova, M.R. Karimova, A.R. Kayumov, A.A. Rizvanov, M.R. Sharipova // XIV international conference “Microbial enzymes in biotechnology and medicine” devoted to the 20th anniversary of partnership between Kazan State University and Justus-Liebig Giessen University. -Kazan, 2009. -P.65.
- Cheryomin A.M. The purification of Bacillus subtilis DegU and Spo0A regulatory proteins from the Escherichia coli recombinant strains / A.M. Cheryomin, A.R. Kayumov, A.A. Toymentseva, N. Pina, M.R. Sharipova // XIV international conference “Microbial enzymes in biotechnology and medicine” devoted to the 20th anniversary of partnership between Kazan State University and Justus-Liebig Giessen University. -Kazan, 2009. -P.12.
- Cheryomin A.M. The purification of Bacillus subtilis DegU and Spo0A regulatory proteins from the Escherichia coli recombinant strains / A.M. Cheryomin, A.R. Kayumov, A.A. Toymentseva, N. Pina, M.R. Sharipova // 14th international conference “Microbial enzymes in biotechnology and medicine”. -Kazan, 2009. -P.72.
- Cheryomin A.M. The purification of Bacillus subtilis DegU and Spo0A regulatory proteins from cells lysate of the Escherichia coli / A.M. Cheryomin, A.R. Kayumov, A.A. Toymentseva, N. Pina, M.R. Sharipova // Materials of IV Russian symposium called “Protein and Peptides”. –Kazan, 2009. -P.310.
- Toymentseva A.A. The Expression of aprBi and gseBi genes under Control of different DNA-binding Proteins in Bacillus intermedius / A. Toymentseva, M. Sharipova, T. Mascher // "3rd Joint Conference of the German Society for Hygiene and Microbiology (DGHM) and the Association for General and Applied Microbiology (VAAM)". -Hannover, Germany, 2009. -P.121.
- Тойменцева А.А. Филогенетическое положение штамма Bacillus intermedius 3-19 / А.А. Тойменцева, А.Д. Мухаметзянова, А.И. Ахметова, М.Р. Шарипова // Сборник тезисов Российской школы для молодых ученых «Актуальные проблемы современной биохимии и молекулярной биологии». Казань, 2010. –С.59.
- Тойменцева А.А. Влияние факторов транскрипции на экспрессию генов сериновых протеаз Bacillus pumilus 3-19 / Ч. Нямсурэн, О.Н. Игонина, А.В. Гончарова, А.А. Тойменцева, Е.И. Шагимарданова, М.Р. Шарипова // X Научная конференция молодых учёных, аспирантов и студентов научно-образовательного центра Казанского (Приволжского) федерального университета. Казань, 2011. –С.77.
- Nyamsuren C. Study of the Impact of Major Transcription Factors on Gene Expression of Serine Proteases from Bacillus Pumilus 3-19 by Using Fusion-Design / C. Nyamsuren, O. Igonina, A. Goncharova, A. Toymentseva, M. Sharipova // Students, Masters and PhD Students III Annual International Scientific Conference in Biology. Ivane Javakhishvili Tbilisi State University. –Tbilisi, Georgia, 2011. -P.69-70.
- Гончарова А.В. Экспрессия fusion-конструкции под контролем промотера гена субтилизиноподобной протеиназы Bacillus pumilus 3-19 в регуляторных мутантах / А.В. Гончарова, О.Н. Игонина, А.А. Тойменцева, М.Р. Шарипова // Итоговая научно-образовательная конференция студентов Казанского (Приволжского) федерального университета. -Казань, 2011. –С.6-7.
- Нямсурэн Ч. Экспрессия fusion-конструкции на основе бетта-галактозидазы под промотором гена глутамилэндопептидазы Bacillus pumilus / Ч. Нямсурэн, А.А. Тойменцева, М.Р. Шарипова // Итоговая научно-образовательная конференция студентов Казанского (Приволжского) федерального университета. -Казань, 2011. –С.15-16.
- Игонина О.Н. Субклонирование гена адамализиноподобной металлоэндопептидазы mprBp B. pumilus / О.Н. Игонина, А.А. Тойменцева, М.Р. Шарипова // XI Научная конференция молодых учёных, аспирантов и студентов научно-образоватльного центра Казанского (Приволжского) федерального университета «Материалы и технологии XXI века». –Казань, 2012. –С.34.
- Нямсурэн Ч. Конструирование системы экспрессии для B. subtilis на основе Lia-промотора / Ч. Нямсурэн, А.А. Тойменцева, М.Р. Шарипова // XI Научная конференция молодых учёных, аспирантов и студентов научно-образоватльного центра Казанского (Приволжского) федерального университета «Материалы и технологии XXI века». –Казань, 2012. –С.51.
- Тойменцева А.А. Регуляция экспрессии генов протеаз Bacillus pumilus на основе репортёрной конструкции / А.А. Тойменцева, М.Р. Шарипова // XI Научная конференция молодых учёных, аспирантов и студентов научно-образоватльного центра Казанского (Приволжского) федерального университета «Материалы и технологии XXI века» - Казань, -2012. –С.67.
- Нямсурэн Ч. Модификация бактериальной системы экспрессии на основеподбора сигнального пептида / Ч. Нямсурэн, А.А. Тойменцева // Международная конференция студентов, аспирантов и молодых учёных «ЛОМОНОСОВ-2012». - Москва, 2012. –С.161.
Благодарности. Автор выражает глубокую признательность научному руководителю д.б.н. профессору М.Р. Шариповой за внимательное отношение к работе и обсуждение полученных результатов; к.б.н., доц. А.М. Мардановой и к.б.н., с.н.с. Н.П. Балабан за постоянные консультации и обсуждение результатов; профессору T. Masher за сотрудничество по созданию новой антибиотико-индуцибельной системы и предоставление для работы лабораторных штаммов и плазмид, а также за возможность проведения части экспериментов в лаборатории Мюнхенского университета Людвига-Максимилиана (LMU, Германия). Автор выражает искреннюю благодарность заведующей кафедрой микробиологии Казанского (Приволжского) федерального университета, профессору О.Н. Ильинской и сотрудникам кафедры микробиологии за помощь и доброжелательную рабочую атмосферу.
Отзывы на автореферат просим высылать по адресу: Казань, 420008, ул. Кремлевская, 18, Казанский университет, отдел аттестации, Ученому секретарю Диссертационного совета Д 212.081.08 Абрамовой Зинаиде Ивановне и по факсу: (843) 238-76-01