

Вовлечение кальпаиновой системы в аутоиммунные нейродегенеративные процессы (на модели аллергического энцефаломиелита крыс)
На правах рукописи
Карпенко
Марина Николаевна
Вовлечение кальпаиновой системы в аутоиммунные нейродегенеративные процессы
(на модели аллергического энцефаломиелита крыс)
03.00.13 — физиология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
САНКТ-ПЕТЕРБУРГ
2009
Работа выполнена в Физиологическом отделе им. И.П. Павлова
ГУ Научно-исследовательский Институт экспериментальной медицины РАМН
(ГУ НИИЭМ РАМН)
Научный руководитель:
доктор медицинских наук, профессор
Виктор Матвеевич Клименко
Официальные оппоненты:
доктор биологических наук, профессор
Нера Яковлевна Лукомская
доктор медицинских наук, профессор
Андрей Федорович Якимовский
Ведущее научное учреждение: Институт физиологии им. И.П. Павлова РАН
Защита состоится «26» марта 2009 г. в часов на заседании совета Д001.022.03 по защите докторских диссертаций при ГУ Научно-исследовательский институт экспериментальной медицины РАМН по адресу: 197376 Санкт-Петербург, Каменноостровский пр., д. 69/71.
С диссертацией можно ознакомиться в научной библиотеке ГУ НИИЭМ РАМН (197376 Санкт-Петербург, ул. Академика Павлова, д. 12).
Автореферат разослан « » февраля 2009 г.
Ученый секретарь Диссертационного совета
доктор биологических наук, профессор Л.В. Пучкова
ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Ограниченный протеолиз белков и пептидов в живых тканях и клетках является особой формой физиологической регуляции и считается универсальным механизмом, ответственным за образование и модификацию ферментов. Реакции ограниченного протеолиза лежат в основе функционирования важнейших физиологических систем, в частности, иммунитета, гемостаза, апоптоза, регуляции экспрессии генов (Чипенс и др., 1985). Поэтому для понимания принципов регуляции физиологических систем необходимо детальное изучение внутриклеточных регуляторных протеаз и тех реакций, которые они катализируют.
Среди внутриклеточных протеолитических систем особого внимания заслуживают те из них, которые активируются в ответ на увеличение в клетке концентрации ионов кальция – ключевого посредника множества клеточных реакций. Однако при развитии ряда патологических процессов существует опасность чрезмерной активации кальций-зависимых протеолитических систем вследствие нарушения гомеостаза ионов кальция. (Lajtha, Banik, 2002). С этой точки зрения особый интерес вызывает Са2+-зависимая кальпаиновая система внутриклеточных протеаз, гиперактивация которой приводит к нарушению регуляции многих физиологических функций организма (Goll et al., 2003). Например, при развитии аутоиммунных нейродегенеративных процессов у людей и при индукции этих процессов у лабораторных животных, в частности, у животных с индуцированным экспериментальным аллергическим энцефаломиелитом (ЭАЭ) в иммуноцитах и клетках ЦНС наблюдается увеличение внутриклеточной концентрации ионов кальция (Lajtha, Banik, 2002). На основании вышеприведенных данных, логично предположить, что в этих клетках кальпаины могут проявлять повышенную протеолитическую активность, сказывающуюся на всех этапах патогенеза: от регуляции миграции активированных Т-лимфоцитов и расщепления белков миелина до активации проапоптотических ферментов. Действительно, расщепление кальпаином основного белка миелина, белка нейрофиламентов может стимулировать образование энцефалитогенных пептидов; расщепление прокаспаз до каспаз способствует запуску апоптотических каскадов; расщепление белков клеточной адгезии увеличивает миграционную способность клетки (Goll et al., 2003). Кроме этого, выявление новых субстратов кальпаина расширяет представление о функциональной роли кальпаиновой системы. Так, недавно было показано, что нейроспецифический белок GAP-43 (growth associated protein) является субстратом кальпаина (Захаров и др., 2005). Поскольку белок GAP-43 известен как маркер регенеративных процессов, протекающих в нервной ткани (Oestreicher et al., 1997), весьма вероятно, что протеолитическое расщепление белка GAP-43 кальпаином в условиях развития аутоиммунного нейродегенеративного процесса может, существенно изменяя его свойства, подавить регенеративную способность клеток ЦНС и усугубить течение заболевания. Приведенные факты свидетельствуют о том, что исследование вовлеченности кальпаиновой системы иммуноцитов и клеток ЦНС в аутоиммунные нейродегенеративные процессы является одним из наиболее актуальных направлений клеточной физиологии.
Цели и задачи исследования.
Цель работы: показать включение кальпаиновой системы клеток ЦНС и иммунной системы в развитие аутоиммунных нейродегенеративных процессов на модели ЭАЭ у крыс.
Задачи исследования были сформулированы следующим образом:
- Определить уровень экспрессии компонентов кальпаиновой системы и активность кальпаинов в иммуноцитах и клетках ЦНС в индуктивную фазу ЭАЭ.
- Определить уровень экспрессии компонентов кальпаиновой системы и активность кальпаинов в иммуноцитах и клетках различных отделов ЦНС на пике развития ЭАЭ. Сопоставить полученные данные с тяжестью клинического течения ЭАЭ.
- Исследовать действие ингибитора кальпаина I на выраженность клинических проявлений ЭАЭ.
- Определить уровень экспрессии мРНК GAP-43, а также изучить распределение белка GAP-43 и его протеолитического фрагмента в клетках спинного мозга на пике развития ЭАЭ. Сопоставить полученные данные с тяжестью клинического течения ЭАЭ.
- Определить уровень белка GAP-43 в ликворе крыс в процессе развития ЭАЭ при различной тяжести течения болезни.
Основные положения, выносимые на защиту.
- Кальпаиновая система клеток ЦНС и иммунной системы вовлечена в процесс формирования и развития нейродегенеративной патологии аутоиммунного генеза.
- Увеличение уровня белка GAP-43 в ликворе крыс с ЭАЭ и увеличение степени его протеолиза кальпаином в клетках ЦНС отражает тяжесть течения заболевания, что может служить диагностическим и/или прогностическим критерием развития и интенсивности аутоиммунного нейродегенеративного процесса.
- Подавление активности кальпаинов с помощью специфических ингибиторов является перспективным направлением для разработки новых подходов в лечении аутоиммунных нейродегенеративных заболеваний.
Научная новизна работы. В проведенном исследовании впервые получены данные о включении кальпаиновой системы клеток лимфатических узлов брыжейки и верхних отделов спинного мозга в процесс формирования и развития нейродегенеративной патологии аутоиммунного генеза. Выявлено, что распространенность гиперактивации и сверхэкспрессии m-кальпаина по различным отделам спинного мозга крыс связана с тяжестью течения ЭАЭ. Впервые показано увеличение уровня экспрессии мРНК кальпастатина и нейроспецифического белка GAP-43 в клетках различных отделов спинного мозга крыс с бессимптомным и легким течением ЭАЭ, а также обнаружено увеличение степени протеолитического расщепления белка GAP-43 кальпаином в клетках различных отделов спинного мозга крыс с тяжелой формой ЭАЭ. Установлено увеличение содержания белка GAP-43 и его протеолитических фрагментов в ликворе крыс с симптомами ЭАЭ на пике заболевания. Впервые продемонстрировано, что подавление активности кальпаинов с помощью интраназального введения низких доз ингибитора кальпаина I предотвращает развитие или снижает выраженность ЭАЭ.
Научно-практическое значение результатов работы. Выявленное в данном исследовании включение кальпаиновой системы клеток ЦНС и иммунной системы в развитие ЭАЭ у крыс углубляет представление о физиологических функциях кальпаинов и раскрывает ряд неизвестных ранее звеньев механизма формирования аутоиммунных нейродегенеративных процессов. Предотвращение развития или снижение выраженности ЭАЭ у крыс в результате подавления активности кальпаинов с помощью ингибитора кальпаина I указывает на возможность использования препаратов на основе ингибиторов кальпаина для лечения аутоиммунных нейродегенеративных заболеваний и других форм патологии ЦНС, связанных с деструкцией нервных клеток. Данные о протеолизе белка GAP-43 под действием кальпаина существенно расширяют и дополняют сложившиеся к настоящему времени представления о механизмах функционирования белка GAP-43 в условиях развития нейродегенеративного процесса. На основе полученных данных об изменении уровня белка GAP-43 и его протеолитических фрагментов в ликворе могут быть разработаны новые подходы для диагностики и прогнозирования течения аутоиммунного нейродегенеративного процесса в клинике.
Личный вклад соискателя. Соискатель проанализировал литературу по теме исследования, осуществил планирование экспериментов и получил основную часть результатов, которые представил в докладах на конференциях и в статьях. Экспериментальная часть исследования полностью выполнена на базе Физиологического отдела им. И.П. Павлова ГУ НИИЭМ РАМН. Индукция ЭАЭ и оценка клинического течения заболевания проводилась совместно с к.б.н. И.Н. Абдурасуловой (Физиологический отдел им. И.П. Павлова ГУ НИИЭМ РАМН) и к.м.н. Ю.Л. Житнухиным (Отдел иммунологии ГУ НИИЭМ РАМН).
Апробация результатов работы. Материалы, изложенные в диссертации, были представлены на VI Сибирском Физиологическом съезде (Барнаул, 2008); IV съезде Российского общества биохимиков и молекулярных биологов (Новосибирск, 2008); Объединенном иммунологическом форуме (Санкт-Петербург, 2008); 11-th Multidisciplinary International Neuroscience and Biological Psychiatry Conference “Stress and behavior” (St.-Petersburg, 2008); XLVII неделе науки Санкт-Петербургского государственного политехнического университета (Санкт-Петербург, 2008); Политехническом симпозиуме: Молодые ученые – промышленности Северо-Западного региона (Санкт-Петербург, 2008).
Диссертационная работа была апробирована на научном заседании Физиологического отдела им. И.П. Павлова ГУ НИИЭМ РАМН 22 октября 2008 г.
Публикации. По материалам диссертации опубликовано 8 работ.
Структура диссертации. Работа содержит введение, обзор литературы, описание материалов и методов, результаты и их обсуждение, заключение, выводы и список цитируемой литературы, включающий 314 источников. Материалы диссертации изложены на 177 страницах машинописного текста и иллюстрированы 30 рисунками и одной таблицей.
СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы исследования. Работа выполнена на 255 самках крыс породы Вистар (Рапполово) весом 240-250 грамм. ЭАЭ вызывали однократной инокуляцией энцефалитогенной смеси из расчета 100 мг гомогената гомологичного спинного мозга, 0,2 мл полного адъюванта Фрейнда (содержание убитых микобактерий 5 мг/мл) (ПАФ) и 0,2 мл физиологического раствора на одно животное. Эцефалитогенную смесь в объеме 0,4 мл вводили подкожно (в основание хвоста) под легким эфирным наркозом. Контрольным животным инъецировали аналогичный объем ПАФ. Ежедневно животных взвешивали и оценивали наличие клинических симптомов ЭАЭ по методу, разработанному Абдурасуловой И.Н. (Отдел физиологии им. И.П. Павлова, ГУ НИИЭМ РАМН). Для этого регистрировали время начала заболевания, его продолжительность и тяжесть, которую оценивали в баллах по наличию у животных мышечной слабости (0,5 балла), стойких парезов (1 балл) и параличей (1,5 балла). При поражении нескольких конечностей баллы суммировались для расчета клинического индекса (КИ). Отсутствие видимых клинических проявлений оценивалось как КИ =0, летальный исход – КИ = 6. Животных с КИ = 0,5-2,0 считали легко болеющими, животных с КИ = 2,5-3,5 – средней тяжести заболевания, животных с КИ = 4,0-5,0 – тяжело болеющими, КИ= 5,5-6,0 – крайне тяжело болеющими. Рассчитывали кумулятивный КИ – сумма всех КИ на протяжении заболевания.
На 3, 5, 7 сутки после индукции ЭАЭ (индуктивная фаза) декапитировали по пять животных опытной и контрольной групп; на 14 день (пик заболевания) декапитировали пять животных контрольной группы и по пять животных с различной тяжестью течения ЭАЭ. В качестве пассивного контроля (контрольная группа I) использовали интактных животных; вторую контрольную группу составили животные, получившие инъекцию ПАФ в день индукции ЭАЭ у животных экспериментальных групп (контрольная группа II). Выделяли спинной мозг, мозжечок, селезенку, лимфатические узлы брыжейки. За день до индукции и на 7, 14, 21 день после индукции ЭАЭ из мозжечково-мозговой цистерны отбирали до 100 мкл СМЖ у 5 контрольных и 25 опытных животных, СМЖ хранили при -200С.
Для исследования экспрессии мРНК - и m-кальпаина, кальпастатина и GAP-43 из полученных тканей выделяли тотальную мРНК в соответствии со стандартным протоколом (Sambrook et al., 1989) с использованием гуанидина тиоционата (Promega, США). Для проведения полимеразной цепной реакции (ПЦР) на полученных кДНК использовали уникальные праймеры, подобранные по программе Primer 3.0. Детали проведения анализа описаны ранее (Карпенко и др., 2008). Активность кальпаина измеряли с помощью коммерческого набора, с использованием флуоресцентно меченного синтетического субстрата кальпаина (BioVision, США). Все процедуры проводили согласно инструкции производителя. Результаты представляли в оптических единицах флюоресценции на мкг белка.
Выделение белка GAP-43 осуществляли с помощью экстракции 1%-ным тритоном Х-100 и 5%-ной хлорной кислотой из гомогенатов тканей спинного мозга (Mosevitsky et al., 1994). Электрофоретическое разделение белков проводили в системе уксусная кислота/мочевина по методике Панима и Чокли (Panyim, Chalkley, 1969). Перенос белков из полиакриламидного геля на нитроцеллюлозный мембранный фильтр (Synpore, Chemapol, размер пор 0,1-0,4 мкм) осуществляли методом "поверхностного отпечатка" (Plekhanov, 1996). В качестве первых антител использовали моноклональные антитела к белку GAP-43 (Sigma, США); в качестве вторых антител использовали антимышиные козьи гамма-иммуноглобулины, конъюгированные с пероксидазой хрена. Иммунные комплексы визуализировали хемилюминесцентным методом. Иммуноферментный анализ проводили по стандартной схеме (Егоров и др., 1991).
Значимость различий между уровнями мРНК кальпаинов, кальпастатина и GAP-43 в сравниваемых группах животных устанавливали с использованием одномерного дисперсионного анализа с последующим применением критерия Тьюки. Достоверность различий длительности латентного периода и клинической фазы ЭАЭ, а также значимость различий клинического и кумулятивного индекса у тестируемых групп крыс по сравнению с контролем устанавливалась с помощью непараметрического критерия Манна-Уитни.
Результаты исследования и их обсуждение.
Характеристика развития ЭАЭ.
Однократное подкожное введение гомогената гомологичного спинного мозга в ПАФ вызывало у самок крыс породы Вистар развитие монофазного ЭАЭ различной тяжести. По результатам трех независимых экспериментов (n=90) заболеваемость животных составила 93,3%. Легкое течение заболевания (КИ = 0,5-2,0) наблюдалось у 14,3% крыс; средней тяжести (КИ = 2,5-3,5) – у 28,5%; тяжелое (КИ = 4,0-5,0) – у 12,5%; крайне тяжелое (КИ = 5,5-6,0) – у 44,7% крыс, из них у 9,8 % заболевание окончилось летально. Клинические симптомы ЭАЭ у крыс появлялись через 6,9±0,8 суток, это наблюдение позволило ограничить латентную фазу заболевания первыми семью днями после индукции ЭАЭ. Максимальный КИ у животных экспериментальной группы наблюдался на 14-й д.п.и. (дней после индукции ЭАЭ), поэтому этот день считали пиком заболевания. Длительность заболевания различалась при легком и тяжелом течении и составила 5,2±2,3 и 21,4±1,8 суток соответственно. В дальнейшем у большинства животных наступало полное или частичное спонтанное выздоровление, поэтому весь период наблюдения был ограничен 30 днями. У животных контрольной группы II (введение ПАФ) в течение указанного периода симптомы ЭАЭ не выявлялись.
Различная тяжесть заболевания у самок крыс Вистар, по-видимому, отражает генетическую разнородность используемых нами животных, подтверждая роль генетических факторов в предрасположенности к аутоиммунным заболеваниям.
Проведенные нами исследования, а также данные, полученные коллегами ранее, показали, что используемая модель ЭАЭ является адекватной для изучения аутоиммунных нейродегенеративных процессов. На это указывает несколько фактов. Во-первых, выявлено сходство клинических симптомов ЭАЭ и аутоиммунного нейродегенеративного заболевания человека - рассеянного склероза (мышечная слабость, парезы и параличи конечностей). Во-вторых, показана общность патогенетических механизмов при ЭАЭ и при нейродегенеративных заболеваниях аутоиммунного генеза человека, которая доказывается сходством морфологических и метаболических изменений в ЦНС (Абдурасулова и др., 2004).
Экспрессия мРНК и активность кальпаинов в иммуноцитах и клетках ЦНС в индуктивную фазу ЭАЭ.
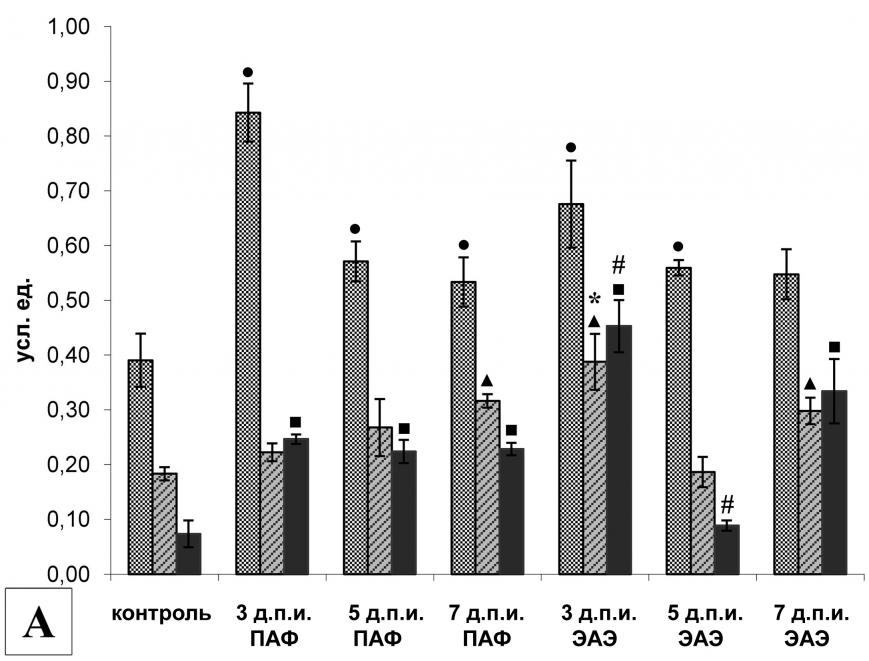 |  | |
| Рис. 1. | Экспрессия мРНК (А) и активность (Б) кальпаинов в клетках селезенки крыс в индуктивную фазу ЭАЭ. | |
| ,, | - р<0,05 по отношению к соответствующему контролю (А). | |
| *, # | - р<0,05 по отношению к соответствующему контролю в группе ПАФ (А). | |
| * | - р<0,05 по отношению к контролю (Б). | |
Анализ экспрессии мРНК компонентов кальпаиновой системы в клетках селезенки крыс в индуктивную фазу ЭАЭ показал, что процесс развития заболевания сопровождается изменением уровня экспрессии мРНК -, m-кальпаина и кальпастатина, причем динамика экспрессии - и m-кальпаинов у крыс с ЭАЭ оказалась сходной, но отличалась от контрольной группы II (введение ПАФ). Действительно, на 3-й д.п.и. отмечалось увеличение экспрессии мРНК обеих протеаз по сравнению с животными контрольных групп. На 5-й д.п.и. уровень экспрессии мРНК кальпаинов у животных экспериментальной группы снижался до уровня контрольной группы I (рис. 1А). На 7-й д.п.и. наблюдалось достоверное увеличение уровня экспрессии мРНК кальпаинов по сравнению с контрольной группой I, однако, достоверных отличий от контрольной группы II не наблюдалось. Динамика экспрессии мРНК кальпастатина у крыс с ЭАЭ и крыс, получавших ПАФ, была сходной: на всех анализируемых сроках уровень экспрессии мРНК кальпастатина был достоверно выше, чем в контрольной группе I, однако на 3-й д.п.и. у экспериментальных крыс увеличение экспрессии мРНК кальпастатина было менее выражено, чем у крыс контрольной группы II.
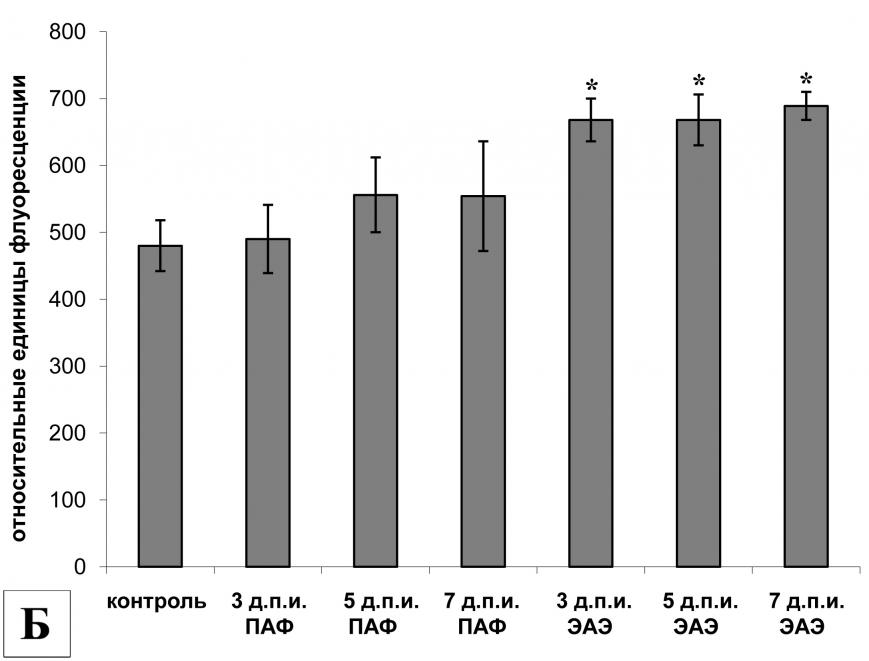
Анализ активности кальпаинов в клетках селезенки выявил высокую базовую активность кальпаинов у интактных животных (рис. 1Б). Стимуляция иммунной системы с помощью ПАФ не вызвала изменений активности кальпаинов в этих клетках. В клетках селезенки экспериментальных животных на 3-й и 5-й д.п.и. активность кальпаинов также была высокой и не отличалась от контрольных групп. Однако к 7-му д.п.и. активность кальпаинов достоверно снизилась по сравнению с интактными животными.
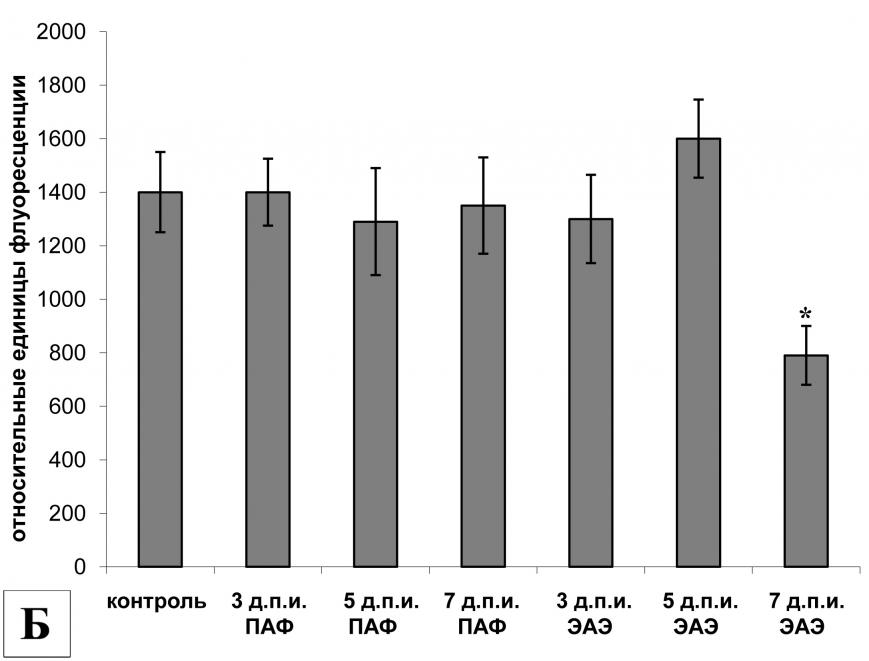
 |  | |
| Рис. 2. | Экспрессия мРНК (А) и активность (Б) кальпаинов в клетках лимфатических узлов брыжейки крыс в индуктивную фазу ЭАЭ. | |
| ,, | - р<0,05 по отношению к соответствующему контролю (А). | |
| * | - р<0,05 по отношению к соответствующему контролю в группе ПАФ (А). | |
| * | - р<0,05 по отношению к контролю (Б). | |
В клетках лимфатических узлов брыжейки крыс с ЭАЭ динамика экспрессии - и m-кальпаинов, как и в клетках селезенки, оказалась сходной (рис. 2А). На 3-й д.п.и. отмечалось достоверное увеличение экспрессии мРНК обеих протеаз по сравнению с животными контрольной группы I. На 5-й д.п.и. экспрессия мРНК кальпаинов экспериментальной группы оставалась на повышенном уровне, а на 7-й д.п.и. уже не отличалась от контрольной группы I. Однако в клетках лимфатических узлов брыжейки крыс контрольной группы II (животные, получившие инъекцию ПАФ) также наблюдалось увеличение экспрессии мРНК - и m-кальпаинов, а на 7-й д.п.и. уровень экспрессии m-кальпаина в этой группе был даже достоверно выше, чем у крыс с ЭАЭ. Динамика экспрессии мРНК кальпастатина у крыс с ЭАЭ и крыс, получавших ПАФ, совпадала: у животных обеих групп выявлялось достоверное увеличение экспрессии мРНК кальпастатина на 3-й и 5-й д.п.и., а на 7-й д.п.и. уровень экспрессии кальпастатина не отличался от контрольной группы I (интактные животные).
Анализ активности кальпаинов в клетках лимфатических узлов брыжейки крыс с ЭАЭ показал, что на 3-й д.п.и. уровень активности кальпаинов был сравним с базовым, но к 5-му д.п.и. активность кальпаинов достоверно увеличилась по отношению к пассивному контролю (контрольная группа I). Однако уже к 7-му д.п.и. активность снизилась до уровня контрольной группы II, но была достоверно выше, чем у интактных животных (рис. 2Б).
В клетках ЦНС животных с ЭАЭ нами была выявлена иная динамика изменения уровня экспрессии мРНК и активности кальпаинов. Так, в клетках ЦНС уровень экспрессии мРНК -кальпаина практически не изменялся. Увеличение экспрессии мРНК -кальпаина отмечалось лишь в клетках поясничного отдела спинного мозга крыс экспериментальной группы, причем исключительно на 7-й д.п.и. При этом экспрессия мРНК m-кальпаина у крыс с ЭАЭ по сравнению с животными контрольных групп I и II усиливалась уже на 3-й д.п.и. и сохранялась на повышенном уровне вплоть до 7-го д.п.и. Экспрессия мРНК кальпастатина, напротив, увеличивалась только у крыс группы ПАФ (рис. 3А).
Протеолитическая активность кальпаинов в клетках поясничного отдела спинного мозга крыс с ЭАЭ также была повышена, причем достоверное увеличение активности кальпаинов наблюдалось уже на 3-й д.п.и. В этих клетках гиперактивация кальпаиновой системы и повышенный уровень экспрессии мРНК m-кальпаина выявлялись на протяжении всего латентного периода (рис. 3Б).
 |  | |
| Рис. 3. | Экспрессия мРНК (А) и активность (Б) кальпаинов в клетках поясничного отдела спинного мозга крыс в индуктивную фазу ЭАЭ. | |
| ,, | - р<0,05 по отношению к соответствующему контролю (А). | |
| * | - р<0,05 по отношению к контролю (Б). | |
Различий в уровне экспрессии мРНК кальпаинов и кальпастатина, а также в уровне протеолитической активности кальпаинов в грудном, шейном отделе спинного мозга крыс с ЭАЭ выявлено не было. Таким образом, в индуктивную фазу ЭАЭ в иммуноцитах и клетках поясничного отдела спинного мозга крыс наблюдается дисбаланс экспрессии мРНК компонентов кальпаиновой системы; в клетках поясничного отдела спинного мозга также выявляется гиперактивация кальпаинов.
Полученные нами данные указывают на различную динамику вовлечения кальпаиновой системы иммуноцитов и клеток ЦНС в развитие ЭАЭ на ранних стадиях формирования патологии. Так, в клетках селезенки и лимфатических узлов брыжейки уже на 3-й д.п.и. происходит увеличение уровня экспрессии мРНК всех анализируемых представителей кальпаиновой системы, включая кальпастатин, что, вероятно, обеспечивает поддержание базовой активности кальпаинов в этих клетках. В клетках поясничного отдела спинного мозга, напротив, при увеличении уровня экспрессии мРНК m-кальпаина изменение уровня экспрессии мРНК кальпастатина не наблюдается. Это, возможно, и приводит к гиперактивации кальпаинов в клетках ЦНС уже на 3-й д.п.и.
Обнаруженное повышение уровня экспрессии кальпаинов и гиперактивация кальпаинов в клетках поясничного отдела спинного мозга крыс в индуктивную фазу ЭАЭ, вероятнее всего, характерны для резидентных и мигрировавших в ЦНС клеток иммунной системы. В пользу этого предположения свидетельствует показанное нами отсутствие увеличения степени протеолиза кальпаином нейроспецифического белка GAP-43, а также обнаружение в клетках поясничного отдела спинного мозга крыс многократного увеличения уровня экспрессии -кальпаина (на 7-й д.п.и.), который в норме в нейронах и клетках глии практически не экспрессируется, но экспрессируется активированными Т-лимфоцитами (Schaecher et al., 2002). Известно, что одним из субстратов -кальпаина является ингибитор транскрипционного фактора NF-kB – белок IB, расщепление которого способствует перемещению транскрипционного фактора NF-B из цитозоля в ядро, где он активирует транскрипцию многих генов, в том числе гена индуцибельной NOS, циклооксигеназы-2а, ФНО-, ИЛ-1 (Ruetten, Thiemermann, 1997). Активация транскрипции этих генов приводит к повышению уровня продукции соответствующих белков, которые индуцируют развитие в ЦНС воспалительной реакции (Baumann, Pham-Dinh, 2001). Таким образом, сверхэкспрессия и гиперактивация -кальпаина в клетках поясничного отдела спинного мозга может способствовать созданию пермиссивной среды для развития воспалительной реакции в ЦНС.
Экспрессия мРНК и активность кальпаинов в иммуноцитах и клетках ЦНС животных с различной тяжестью течения ЭАЭ
Анализ уровня экспрессии мРНК - и m-кальпаина, а также кальпастатина в клетках селезенки и лимфатических узлов брыжейки крыс с различной тяжестью течения ЭАЭ показал, что на 14-й д.п.и. уровень экспрессии анализируемых компонентов кальпаиновой системы не отличался от базового; протеолитическая активность кальпаинов также была на уровне контрольных значений. Следовательно, вне зависимости от тяжести течения заболевания экспрессия мРНК -, m-кальпаина и кальпастатина, а также уровень активности кальпаинов к 14-му д.п.и. возвращаются к исходному базовому значению.
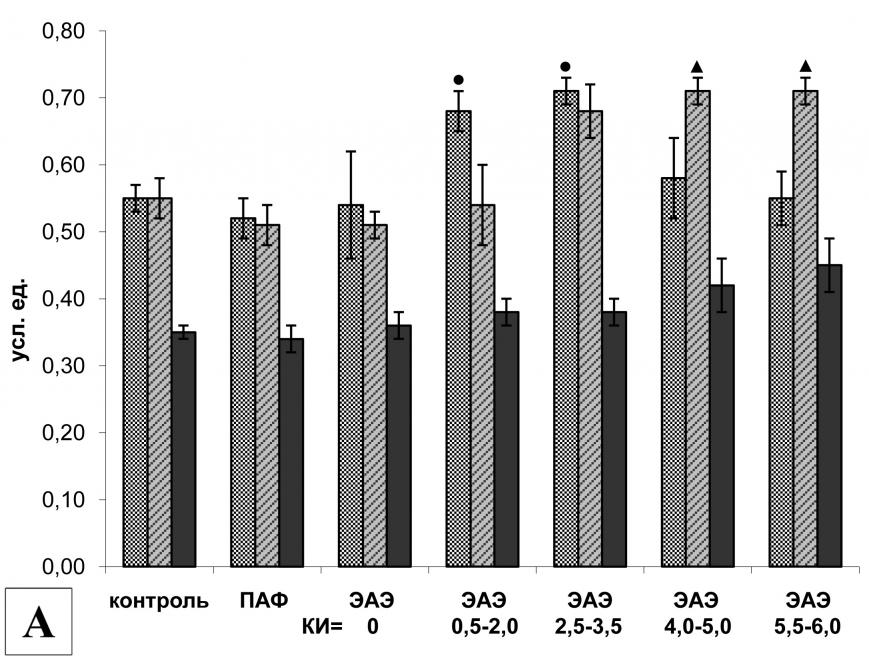
Анализ экспрессии мРНК компонентов кальпаиновой системы в клетках ЦНС показал преимущественное вовлечение в процесс развития ЭАЭ m-кальпаина и кальпастатина. Действительно, в клетках поясничного отдела спинного мозга крыс с различной тяжестью течения ЭАЭ и у животных контрольных групп обнаружен практически одинаковый уровень экспрессии мРНК -кальпаина. Однако уровень экспрессии мРНК m-кальпаина и кальпастатина у крыс экспериментальных групп заметно различался. Так, на 14-й д.п.и. у животных контрольной группы II (введение ПАФ) в клетках поясничного отдела спинного мозга выявлялось достоверное увеличение экспрессии мРНК m-кальпаина по сравнению с интактными животными при сохранении базового уровня экспрессии мРНК кальпастатина. У животных без видимых клинических проявлений заболевания и животных с легкой формой ЭАЭ уровень экспрессии мРНК m-кальпаина был достоверно выше, чем в контрольной группе I, но не отличался от контрольной группы II. При развитии ЭАЭ средней тяжести, а также при тяжелых формах заболевания, уровень экспрессии мРНК m-кальпаина увеличивался и был достоверно выше, чем в обеих контрольных группах. Экспрессия m-кальпаина достигала наибольшего значения в группе с крайне тяжелой формой ЭАЭ (КИ= 5,5-6,0). Напротив, экспрессия мРНК кальпастатина достоверно увеличивалась только у крыс с легкой формой заболевания (рис. 4А).
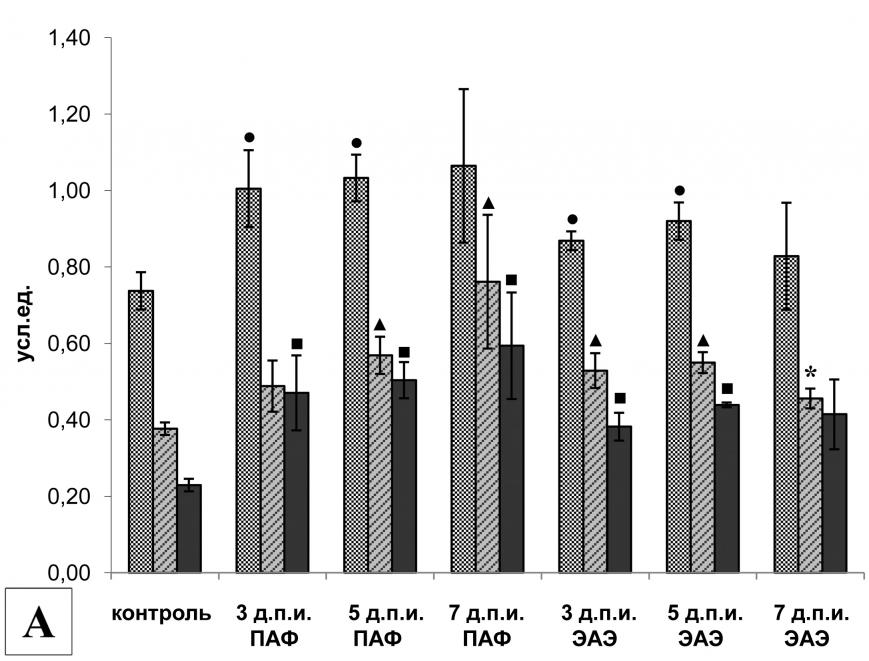
 |  | |
| Рис. 4. | Экспрессия мРНК (А) и активность (Б) кальпаинов в клетках поясничного отдела спинного мозга крыс с различной тяжестью течения ЭАЭ (14-й д.п.и.). | |
| , | - р<0,05 по отношению к соответствующему контролю (А). | |
| * | - р<0,05 по отношению к соответствующему контролю в группе ПАФ (А). | |
| * | - р<0,05 по отношению к контролю (Б). | |
| ** | - р<0,001 по отношению к контролю (Б). | |
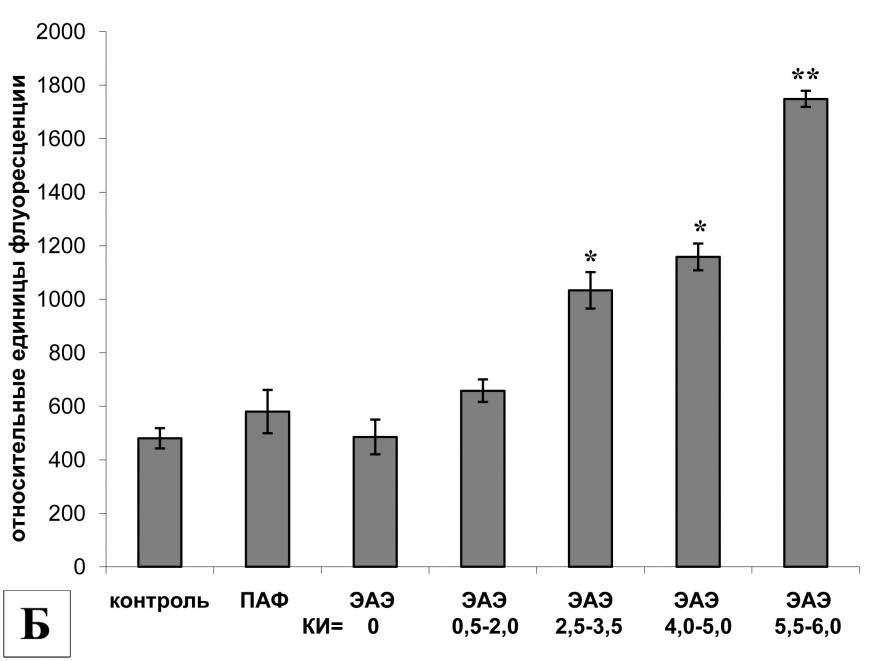
Протеолитическая активность кальпаина в клетках поясничного отдела спинного мозга крыс экспериментальных групп также различалась. У животных без видимых неврологических нарушений и крыс с легким течением ЭАЭ активность кальпаинов сохранялась на базовом уровне. Однако даже у этих животных мы наблюдали повышение экспрессии мРНК m-кальпаина, которое сопровождалось увеличением уровня экспрессии мРНК кальпастатина (эндогенного ингибитора кальпаина). Вероятно, поэтому уровень протеолитической активности кальпаинов практически не отличался от интактных животных. Повышенная ферментативная активность кальпаина наблюдалась только у животных со стойкими неврологическими нарушениями в виде парезов и параличей одной или нескольких конечностей, причем активность кальпаинов в клетках поясничного отдела спинного мозга увеличивалась с повышением тяжести течения заболевания и достигала наибольшего значения у животных с крайне тяжелой формой ЭАЭ (рис. 4Б).
В клетках грудного отдела спинного мозга уровень экспрессии мРНК m-кальпаина у животных без признаков ЭАЭ, у крыс с легкой и средней формой ЭАЭ не отличался от контрольных групп. Однако при тяжелых формах заболевания (КИ>4) экспрессия m-кальпаина достоверно увеличилась, при этом протеолитическая активность кальпаинов также была повышена. Следует отметить, что у крыс с КИ = 0,5-2,0 и КИ = 2,5-3,5 наблюдалось увеличение уровня экспрессии мРНК кальпастатина, что, вероятно, привело к снижению протеолитической активности кальпаина. Действительно, у животных с легкой формой ЭАЭ экспрессия мРНК - и m-кальпаина сохранялась на базовом уровне, однако, экспрессия мРНК кальпастатина была повышена, в результате чего протеолитическая активность кальпаинов снизилась и стала даже ниже, чем у животных контрольных групп (рис. 5).
 |  | |
| Рис. 5. | Экспрессия мРНК (А) и активность (Б) кальпаинов в клетках грудного отдела спинного мозга крыс с различной тяжестью течения ЭАЭ (14-й д.п.и.). | |
| , | - р<0,05 по отношению к соответствующему контролю (А). | |
| * | - р<0,05 по отношению к контролю (Б). | |
| ** | - р<0,001 по отношению к контролю (Б). | |
В шейном отделе спинного мозга крыс на пике ЭАЭ повышенный уровень экспрессии мРНК m-кальпаина наблюдался только у экспериментальных животных с крайне тяжелым течением заболевания. Однако у этих животных уровень экспрессии мРНК кальпастатина был также повышен, что, вероятно, предотвратило гиперактивацию кальпаинов. Высокая протеолитическая активность кальпаинов наблюдалась лишь у животных с КИ = 4,0-5,0, причем у этих крыс экспрессия мРНК кальпаинов сохранялась на базовом уровне, зато уровень экспрессии мРНК кальпастатина значительно снизился (данные не показаны).
Таким образом, в клетках спинного мозга животных с ЭАЭ наблюдается сверхэкспрессия m-кальпаина и гиперактивация кальпаинов, причем с усилением тяжести течения заболевания повышение протеолитической активности кальпаинов выявляется не только в нижних, но и в верхних отделах спинного мозга. Поскольку в клетках ЦНС животных с ЭАЭ обнаружено повышение экспрессии мРНК m-, но не -кальпаина, по-видимому, наблюдаемое увеличение протеолитической активности кальпаинов в этих клетках в большей степени опосредовано гиперактивацией именно m-кальпаина.
Известно, что кальпастатин является единственным известным на сегодняшний день эндогенным ингибитором кальпаинов, повышение экспрессии которого, как правило, приводит к подавлению протеолитической активности кальпаинов (Goll et al., 2003). Действительно, анализ уровня экспрессии мРНК кальпастатина и активности кальпаинов в одних и тех же образцах выявил подавление активности кальпаинов при повышении уровня экспрессии кальпастатина. В этом отношении особый интерес представляет обнаруженное нами увеличение уровня экспрессии мРНК кальпастатина в клетках различных отделов спинного мозга крыс с бессимптомным и легким течением ЭАЭ. Таким образом, одним из факторов, определяющих тяжесть течения ЭАЭ у крыс породы Вистар, является степень и распространенность сверхэкспрессии, а также гиперактивации кальпаинов клеток ЦНС; способность клеток ЦНС сверхэкспрессировать кальпастатин обеспечивает развитие легких форм ЭАЭ.
Выявленная нами связь распространения сверхэкспрессии и гиперактивации m-кальпаина с тяжестью течения ЭАЭ согласуется с данными об объеме нейродеструктивных повреждений ЦНС, что позволяет предположить вовлеченность m-кальпаина в процессы инициации гибели нейронов и клеток глии в условиях развития аутоиммунного нейродегенеративного процесса. Действительно, развитие воспалительной реакции сопровождается высвобождением в межклеточную среду провоспалительных цитокинов, лейкотриенов, активных форм кислорода и азота, эйкозаноидов (глутамата, аспартата) и других биологически активных молекул. Повышение во внеклеточном пространстве концентрации этих веществ вызывает нарушение регуляции гомеостаза различных ионов, в том числе, ионов кальция и приводит к гиперактивации кальпаинов в клетках ЦНС. Активированный кальпаин, в свою очередь, расщепляет кальпаин-чувствительные белки (белки клеточной адгезии, белки миелина, некоторые белки клеточного цитоскелета и транскрипционные факторы), вызывая тем самым необратимые повреждения в клетке и в итоге - ее гибель (Goll et al., 2003).
Влияние интраназального введения низких доз ингибитора кальпаина I на выраженность ЭАЭ.
ЭАЭ индуцировали у 35 самок крыс Вистар как описано ранее. За несколько дней до индукции заболевания животных случайным образом распределили на две группы. Численность контрольной группы составила 20 животных, экспериментальной группы – 15 животных. Ингибитор кальпаина I растворили в 2% этиловом спирте. Со 2-го по 16-й д.п.и. крысам экспериментальной группы интраназально вводили ингибитор кальпаина I в дозе 1 мкг/животное в день; крысам контрольной группы аналогичным образом вводили 2% этиловый спирт.
В контрольной группе у 100% крыс наблюдались клинические признаки ЭАЭ, причем у 30% животных заболевание протекало в тяжёлой форме с развитием параличей нескольких конечностей (клинический индекс на пике болезни от 4,0 до 5,5 баллов). У остальных животных этой группы наблюдалось легкое и среднетяжелое течение заболевания, а именно, развивался паралич хвоста и парезы задних лап (клинический индекс на пике болезни от 1,5 до 3,5 баллов). Первые признаки ЭАЭ у животных контрольной группы с тяжёлым течением ЭАЭ проявлялись через 7,2±0,7 дня, у остальных животных через 11,4±1,2 дня; пик болезни наблюдался на 14-й д.п.и. На пике ЭАЭ у животных контрольной группы средний клинический индекс составил 3,4 балла, средний кумулятивный индекс – 23,1 балла; длительность ЭАЭ составила 11,0±0,8 дней (табл. 1.).
| Табл. 1. Влияние курсового введения ингибитора кальпаина I в дозе 1,0 мкг/животное по лечебной схеме (с 1 по 16 день после индукции ЭАЭ) на тяжесть и продолжительность ЭАЭ. | |||||||
| Вещество | Число заболевших животных (%) | Длительность латентного периода ЭАЭ у заболевших крыс (дни)2 | Средний клини-ческий индекс на пике ЭАЭ (баллы)2 | Средний кумуля-тивный индекс (баллы)2 | Длитель-ность ЭАЭ (дни)2 | ||
| Всего1 | Тяжело1 | ||||||
| лёгкое течение | тяжёлое течение | ||||||
| Контроль (2% этанол) | 100 | 30 | 11,4±1,2 | 7,2±0,7 | 3,4 | 23,1 | 11,0±0,8 |
| Ингибитор кальпаина I | 67b | 20 | 13,5±1,9 | 8,1±1,3 | 1,5а | 12,8а | 6,7±1,4а |
| 1 Тест Фишера. | 2 Непараметрический Манн-Уитни тест. | ||||||
| a p<0,05 по сравнению с контролем | b p<0,009 по сравнению с контролем | ||||||
Курсовое введение ингибитора кальпаина I предупреждало развитие ЭАЭ у 33% животных (p<0,009), при этом у заболевших животных в среднем отмечалось более легкое течение ЭАЭ: в 2,2 раза снизился средний клинический индекс на пике заболевания по сравнению с контрольной группой; кумулятивный индекс уменьшился в 1,8 раза (табл. 1.). Несмотря на то, что доля тяжело болеющих животных сохранилась, КИ на пике заболевания у этих крыс снизился. У животных, получавших ингибитор кальпаина I, сократилась длительность заболевания, хотя продолжительность латентного периода не изменилась. Следовательно, интраназальное введение ингибитора кальпаина I в дозе 1 мкг/животное в день в течение 16 дней после индукции ЭАЭ снижало выраженность заболевания.
Таким образом, подавление активности кальпаинов с помощью ингибитора кальпаина I предотвращает развитие или снижает выраженность ЭАЭ у крыс, что подтверждает вовлеченность кальпаиновой системы в патогенез ЭАЭ и указывает на возможность использования препаратов на основе ингибиторов кальпаинов для клинического применения при лечении аутоиммунных нейродегенеративных заболеваний.
Экспрессия мРНК GAP-43, распределение белка GAP-43 и его фрагмента GAP-43-3 в клетках ЦНС в процессе развития ЭАЭ при различной тяжести течения болезни.
На 7-ой д.п.и. ЭАЭ в клетках поясничного отдела спинного мозга нами было выявлено значительное усиление экспрессии мРНК GAP-43. Поскольку усиление экспрессии GAP-43, наблюдаемое после повреждения клеток ЦНС, является маркером регенеративных процессов (Сoggins, Zwiers, 1991), обнаруженный нами факт свидетельствует об инициации регенеративных процессов в поясничном отделе спинного мозга крыс с ЭАЭ уже в индуктивную фазу заболевания.
Исследование степени протеолиза белка GAP-43 кальпаином в гомогенате клеток различных отделов спинного мозга показало, что во всех анализируемых образцах присутствовал целый белок GAP-43 и его протеолитический фрагмент GAP-43-3. Несмотря на обнаруженную нами гиперактивацию кальпаина в клетках поясничного отдела спинного мозга крыс с ЭАЭ, выявляющуюся уже на 3-й д.п.и., доля протеолитического фрагмента GAP-43-3 в этих клетках по сравнению с контрольной группой I и II не увеличилась. Обнаруженный факт имеет следующее объяснение. Мы анализировали активность кальпаина в гомогенате тканей, поэтому не могли определить в каких именно клетках (нейронах, клетках глии или других) наблюдалась гиперактивация кальпаина. Поскольку GAP-43 экспрессируется нейронами и в меньшей степени клетками глии (Oestreicher et al., 1997), весьма вероятно, что гиперактивированный кальпаин и GAP-43 не были солокализованы.
 | |
| Рис. 6. | Уровень экспрессии мРНК GAP-43 в клетках поясничного отдела спинного мозга крыс с различной тяжестью течения ЭАЭ (n=5). |
| * | - р<0,05 по отношению к контролю. |
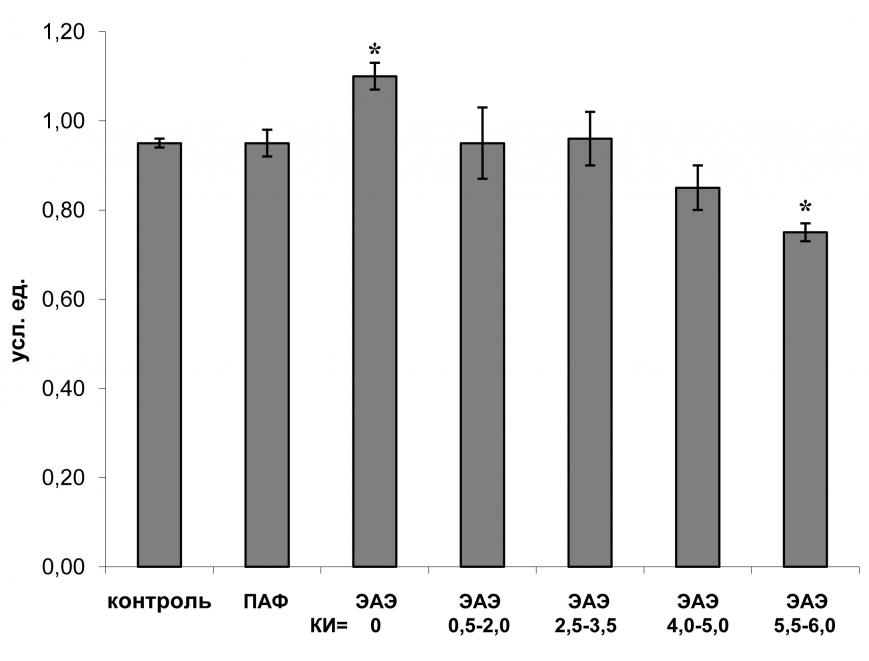
Анализ экспрессии мРНК GAP-43 в клетках поясничного отдела спинного мозга животных с различной тяжестью течения ЭАЭ показал, что уровень экспрессии мРНК GAP-43 связан с тяжестью течения заболевания: у крыс с бессимптомным ЭАЭ (КИ=0) в поясничном отделе спинного мозга наблюдалось достоверное увеличение экспрессии мРНК GAP-43, а при крайне тяжелой форме заболевания (КИ=5,5-6,0), напротив, уровень экспрессии GAP-43 снижался (рис. 6). Поскольку при бессимптомном течении заболевания (КИ=0) повреждено минимальное количество клеток ЦНС, мы полагаем, что эти повреждения могут компенсироваться за счет регенеративных процессов, сопровождающихся выявленным нами усилением экспрессии белка GAP-43. При крайне тяжелом течении ЭАЭ (КИ=5,5-6,0) в поясничном отделе спинного мозга наблюдается массовая гибель нейронов и клеток глии, что приводит к снижению уровня экспрессии мРНК GAP-43.
При развитии ЭАЭ в грудном и шейном отделе спинного мозга нейродегенеративные изменения выявляются только у животных с крайне тяжелой формой ЭАЭ (Steinbrecher et al., 2005). Это согласуется с результатами нашего исследования: уровень экспрессии мРНК GAP-43 в клетках грудного и шейного отдела спинного мозга увеличивается при КИ5,0; остается неизменным или снижается при КИ=5,5-6,0.
 | |
| Рис. 7. | Иммуноблоты, проявленные с помощью коммерческих антител к белку GAP-43 крысы. |
Исследование степени протеолиза белка GAP-43 в гомогенате клеток спинного мозга крыс с различной тяжестью течения ЭАЭ показало, что у всех животных с видимыми неврологическими нарушениями (КИ>0) в клетках поясничного отдела спинного мозга доля GAP-43-3 увеличилась по сравнению с контролем (рис. 7). Так, у интактных животных и крыс контрольной группы II (введение ПАФ) в клетках поясничного отдела спинного мозга в среднем 37 % белка GAP-43 находилось в форме GAP-43-3 (здесь и далее степень протеолиза белка GAP-43 кальпаином вычислена с помощью денситометрирования иммуноблотов; данные представлены в виде процентной доли белка GAP-43-3 от общей интенсивности зон, соответствующих белку GAP-43 и GAP-43-3, n=5). У животных с легким и среднетяжелым течением ЭАЭ доля GAP-43-3 составила уже 60 %, а при тяжелых формах заболевания – 85 %. Это неудивительно, ведь GAP-43 является субстратом кальпаина (Zakharov et al., 2005), а развитие ЭАЭ, как было нами установлено, приводит к гиперактивации кальпаина клеток спинного мозга, причем наибольшая протеолитическая активность кальпаина наблюдается именно в клетках поясничного отдела спинного мозга животных с тяжелым течением ЭАЭ.
В грудном и шейном отделах спинного мозга процентное содержание GAP-43-3 увеличилось по сравнению с контролем лишь в группе животных с тяжелыми формами ЭАЭ, то есть при КИ4,0. Действительно, в клетках грудного и шейного отдела спинного мозга в норме около 40 % белка GAP-43 находилось в форме GAP-43-3; при развитии легкой и среднетяжелой формы заболевания доля GAP-43-3 не изменялась, а при тяжелых и крайне тяжелых формах ЭАЭ доля GAP-43-3 составила уже 65 % и 85 % соответственно (рис. 7). Эти данные полностью согласуются с выявленными нами ранее изменениями уровня активности кальпаинов в клетках грудного и шейного отделов спинного мозга крыс с ЭАЭ. Полученные данные позволяют заключить, что гиперактивация кальпаина, наблюдаемая в клетках различных отделов спинного мозга животных с индуцированным ЭАЭ, приводит к протеолитическому расщеплению белка GAP-43. В зависимости от тяжести течения ЭАЭ степень протеолиза GAP-43 кальпаином увеличивается, причем не только в поясничном отделе спинного мозга, но и в его верхних отделах.
Таким образом, при развитии бессимптомного ЭАЭ в спинном мозге крыс наблюдается повышение уровня экспрессии мРНК GAP-43, при этом доля GAP-43-3 сохраняется на уровне контрольного значения. Это означает, что в клетках спинного мозга таких животных возрастает содержание целой молекулы GAP-43. У животных с тяжелым течением заболевания в клетках всех отделов спинного мозга, напротив, уровень GAP-43 уменьшается за счет снижения экспрессии мРНК GAP-43 и увеличения степени протеолитического расщепления GAP-43 кальпаином с образованием GAP-43-3.
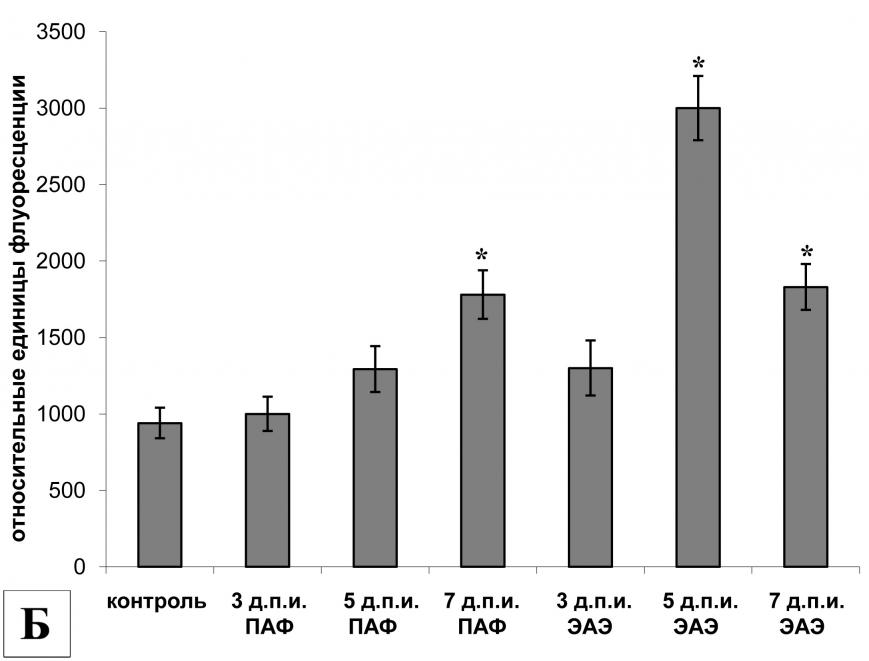
Белок GAP-43 в СМЖ крыс с различной тяжестью течения ЭАЭ
 | |
| Рис. 8. | Общее содержание белка GAP-43 и его протеолитических фрагментов в СМЖ крыс в процессе развития ЭАЭ при различной тяжести течения болезни (n=5-14). |
| * | - р<0,05 по сравнению с интактными животными. |
| ** | - р<0,001 по сравнению с интактными животными. |
Исследование проб ликвора, взятых до индукции ЭАЭ, показало наличие базового уровня белка GAP-43 в ликворе крыс, составившего в среднем 0,45 нг/мл. Общее содержание белка GAP-43 и его протеолитического фрагмента GAP-43-3 у животных без видимых неврологических нарушений в процессе развития ЭАЭ не изменялось и оставалось на базовом уровне на всех исследованных сроках. У заболевших животных выявлено изменение содержания белка GAP-43 и его протеолитического фрагмента в ликворе в процессе развития ЭАЭ. На 7-й д.п.и. в СМЖ всех впоследствии заболевших животных уровень белка GAP-43 практически не изменился по сравнению с базовым уровнем. В дальнейшем у заболевших животных наблюдалось увеличение общего количества белка GAP-43 и GAP-43-3 в СМЖ, причем у тяжело болевших животных подъем был более выражен, чем у легко болевших крыс. На пике заболевания (14-й д.п.и.) у животных с тяжелыми неврологическими нарушениями общее количество белка GAP-43 и GAP-43-3 в СМЖ составило 0,62 ± 0,02 нг/мл, т.е. уровень белка возрос в 1,4 раза по сравнению с базовым (рис. 8). У крыс с более легким течением заболевания содержание белка составило 0,57 ± 0,01 нг/мл. К 21-му д.п.и. у всех животных, имевших симптомы ЭАЭ на пике заболевания, видимых неврологических нарушений уже не отмечалось, однако в ликворе животных наблюдалось дальнейшее увеличение общего уровня белка GAP-43 и его протеолитического фрагмента. Ранее выявленная тенденция сохранялась: более высокий уровень белка GAP-43 в ликворе имели тяжело болевшие (на 14-й д.п.и.) животные по сравнению с легко болевшими – 0,93±0,03 нг/мл и 0,65±0,02 нг/мл, соответственно.
Общий уровень белка GAP-43 и GAP-43-3 в СМЖ крыс увеличивался при появлении клинических симптомов заболевания и продолжал возрастать в период выздоровления, достигая наибольшего значения при исчезновении видимых клинических проявлений. По-видимому, наличие высокого уровня белка GAP-43 и/или его протеолитического фрагмента GAP-43-3 в СМЖ крыс с ЭАЭ отражает течение двух противоположно направленных процессов: на пике заболевания – дегенерации клеток ЦНС и в период выздоровления – регенерации аксонов. Мы полагаем, что в результате гибели нейронов путем апоптоза или некроза в СМЖ попадает не только целая молекула белка GAP-43, но и его фрагмент GAP-43-3. Регенерация аксонов сопровождается регулируемым процессом секреции белка GAP-43 или GAP-43-3, возможно, опосредованным кальпаином. Действительно, при развитии нейродегенеративных процессов у крыс в СМЖ обнаруживаются некоторые внутриклеточные нейроспецифические белки (tau и спектрин) и их протеолитические фрагменты, являющиеся продуктами расщепления кальпаином (Siman et al., 2004).
Таким образом, уровень белка GAP-43 в СМЖ крыс с ЭАЭ отражает тяжесть и фазу течения болезни, что может служить диагностическим и/или прогностическим критерием развития аутоиммунного нейродегенеративного процесса.
Выводы
1. В индуктивную фазу ЭАЭ в иммуноцитах и клетках поясничного отдела спинного мозга крыс наблюдается дисбаланс экспрессии мРНК компонентов кальпаиновой системы; в клетках поясничного отдела спинного мозга выявлена гиперактивация кальпаинов.
2. Степень и распространенность сверхэкспрессии, а также гиперактивации кальпаинов клеток ЦНС является одним из факторов, определяющих тяжесть течения ЭАЭ у крыс; способность клеток ЦНС сверхэкспрессировать кальпастатин обеспечивает развитие легкой формы ЭАЭ.
3. Подавление активности кальпаинов с помощью ингибитора кальпаина I предотвращает развитие или снижает выраженность ЭАЭ у крыс, что указывает на возможность использования препаратов на основе ингибиторов кальпаина для лечения аутоиммунных нейродегенеративных заболеваний.
4. Бессимптомное развитие ЭАЭ сопровождается повышением уровня экспрессии мРНК GAP-43 в клетках спинного мозга; степень протеолиза белка GAP-43 сохраняется на уровне контрольного значения. У животных с тяжелым течением заболевания в клетках всех отделов спинного мозга уровень GAP-43 уменьшается за счет снижения экспрессии мРНК GAP-43 и увеличения степени протеолитического расщепления GAP-43 кальпаином.
5. Повышение уровня белка GAP-43 в СМЖ крыс с ЭАЭ отражает тяжесть и фазу течения болезни, что может служить диагностическим и/или прогностическим критерием развития аутоиммунного нейродегенеративного процесса.
Список работ, опубликованных по материалам диссертации.
1. Карпенко М.Н., Абдурасулова И.Н., Житнухин Ю.Л., Клименко В.М. Экспрессия мРНК кальпаинов в иммуноцитах и клетках ЦНС в индуктивную фазу экспериментального аллергического энцефаломиелита (ЭАЭ). Медицинский академический журнал, 2008, т.8, №4, с. 56-56.
2. Захаров В.В., Богданова М.Н., Мосевицкий М.И. Специфический протеолиз нейронального белка GAP-43 кальпаином. Характеристика, регуляция, физиологическая роль. Биохимия, 2005, т.70, с. 1086-1098.
3. Карпенко М.Н. Абдурасулова И.Н. Экспрессия мРНК и активность кальпаинов в клетках ЦНС крыс с различной тяжестью течения экспериментального аллергического энцефаломиелита. Материалы конференции Политехнического симпозиума, 9 декабря 2008 г, Санкт-Петербург, Сборник тезисов, с. 117.
4. Афанасьев П.В., Карпенко М.Н. Влияние ингибитора кальпаина на
выраженность экспериментального аллергического энцефаломиелита. XLVII неделя науки Санкт-Петербургского государственного политехнического университета (2008). Ч. IV: Материалы межвузовской научной конференции. СПб.: Изд-во СПбГПУ, стр. 4.
5. Карпенко М.Н. Абдурасулова И.Н. Клименко В.М. Экспрессия кальпаина в клетках различных отделов ЦНС и на периферии в процессе развития экспериментального аллергического энцефаломиелита. Объединенный иммунологический форум, Санкт-Петербург, 30 июня -5 июля 2008 г., Российский иммунологический Ж., 2008, т. 2(11), № 2-3, с. 216.
6. Карпенко М.Н., Абдурасулова И.Н., Клименко В.М. Изменение уровня экспрессии кальпаина в латентный период развития экспериментального аллергического энцефаломиелита у крыс. VI Сибирский Физиологический съезд, 25-27 июня 2008 г. Барнаул. Тезисы докладов. Том I, с. 236.
7. Карпенко М.Н., Абдурасулова И.Н., Клименко В.М. Изменение уровня экспрессии кальпаина в клетках различных отделов ЦНС и на периферии в процессе развития экспериментального аллергического энцефаломиелита. IV съезд Российского общества биохимиков и молекулярных биологов, 11-15 мая 2008 г, Новосибирск, Сборник тезисов, с. 468.
8. Karpenko M.N., Abdurasulova I.N., Klimenko V.M. Calpain expression in CNS and peripheral cells associated with experimental allergic encephalomyelitis. 11-th Multidisciplinary International Neuroscience and Biological Psychiatry Conference “Stress and behavior”, St-Petersburg, 2008, may 16-20, p. 64.
Marina N. Karpenko
Summary of the PhD-thesis “Calpain system involvement into autoimmune neurodegenerative processes (on the model of Experimental Autoimmune Encephalomyelitis)” (carried out at the Department of Physiology named after I.P. Pavlov, Research Institute for Experimental Medicine, St.-Petersburg, Russia)
Background. The calpain system is a system of cellular cysteine proteases that require calcium and are functionally active at neutral pH. Calpain activation takes place in two models: controlled activation under physiological conditions (only a few molecules of calpain are activated per cell), and hyperactivation under pathological conditions that involves sustained calcium overload (all available calpain molecules are activated). Regulated activation of calpain may be critical for function of many cells because of various structural proteins, enzymes and transcription factors are calpain substrate. Hyperactivation of calpain was revealed on the pathophysiology of many neurological disorders, including spinal cord injury, Parkinson’s disease, Alzheimer’s disease, brain ischemia, optic neuritis and on inflammatory diseases such as sepsis, pancreatitis etc. We believe that calcium influx mediates an increase of calpain activity that may cause inflammation and neurodegeneration in the spinal cord of rats with Experimental Autoimmune Encephalomyelitis (EAE) - an animal model of autoimmune neurodegenerative process. Thus, the aim of the investigation was to establish the calpain system involvement of immunocytes and CNS cells into autoimmune neurodegenerative processes on the model of Experimental Autoimmune Encephalomyelitis.
Methods. EAE induction and tissue sampling, RNA and protein extraction, RT-PCR, electrophoresis of nuclear acids and proteins in agarose and polyacrylamide gels, immunoblot analysis, calpain activity analysis, ELISA. Data are expressed as mean ± S.E.M. Statistical comparisons were made using one way ANOVA, Tukey’s post hoc test and Fisher’s test. Differences were considered significant at p<0.05.
Result. At the present study, the calpain expression at transcriptional level and calpain activity were evaluated in splenic, mesenteric lymph nodes inflammatory cells and spinal cord cells of Wistar rats with EAE before or after the onset of clinical symptoms. We observed an increased calpain expression in spleen and mesenteric lymph nodes of animals with EAE prior to symptomatology compared to normal control. The calpain activity of these cells did not differ from the control. Calpain expression was decreased up to normal control in spleen and mesenteric lymph nodes of animals with clinical symptoms (paralysis of tail and limbs). Calpain expression and activity was significantly increased prior to the onset of clinical symptoms in lumbar section of spinal cord. We observed an increase of m-calpain expression and activity in the cells of lumbar, thoracic and cervical spinal cord sections of rats with complete paralysis of forelimb on the peak of the disease. The increase of m-calpain expression and activity was not found out among the rats without signs of the disease. However the calpastatin expression increased.
The inhibition of enhanced calpain activity can be target for the treatment of autoimmune neurodegenerative process probably. We tested calpain inhibitor I as a potential therapeutic agent in autoimmune neurodegenerative processes as it readily cross the BBB. The prodrug was administered intranasaliy (treatment with a total of 1g calpain inhibitor I per rat a day from 1st to 16th day post immunization). This treatment demonstrated that the onset of EAE signs reduced. In this case the symptoms were limited to the flaccid tail and minimal hind limb weakness. Calpain inhibitor I also reduced the progression of the disease (p<0.009). The results showed calpain inhibitor I could be beneficial in the treatment of patients with neurodegenerative disorder of autoimmunity genesis.
Then we investigated the calpain capability to cleave GAP-43 (growth associated protein 43). The neuron specific protein GAP-43 is one of important proteins involved to basic nervous system specific processes such as neuroplasticity, regeneration and apoptosis. Proteolysis of GAP-43 by calpain brakes down the same properties. We observed an increase of GAP-43 proteolysis by calpain in the cells of lumbar, thoracic and cervical spinal cord sections of rats with complete paralysis of the forelimb on the peak of disease. We supposed that proteolysis of GAP-43 disrupts capability of axons to regeneration. An addition we detected an increase of GAP-43 level in cerebrospinal fluid (CSF) of rats with forelimb complete paralysis. Thus, the level of GAP-43 in CSF may be considered as a marker of autoimmune neurodegenerative processes development.