

Исследование ассоциации ряда генов-кандидатов с диабетической полинейропатией при сахарном диабете типа 1
На правах рукописи
СПИЦИНА ЕКАТЕРИНА ВАЛЕНТИНОВНА
ИССЛЕДОВАНИЕ АССОЦИАЦИИ РЯДА ГЕНОВ-КАНДИДАТОВ С ДИАБЕТИЧЕСКОЙ ПОЛИНЕЙРОПАТИЕЙ ПРИ САХАРНОМ ДИАБЕТЕ ТИПА 1
03.00.03 – молекулярная биология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Москва – 2008
Работа выполнена в лаборатории молекулярной диагностики и геномной дактилоскопии ФГУП «Государственный научно-исследовательский институт генетики и селекции промышленных микроорганизмов (ФГУП «ГосНИИ генетика»).
| |||||||||||||||||
Защита состоится «___» апреля 2008 г. в 14 часов на заседании Диссертационного совета Д.217.013.01 при Государственном научно-исследовательском институте генетики и селекции промышленных микроорганизмов по адресу: 117545, Москва, 1-й Дорожный проезд, 1.
С диссертацией можно ознакомиться в библиотеке ФГУП «ГосНИИ генетика».
Реферат разослан «___» марта 2008 г.
| Учёный секретарь Диссертационного совета, кандидат биологических наук | Г.Г. Заиграева |
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. В настоящее время сахарный диабет типа 1 относят к многофакторным, полигенным заболеваниям. Это заболевание является одной из основных причин ранней инвалидизации и смертности больных, что, в первую очередь обусловлено развитием его сосудистых осложнений. К последним относятся микроангиопатии – поражение капилляров, артериол и венул, клиническим проявлением которых является нейропатия, ретинопатия и нефропатия; и макроангиопатии – поражение сосудов крупного и среднего размера, которые приводят к инфаркту миокарда, инсульту и гангрене нижних конечностей.
Значение генетических факторов в развитии сахарного диабета и его осложнений в настоящее время общепризнанно. Актуальность исследований в области генетики СД типа 1 и его осложнений определяется, главным образом, тем, что эти исследования позволяют приблизиться к идентификации генов, предрасполагающих к развитию осложнений при СД типа 1, и пониманию того, какие именно гены и как определяют развитие этого заболевания и его осложнений, в частности, диабетической полинейропатии.
Диабетическая полинейропатия (ДПН) представляет собой серьезную медико-социальную проблему. Поражение нервной системы при сахарном диабете типа 1 сопровождается симптоматикой, у ряда больных значительно снижающей качество жизни. Диабетическую полинейропатию при СД типа 1 лишь условно можно считать его осложнением, так как функциональные нарушения периферической нервной системы наблюдаются уже в дебюте заболевания, что связано с высокой чувствительностью нервных клеток к повышенной концентрации глюкозы.
Генетическая предрасположенность к развитию ДПН при СД типа 1 связана с наследованием определенных аллелей обычных «здоровых» генов. Иногда эти аллели, которые определяют предрасположенность к развитию ДПН при СД типа 1 и сцеплены с заболеванием, называют этиологическими мутациями или вариантами. Часто этиологические варианты широко распространены в популяции, но при этом каждый из них сам по себе не приводит к развитию заболевания. Только наличие определенной комбинации этиологических вариантов в целом ряде генов, определяющих предрасположенность к заболеванию и его осложнений, может приводить к физиологическим нарушениям, находящим свое выражение в развитии ДПН при СД типа 1.
В связи с этим, большое значение отводится изучению генетической предрасположенности к развитию осложнений сахарного диабета типа 1 с использованием полиморфных маркеров различных генов-кандидатов, т.е. тех генов, чьи белковые продукты (регуляторные и структурные белки) могут быть потенциально вовлечены в развитие какого-либо заболевания. Для каждой многофакторной и полигенной патологии очерчивают определенный круг генов-кандидатов.
Исследование генов-кандидатов позволяет определить, существует ли вообще для данной патологии в конкретной популяции, предрасполагающие или предохраняющие генетические факторы (маркеры) и можно ли с помощью этих маркеров предсказать развитие болезни и ее осложнений задолго до появления симптомов, то есть прогнозировать течение заболевания.
Цель и задачи работы. Целью данной работы явилось изучение ассоциации с ДПН при СД типа 1 полиморфных маркеров нескольких генов-кандидатов. Эти гены кодируют белок р53 (TP53), NO-синтетазу клеток эндотелия (NOS3) сосудов, пероксидазу глутатиона 3 (GPX3), 3-субъединицу G-белка (GNB3), переносчик АТФ/АДФ типа 1 (ANT1), хеликазу мтДНК (PEO1), ДНК-полимеразу гамма 1 (POLG1) и поли(АДФ-рибозил)полимеразу (ADPRT1).
Предполагается, что продукты этих генов могут быть вовлечены в патогенез сосудистых осложнений диабета. В настоящей работе исследована ассоциация полиморфных маркеров данных генов с развитием диабетической полинейропатии при сахарном диабете типа 1 в русской популяции г. Москвы.
Для достижения этой цели были поставлены следующие задачи:
1. Определить частоты аллелей и генотипов полиморфных маркеров генов TP53, NOS3, GPX3, GNB3, ANT1, PEO1, POLG1 и ADPRT1 с диабетической полинейропатией при сахарном диабете типа 1 у русских пациентов, проживающих в г. Москве.
2. Провести сравнительный анализ распределения аллелей и генотипов полиморфных маркеров вышеуказанных генов–кандидатов в исследованной выборке для выявления ассоциации с развитием ДПН при СД типа 1 и определения вклада данных генов в наследственную предрасположенность к патологии.
Научная новизна работы. В данной работе впервые исследована ассоциация полиморфных маркеров C(–594)CC и Pro72Arg гена TP53, T(–786)C гена NOS3, G4077A гена GPX3, С825T гена GNB3, G(–25)A гена ANT1, G(–605)T гена PEO1, T(–365)C гена POLG1, Val762Ala и Leu54Phe гена ADPRT1 с диабетической полинейропатией при сахарном диабете типа 1 у русских пациентов, проживающих в г. Москве.
Практическая ценность работы. Выявление аллельных вариантов полиморфных маркеров различных генов-кандидатов, обуславливающих повышенный генетический риск развития ДПН при СД типа 1, создает базу для разработки диагностических методов прогнозирования течения заболевания.
Апробация работы. Диссертационная работа была представлена на заседании секции молекулярной биологии Ученого Совета ФГУП «ГосНИИ Генетика» 19 февраля 2008 г. Результаты настоящей работы докладывались на 41-ом ежегодном конгрессе европейской ассоциации по изучению диабета (г. Афины, Греция, 2005 г.), на 16-ой ежегодной научной конференции NEURODIAB (г. Юстад, Швеция, 2006 г.), на V Всероссийском конгрессе эндокринологов «Высокие медицинские технологии в эндокринологии» (г. Москва, Россия, 2006 г.), а также на научной конференции молодых ученых, посвященной 120-ти летию Н. И. Вавилова (г. Киев, Украина, 2007 г.).
Публикации. По материалам диссертации опубликовано 6 печатных работ, включая одну статью, а также тезисы докладов и сообщений на конференциях.
Структура диссертации. Диссертация состоит из следующих разделов: введение, обзор литературы, описание использованных материалов и методов, результаты и их обсуждение, а также выводы и список литературы. Материалы диссертации изложены на ___ страницах машинописного текста и содержат 13 таблиц и 10 рисунков.
СОДЕРЖАНИЕ РАБОТЫ
1. Формирование группы больных СД типа 1 с наличием и отсутствием диабетической полинейропатии.
Группы больных были сформированы из числа пациентов отделения эндокринологии Центральной клинической больницы Министерства путей сообщения Российской Федерации (ЦКБ МПС РФ) и обратившихся на амбулаторный прием на кафедру эндокринологии и диабетологии Российской медицинской академии последипломного образования (РМАПО), а также в ФГУ Эндокринологический научный центр (ЭНЦ) Росмедтехнологий.
Для проведения исследования использовали геномную ДНК 213 больных СД типа 1 с наличием («ДПН+», n = 100) и отсутствием («ДПН–», n = 113) диабетической полинейропатии. Группы пациентов формировали по принципу не перекрывающихся («полярных») фенотипов. В группу «ДПН+» вошли больные с длительностью диабета не менее 5 лет и наличием диабетической полинейропатии. Контрольную группу «ДПН–» составили пациенты, болеющие СД типа 1 на протяжении 10 и более лет без клинического диагноза ДПН. Характеристики групп больных представлены в таблице 1.
Геномную ДНК пациентов использовали для амплификации фрагментов ДНК, содержащих полиморфные маркеры ряда генов, предположительно вовлеченных в патогенез ДПН при СД типа 1.
Таблица 1.
Общая характеристика обследованных групп больных СД типа 1 с наличием («ДПН+») и отсутствием («ДПН–») диабетической полинейропатии
| Показатель (средний ± S.D.*) | «ДПН+» (n = 100) | «ДПН–» (n = 113) |
| Пол, м/ж | 61/39 | 43/70 |
| Возраст, лет* | 25,5 ± 14,5 | 27,5 ± 8,5 |
| Возраст начала СД типа 1, лет* | 22,4 ± 6,3 | 12,0 ± 3,0 |
| Длительность СД типа 1, лет* | 1,5 ± 1,4 | 15,5 ± 5,5 |
| Гликозилированный гемоглобин HbA1, %* | 6,3 ± 1,5 | 7,1 ± 1,7 |
*Среднее ± S. D. (стандартное отклонение)
Анализ нуклеотидных последовательностей интересующих нас хромосомных областей осуществляли с помощью системы NCBI в сети Интернет (www.ncbi.nlm.nih.gov), используя при этом следующие разделы: MapView (расположение этих полиморфных маркеров на хромосоме), dbSNP (информация об однонуклеотидных полиморфизмах). Для подбора праймеров и рестриктаз использовали пакеты программ DNAStar и VectorNTI 9.0.
Идентификация аллелей полиморфных маркеров проводилась с использованием полимеразной цепной реакции, дальнейшего расщепления фрагментов ДНК рестриктазами и электрофоретического разделения фрагментов ДНК в 8-12%-ном полиакриламидном геле или в 2-3% –ном агарозном геле.
2. Исследование ассоциации полиморфных маркеров C(–594)CC и Pro72Arg гена TP53 с ДПН при СД типа 1.
Белок р53 играет важную роль в регуляции транскрипции и поддержании геномной стабильности, взаимодействуя со многими клеточными белками. Показано, что повреждение ДНК способствует накоплению р53, который, в свою очередь, блокирует прогрессию клеточного цикла в фазе G1, препятствуя, таким образом, репликации ДНК до репарации повреждения. Если репарация повреждения невозможна, то белок р53 запускает механизм апоптоза (Kastan et al., 1991). При стрессах и повреждениях клеток активность и содержание р53 в них повышается. Также было показано, что активация р53 может происходить на фоне окислительного клеточного стресса, вызванного NO. Кроме того, активация p53 ведёт к изменению экспрессии генов некоторых секретируемых факторов, в результате может изменяться размножение и миграция не только поврежденной клетки, но и окружающих её клеток. При этом, находясь в стрессовой конформации, p53 в значительной степени утрачивает активности, стимулирующие рекомбинацию или репарацию ДНК (Копнин, 2001; Чумаков, 2000). Такой белок р53 значительно более стабилен (т.е. значительно увеличивается его количество в клетке) и эффективно активирует и/или репрессирует специфические гены-мишени, следствием чего является индукция в аномальных клетках либо остановки клеточного цикла, либо апоптоза.
Ген TP53 расположен на хромосоме 17q13.1. В этом гене и во фланкирующих его областях обнаружен ряд полиморфных участков, в том числе однонуклеотидный полиморфизм G/C, которому соответствует аминокислотный полиморфизм Pro/Arg (rs1042522) в положении 72 полипептидной цепи (Бухман и соавт., 1988; Ara et al., 1990) и полиморфизм, обусловленный встраиванием остатка С в положении –594 от точки инициации транскрипции (rs17881686).
Использование ряда экспериментальных моделей (животные и линии клеток) позволило установить, что белок р53 может быть вовлечен в патогенез диабетических осложнений. Так, на мышах линии NOD было показано, что гипергликемия приводит к апоптозу, опосредованному белком р53 (Adeghate, 2004; Keim et al., 2001). Кроме того, на экспериментальных моделях установлено, что апоптоз играет существенную роль в развитии диабетической ретинопатии (Barber et al., 1998) и нейропатии (Russell et al., 1999). Изучение ассоциации полиморфных маркеров Pro72Arg и C(–594)CC гена TP53 с ДПН при СД типа 1 на других популяциях ранее не проводилось. Проведенные на зарубежных популяциях исследования с использованием полиморфных маркеров гена ТР53 в основном посвящены изучению генетической предрасположенности к раку (Granja et al., 2004; Papadakis et al., 2002).
В нашей работе частоты аллелей полиморфного маркера С(–594)CС в группах «ДПН+» и «ДПН–» были следующими: аллель СС – 0,110 и 0,146, аллель С – 0,890 и 0,854 соответственно. Частоты генотипов: в группе «ДПН+»: СС/СС – 0,000, С/СС – 0,220, С/С – 0,780; в группе «ДПН–»: СС/СС – 0,000, С/СС – 0,292, С/С – 0,708 (табл. 2).
Этот полиморфный маркер расположен в промоторной зоне гена ТР53 и может, предположительно, влиять на регуляцию транскрипции. Однако различия в распределении частот аллелей и генотипов в сравниваемых в нашей работе группах не были статистически достоверными. Таким образом, полиморфный маркер С(–594)CС гена TР53 не ассоциирован с ДПН при СД типа 1 в русской популяции г. Москвы.
Частоты аллелей полиморфного маркера Pro72Arg в группах «ДПН+» и «ДПН–» составили: аллель Pro – 0,330 и 0,491, аллель Arg – 0,670 и 0,509. Известно, что в различных популяциях частота аллеля Pro составляет от 0,17 (среди саамов Швеции) до 0,63 (у негров Нигерии) (Beckman et al., 1994). В большом популяционном исследовании, проведенном в России, частота аллеля Pro составила 0,307 у белоруссов и 0,246 у русских из г. Смоленска (Khrunin et al., 2005). Наблюдаемые нами в группах больных СД типа 1 частоты аллелей данного маркера близки к полученным в этом популяционном исследовании.
Таблица 2.
Распределение частот аллелей и генотипов полиморфного маркера
С(–594)CС гена TP53 в группах больных сахарным диабетом (СД) типа 1 с наличием («ДПН+») и отсутствием («ДПН–») диабетической полинейропатии
| Аллель и генотип | Частота аллелей и генотипов | Уровень значимости, p | |
| «ДПН+» (n = 100) | «ДПН–» (n = 113) | ||
| Аллель С | 0,890 | 0,854 | >0,05 |
| Аллель СС | 0,110 | 0,146 | >0,05 |
| Генотип С/С | 0,780 | 0,708 | >0,05 |
| Генотип С/СС | 0,220 | 0,292 | >0,05 |
| Генотип СС/СС | 0,000 | 0,000 | >0,05 |
Распределились частоты генотипов следующим образом: в группе «ДПН+»: Pro/Pro – 0,160, Pro/Arg – 0,340, Arg/Arg – 0,500; в группе «ДПН–»: Pro/Pro – 0,301, Pro/Arg – 0,381, Arg/Arg – 0,319 (табл. 3).
Таблица 3.
Распределение частот аллелей и генотипов полиморфного маркера Pro72Arg гена TP53 в группах больных сахарным диабетом (СД) типа 1 с наличием («ДПН+») и отсутствием («ДПН–») диабетической полинейропатии
| Аллель и генотип | Частота аллелей и генотипов | Уровень значимости, p | OR | CI 95% | |
| «ДПН+» (n = 100) | «ДПН–» (n = 113) | ||||
| Аллель Arg | 0,670 | 0,509 | 0,0008 | 1,96 | 1,32 – 2,90 |
| Аллель Pro | 0,330 | 0,491 | 0,51 | 0,34 – 0,76 | |
| Генотип Arg/Arg | 0,500 | 0,319 | 0,0080 | 2,14 | 1,23 – 3,73 |
| Генотип Arg/Pro | 0,340 | 0,381 | 0,5698 | 0,84 | 0,48 – 1,47 |
| Генотип Pro/Pro | 0,160 | 0,301 | 0,0228 | 0,44 | 0,23 – 0,86 |
Сравнительный анализ выявил достоверные различия частот аллелей и генотипов данного полиморфного маркера в группах «ДПН+» и «ДПН–». При этом риск развития патологии оказался связан с носительством аллеля Arg (OR = 1,96; CI = 1,32 – 2,90) и генотипа Arg/Arg (OR = 2,14; CI = 1,23 – 3,73). Аллель Pro, напротив, ассоциирован с пониженным риском развития ДПН (OR = 0,51; CI = 0,34 – 0,76), так же как и генотип Pro/Pro (OR = 0,44; CI = 0,23 – 0,86).
Белок р53 содержит несколько функционально значимых доменов (Ko & Prives, 1996). Полиморфный маркер Pro72Arg расположен в домене, обогащенном остатками пролина, который образован аминокислотными остатками 61 – 94. Этот домен содержит повторы РХХР, где Р – остаток пролина, а X – любой аминокислотный остаток (Zhu et al., 1999). Домен, богатый пролином, по-видимому, участвует в регуляции транскрипции ряда генов, вовлеченных в индукцию апоптоза. Так, богатый пролином домен белка р53 (Venot et al., 1998) вовлечен в регуляцию транскрипции гена PIG3, белковый продукт которого гомологичен ряду оксидоредуктаз и участвует, предположительно, в апоптозе, зависимом от р53. Делеции в этом домене влияют на транскрипцию гена Р85 (Yin et al., 1998), белковый продукт которого, регуляторная субъединица киназы PI3K, вовлечен в апоптоз при окислительном стрессе, зависимом от р53 (Walker & Levine, 1996). Домен, богатый пролином, необходим также для взаимодействия р53 с киназой Chk2 в ответ на повреждение ДНК (Berger et al., 2005).
В зависимости от того, какой аминокислотный остаток находится в положении 72 полипептидной цепи белка р53, меняется способность р53 запускать апоптоз (Thomas et al., 1999). В частности, белок р53 с остатком Arg в положении 72 более эффективно вызывал индукцию апоптоза, и его количество в митохондриальной фракции превышало почти на порядок количество белка р53, содержащего остаток Pro (Dumont et al., 2003).
Результаты нескольких популяционных исследований ассоциации полиморфного маркера Pro72Arg гена TP53 с рядом заболеваний нервной системы противоречивы. Так, в китайской популяции выявили ассоциацию аллеля Pro этого полиморфного маркера с развитием нейропатии при первичной открытоугольной глаукоме (Lin et al., 2002). Наследственная нейропатия зрительного нерва Лебера раньше развивалась у японцев, гомозиготных по аллелю Arg (Ishikawa et al., 2005), т. е. генотип Arg/Arg так же, как и в нашей работе, служил фактором риска развития нейропатии. В то же время, в одной из недавних работ, проведенной в Кувейте (Alkhalaf et al., 2007), обнаружено отсутствие ассоциации этого маркера с диабетом и ишемической болезнью сердца (эти заболевания имеют много общих факторов риска, связанных с метаболическими нарушениями).
Полученные в нашей работе результаты позволяют говорить об ассоциации полиморфного маркера Pro72Arg гена TP53 с ДПН у больных СД типа 1 русских жителей г. Москвы. Ассоциация этого полиморфного маркера с ДПН свидетельствует о важной роли белка р53 в патогенезе данного осложнения СД.
Представлялось интересным сравнить нам данные с результатами аналогичных работ, полученными на других популяциях. Однако наша работа представляет собой первую попытку оценить вклад гена ТР53 в развитие ДПН при СД типа 1 в русской популяции г. Москвы.
3. Исследование ассоциации полиморфного маркера T(–786)C гена NOS3 с ДПН при СД типа 1.
Ген NOS3 расположен на хромосоме 7 в области q36 (Marsden et al., 1993; Robinson et al., 1994) и состоит из 26 экзонов. В экзонах и интронах этого гена был обнаружен ряд полиморфных участков, из которых в большинстве исследований использовали два: минисателлит в интроне 4 (ecNOS4a/4b), состоящий из 4 (аллель 4a) или 5 (аллель 4b) повторяющихся единиц длиной 27 п.н. (Miyahara et al., 1994), и однонуклеотидный полиморфизм (G/T) в положении 894 в экзоне 7, которому соответствует аминокислотный полиморфизм в положении 298 (Glu298Asp) (Shimasaki et al., 1998). Кроме того, в единичных работах использовался полиморфный маркер Т(–786)С, расположенный в области промотора в положении –786.
В одной из работ было показано, что полиморфизм T(–786)C гена NOS3 ассоциирован с развитием ранних проявлений заболевания коронарных артерий (Alvarez et al., 2001). Исследователи предположили, что более всего подвержены ранним проявлениям этого заболевания носители генотипа CC, вероятно, из-за нарушения функции эндотелия. В этой же работе было показано совместное влияние генотипов NOS3-CC и ACE-DD (фермент, превращающий ангиотензин I) на риск развития ранних проявлений заболевания коронарных артерий.
На русской популяции г. Москвы была показана ассоциация полиморфного маркера Т(–786) гена NOS3 с развитием диабетической нефропатии при СД типа 1. Риск развития диабетической нефропатии в этом случае был существенно выше у носителей генотипа СС, по сравнению с носителями генотипов ТС и ТТ (Носиков, 2004).
В японской популяции была обнаружена ассоциация полиморфного маркера ecNOS4a/4b гена NOS3 с диабетической нейропатией у больных СД типа 2. У больных с ДПН содержание аллеля 4а и генотипа 4а/4а и 4а/4b связано с повышенным риском развитием данной патологии, в то время как, носительство аллелям 4b и гомозигот 4b/4b является защитным фактором (Tanaka et al., 1998).
В нескольких работах было показано, что полиморфизм ecNOS4a/4b ассоциирован с уровнем нитросоединений (нитратов и нитритов) в крови, напрямую связанным со скоростью выработки NO эндотелием сосудов. У австралийцев и японцев, гомозиготных по аллелю 4a, содержание нитросоединений было понижено (Wang et al., 1997; Tsukada et al., 1998). В условиях окислительного стресса под действием перекисей и свободных радикалов кислорода резко падает содержание окиси азота в плазме, что способствует развитию патологических изменений в сосудах и прогрессированию атеросклероза.
Таким образом, с высокой степенью вероятности можно сделать вывод о том, что ген NO-синтетазы клеток эндотелия (NOS3) вовлечен в развитие не только сердечно-сосудистых патологий, но и связан с генетической предрасположенностью к сосудистым осложнениям при СД типа 1 и 2.
В нашей работе аллели полиморфного маркера Т(–786)С выявляли после расщепления амплифицированного фрагмента длиной 273 п.н., содержащего полиморфный участок, рестриктазой MroNI. Фрагмент ДНК, содержащий аллель Т остается нерасщепленным, в то время как фрагмент ДНК, содержащий аллель С расщепляется на фрагменты размером 172 и 101 п.н. (рис. 1).
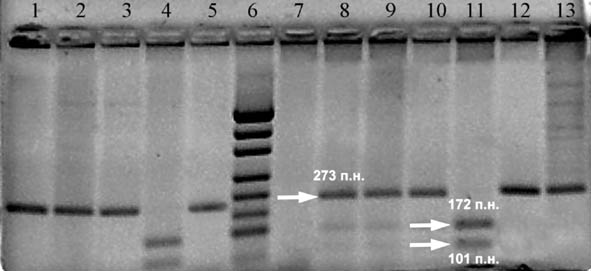
Рисунок 1. Электрофоретическое разделение в 2% агарозном геле продуктов расщепления амплифицированного фрагмента ДНК, содержащего полиморфный маркер Т(–786)С гена NOS3. Генотипы по дорожкам: 1-3, 5, 12, 13 – ТТ; 4, 11 – СС; 8-10 – ТС; 6 – pUC19/MspI.
При исследовании распределения аллелей и генотипов полиморфного маркера Т(–786)С гена NOS3 среди больных СД типа 1 с наличием и отсутствием ДПН обнаружено преобладание частоты аллеля Т над частотой аллеля С, и встречаемости генотипа СС над встречаемостью генотипов ТТ и ТС в обеих исследованных группах (табл. 4).
Таблица 4.
Распределение частот аллелей и генотипов полиморфного маркера Т(–786)С гена NOS3 в группах больных СД типа 1 с наличием (группа «ДПН+») и отсутствием (группа «ДПН–») диабетической полинейропатии
| Аллель и генотип | Частота аллелей и генотипов | Уровень значимости, p | |
| «ДПН+» (n = 100) | «ДПН–» (n = 113) | ||
| Аллель T | 0,520 | 0,527 | >0,05 |
| Аллель C | 0,480 | 0,473 | >0,05 |
| Генотип TT | 0,260 | 0,257 | >0,05 |
| Генотип CC | 0,520 | 0,540 | >0,05 |
| Генотип TC | 0,220 | 0,204 | >0,05 |
Однако различия в распределении аллелей и генотипов в обеих группах были незначительными и носили недостоверный характер. Таким образом, полиморфный маркер Т(–786)С гена NOS3 не ассоциирован с ДПН при СД типа 1 в русской популяции г. Москвы.
Решение о проведении исследования по поиску ассоциации полиморфного маркера T(–786)C гена NOS3 у больных СД типа 1 с ДПН, было принято на основании того, что в нашей лаборатории была обнаружена ассоциация этого полиморфного маркера с диабетической нефропатией при СД типа 1. Причем, риск развития диабетической нефропатии был существенно выше у носителей генотипа СС по сравнению с носителями генотипов ТС и ТТ (Носиков, 2004).
4. Исследование ассоциации полиморфного маркера G4077A гена GPX3 с ДПН при СД типа 1.
Пероксидазы глутатиона (GPX) – важнейшие ферменты, обеспечивающие инактивацию активных форм кислорода, так как они разрушают перекись водорода и гидроперекиси липидов. Хотя эти ферменты обнаруживаются повсеместно, уровень каждой изоформы зависит от типа ткани.
В течение длительного времени идет спор относительно первоначальной роли этих ферментов. В то время, как некоторые авторы придерживаются мнения, что эти ферменты играют главную роль в защите против окислительных нарушений при нормальном физиологическом состоянии (Jones et al., 1981; Makino et al., 1994), другие считают, что защитная роль этих ферментов проявляется только в состоянии окислительного стресса (Giri et al., 1983; Kelner et al., 1990).
Ген GPX3 расположен в области 5q32-q33.1, и состоит из 5 экзонов и 4 интронов (Chu, 1994) В нашей работе мы исследовали полиморфный участок G4077A, расположенный в 1 интроне этого гена. Полиморфный маркер G4077A гена GPX3 представляет собой однонуклеотидный полиморфизм: остатки гуанина или аденина в положении 4077 нуклеотидной последовательности, не приводящие к аминокислотному полиморфизму.
Считается, что пероксидаза глутатиона 3 (GPX3) присутствует в основном в плазме и действует, как антиокислитель, защищая клеточные мембраны. Фермент GPX3 инактивирует перекиси фосфолипидов, таким образом напрямую участвуя в защите мембран. Хотя GPX3 синтезируется в разных тканях, но основной источник этого фермента находится в почках.
В эксперименте на 3-х группах нокаутных мышей: без сахарного диабета, только заболевших сахарным диабетом и с длительным течением сахарного диабета, было показано, что, по сравнению с контролем, ген GPX3 больше всего экспрессировался у мышей с ранними проявлениями сахарного диабета. Меньше всего ген GPX3 экспрессировался у мышей, длительно болеющих сахарным диабетом (Wilson et al., 2003).
Известно, что окислительный стресс играет немаловажную роль в развитии диабетических микроангиопатий. Поэтому, было интересно изучить ассоциацию полиморфного маркера G4077A гена GPX3 с развитием диабетической полинейропатии при СД типа 1.
В нашей работе в группах «ДПН+» и «ДПН–» частота аллеля А преобладала над частотой аллеля G, а встречаемость гомозигот АА – над встречаемостью генотипов GG и GA (табл. 5).
При сравнении распределения аллелей и генотипов полиморфного маркера G4077A гена GPX3 в группах больных СД типа 1 с наличием и отсутствием диабетической полинейропатии, не было обнаружено никаких статистически достоверных различий. Это свидетельствует об отсутствии ассоциации между данным полиморфным маркером и ДПН при СД типа 1 в исследованной нами выборке. Литературные данные об исследовании ассоциации этого полиморфного маркера с развитием ДПН при СД типа 1 в других популяциях отсутствуют.
Таблица 5.
Распределение частот аллелей и генотипов полиморфного маркера G4077А гена GPX3 в группах больных СД типа 1 с наличием («ДПН+») и отсутствием («ДПН–») диабетической полинейропатии
| Аллель и генотип | Частота аллелей и генотипов | Уровень значимости, p | |
| «ДПН+» (n = 100) | «ДПН–» (n = 113) | ||
| Аллель G | 0,410 | 0,425 | >0,05 |
| Аллель A | 0,590 | 0,575 | >0,05 |
| Генотип GG | 0,100 | 0,097 | >0,05 |
| Генотип AA | 0,620 | 0,655 | >0,05 |
| Генотип GA | 0,280 | 0,248 | >0,05 |
5. Исследование ассоциации полиморфного маркера С825Т гена GNB3 с ДПН при СД типа 1.
G-белки – гетеротримеры, в которых -субъединица непрочно связана с димером. Все известные -субъединицы (мол. масса 40-50 кДа), гомологичны, и у большинства из них одинаковые (или очень сходные) -субъединицы (мол. масса 35 кДа) и -субъединицы (мол. масса 8 кДа).
Существует 5 подтипов -субъединиц и 12 подтипов -субъединиц, однако не все возможные комбинации этих субъединиц существуют в природе. У каждого G-белка может быть несколько мишеней.
-субъединицы обеспечивают локализацию, эффективное связывание и инактивацию -субъединиц, регулируют сродство рецепторов к их активирующим лигандам, понижают способность GDP (гуанозиндифосфат) к диссоциации от -субъединицы (стабилизация инактивированного состояния), открывают К+-канал в сердце, закрывают Са2+-канал в пресинаптической мембране, активируют фосфолипазу А2, и некоторые изоформы фосфолипазы С, регулируют сродство рецептора к агонисту.
Ген GNB3 локализован на хромосоме 12 в области р12, и состоит из 11 экзонов и 10 интронов. Мы исследовали ассоциацию однонуклеотидного полиморфизма С/Т в экзоне 10 (положение 825 мРНК) (Rosskopf et al., 2000).
Замена остатка С на Т в положении 825 в гене GNB3, который кодирует 3-субъединицу G-белка, сопровождается альтернативным сплайсингом, что приводит к укороченному варианту 9-го экзона. Следствием этого является утрата одного белкового домена. Молекулярные механизмы, вовлеченные в альтернативный сплайсинг, еще полностью не исследованы.
Так, было установлено, что высокая частота аллеля 825Т гена GNB3 у «древних» народностей, например у бушменов, австралийских аборигенов, а также в популяции народов с темной кожей, выраженно ассоциирована с ожирением. Также выяснилось, что этот аллель ассоциирован с низкой активностью ренина при артериальной гипертензии и предрасполагает к развитию гипертрофии левого желудочка (ГЛЖ). При СД типа 2 аллель 825Т предрасполагает к конечной стадии заболеваний почек. Но для пациентов с СД типа 1 такой ассоциации найдено не было (Siffert, 2000).
За последние несколько лет было показано, что в действии инсулина участвуют механизмы, связанные с G-белками. Инсулин меняет чувствительность к агентам, которые действуют посредством G-белков и обратно. Было показано, что у носителей аллеля 825T чувствительность к инсулину повышена (Fernndez-Real et al., 2003).
Таким образом, ген 3-субъединицы G-белка может являться одним из генов, определяющих генетическую предрасположенность к развитию ДПН у больных сахарным диабетом типа 1. Однако данные по ассоциации полиморфных маркеров гена GNB3 с ДПН при СД типа 1 отсутствуют, как для российских, так и для зарубежных популяций.
В группах «ДПН+» и «ДПН–» частота аллеля С значительно преобладала над частотой аллеля Т, а частота гомозиготного генотипа СС – над частотой генотипов ТТ и СТ (табл. 6).
Таблица 6.
Распределение частот аллелей и генотипов полиморфного маркера С825Т гена GNB3 в группах больных СД типа 1 с наличием («ДПН+») и отсутствием («ДПН–») диабетической полинейропатии
| Аллель и генотип | Частота аллелей и генотипов | Уровень значимости, p | OR | CI 95% | |
| «ДПН+» (n = 100) | «ДПН–» (n = 113) | ||||
| Аллель C | 0,620 | 0,779 | 0,0004 | 0,46 | 0,30 – 0,71 |
| Аллель T | 0,380 | 0,221 | 2,16 | 1,41 – 3,30 | |
| Генотип CC | 0,580 | 0,637 | 0,402 | 0,79 | 0,45 – 1,37 |
| Генотип CT | 0,340 | 0,080 | 0,0002 | 5,95 | 2,68 – 13,21 |
| Генотип TT | 0,080 | 0,283 | 0,0002 | 0,22 | 1,10 – 0,50 |
Достоверное увеличение частоты аллеля С (OR = 0,46; p = 0,0004) и генотипа СС (OR = 0,79; p = 0,402) в группе говорит о повышенном риске развития ДПН при СД типа 1. Таким образом, можно сделать вывод об ассоциации полиморфного маркера С825Т гена GNB3 с ДПН при СД типа 1.
6. Исследование ассоциации полиморфного маркера G(–25)A гена ANT1 с ДПН при СД типа 1.
Известно, что митохондрии принимают участие в программируемой гибели клеток (апоптоз), и, что значимую роль в этом процессе играет образование митохондриальной поры (мегаканала) (Kristal et al., 1996), которая в том числе может открываться и в ответ на окислительный стресс (Halestrap et al., 2002). В образовании митохондриальной поры как раз и принимают участие переносчики АТФ/АДФ (Vyssokikh & Brdiczka, 2003). Согласно одной из работ, в митохондриях при диабете происходит ряд метаболических изменений, результатом которых является положительная корреляция между тяжестью сахарного диабета (и/или длительностью заболевания) и степенью снижения чувствительности мембран митохондрий к открытию мегаканала (Ferreira et al., 2003).
Существует несколько переносчиков АТФ/АДФ: АNT1, АNT2, АNT3 и АNT4. Каждый из них кодируется своим геном (Cozens et al., 1989), причем экспрессия некоторых из этих генов тканеспецифична (Doerner et al., 1997). Так, АNT1 преимущественно экспрессируется в сердце, мышцах и мозге, АNT2 – в печени, почках и селезенке (Doerner et al., 1997), АNT4 – в тестикулах (Brower et al., 2007), а уровень экспрессии АNT3 примерно одинаков во всех тканях (Doerner et al., 1997).
Переносчик АТФ/АДФ типа 1 (ANT1) представляет собой гомодимер, образованный двумя субьединицами. Его основная функция – это транспорт АТФ и АДФ через внутреннюю мембрану митохондрий (Klingenberg, 1985). Переносчики АТФ/АДФ также способны взаимодействовать с факторами транскрипции (Bottero et al., 2001) и с белками, вовлеченными в регуляцию процесса апоптоза (Belzacq et al., 2002), такими как bcl-2 и bax.
Ген ANT1 расположен на хромосоме 4 в области 4q35. В митохондриях мышей, нокаутных по этому гену, наблюдался повышенный уровень активных форм кислорода и, как следствие, повреждение митохондриальной ДНК (Graham et al., 1997).
Ассоциация полиморфного маркера G(–25)A гена ANT1 (rs3733652) с осложнениями сахарного диабета до настоящего времени не изучалась. Большинство исследований ассоциации гена ANT1 с заболеваниями проводилось на больных с аутосомно-доминантной прогрессирующей наружной офтальмоплегией, заболеванием, при котором поражаются черепные нервы центрального генеза или периферического – ретробульбарного. У больных с этой патологией обнаружены мутации A114P и D104G гена ANT1 (Lodi et al., 2006), а также мутация R334Q (Van Goethem et al., 2003).
В данной работе мы исследовали ассоциацию полиморфного маркера
G (–25)A гена ANT1, исходя из того, что ген ANT1 может определять наследственную предрасположенность к развитию ДПН при СД типа 1.
В группах пациентов с СД типа 1 с диабетической полинейропатией («ДПН+») и без этого осложнения («ДПН–») были определены частоты аллелей и генотипов полиморфного маркера G(–25)A гена ANT1. Частоты аллелей в группах «ДПН+» и «ДПН–» составили: аллель G – 0,700 и 0,708, аллель А – 0,300 и 0,292, соответственно. Распределение частот генотипов оказалось следующим: в группе «ДПН+»: GG – 0,550, GA – 0,300, AA – 0,150; в группе «ДПН–»: GG – 0,593, GA – 0,230, AA – 0,177 (табл. 7).
Таблица 7.
Распределение частот аллелей и генотипов полиморфного маркера G(–25)A гена ANT1 в группах больных сахарным диабетом (СД) типа 1 с наличием («ДПН+») и отсутствием («ДПН–») диабетической полинейропатии
| Аллель и генотип | Частота аллелей и генотипов | Уровень значимости, p | |
| «ДПН+» (n = 100) | «ДПН–» (n = 113) | ||
| Аллель G | 0,700 | 0,708 | > 0,05 |
| Аллель A | 0,300 | 0,292 | > 0,05 |
| Генотип GG | 0,550 | 0,593 | > 0,05 |
| Генотип GA | 0,300 | 0,230 | > 0,05 |
| Генотип AA | 0,150 | 0,177 | > 0,05 |
Различия в распределении аллелей и генотипов между группами были незначительными и носили недостоверный характер. Это свидетельствует об отсутствии ассоциации полиморфного маркера G(–25)A гена ANT1 и ДПН при СД типа 1 в русской популяции г. Москвы. Впрочем, нельзя исключить, что данный полиморфный маркер гена ANT1 не имеет функционального значения и не сцеплен с другим функционально важным полиморфным маркером.
7. Исследование ассоциации полиморфного маркера G(–605)Т гена PEO1 с ДПН при СД типа 1.
Ген PEO1 расположен на 10 хромосоме в области 10q23.3-24.3. Этот ген кодирует хеликазу митохондриальной (мтДНК). В условиях in vitro было показано, что данный белок обладает хеликазной активностью (Korhonen et al., 2003). Наряду с ДНК-полимеразой гамма 1 (POLG1), хеликаза участвует в репликации мтДНК, что было показано в условиях in vitro (Korhonen et al., 2004), при этом повышение уровня ДНК-полимеразы гамма не приводит к увеличению числа копий мтДНК (Spelbrink et al., 2000), а в результате повышенной экспрессии хеликазы мтДНК наблюдается трехкратное возрастание количества мтДНК (Tyynismaa et al., 2004).
Основные работы по исследованию ассоциации гена PEO1 с заболеваниями проводились на пациентах с аутосомно-доминантной прогрессирующей наружной офтальмоплегией, также как и в случае гена ANT1. В этих работах было обнаружено несколько мутаций гена PEO1 – A359T, A475P (Spelbrink et al., 2001) и мутация R374Q (Baloh et al., 2007).
Для данного исследования нами был выбран полиморфный маркер G(–605)Т гена PEO1 (rs3740484), расположенный в промоторной области, так как он, предположительно, может влиять на уровень экспрессии гена. Ассоциация полиморфного маркера G(–605)T гена PEO1 с осложнениями сахарного диабета до настоящего времени не изучалась. В одной из недавних работ было показано, что снижение уровня хеликазы мтДНК приводит к уменьшению числа копий мтДНК. Эти результаты были получены на культуре клеток и подтверждены в условиях in vivo (Tyynismaa et al., 2004).
В группах пациентов с СД типа 1 с диабетической полинейропатией («ДПН+») и без этого осложнения («ДПН–») были определены частоты аллелей и генотипов полиморфного маркера G(–605)Т гена PEO1 (табл. 8).
Таблица 8.
Распределение частот аллелей и генотипов полиморфного маркера G(–605)Т гена PEO1 в группах больных сахарным диабетом (СД) типа 1 с наличием («ДПН+») и отсутствием («ДПН–») диабетической полинейропатии
| Аллель и генотип | Частота аллелей и генотипов | Уровень значимости, p | |
| «ДПН+» (n = 100) | «ДПН–» (n = 113) | ||
| Аллель T | 0,485 | 0,429 | > 0,05 |
| Аллель G | 0,515 | 0,571 | > 0,05 |
| Генотип TT | 0,260 | 0,221 | > 0,05 |
| Генотип TG | 0,450 | 0,416 | > 0,05 |
| Генотип GG | 0,290 | 0,363 | > 0,05 |
Частоты аллелей в группах «ДПН+» и «ДПН–» составили: аллель Т – 0,485 и 0,429, аллель G – 0,515 и 0,571, соответственно. Распределение частот генотипов оказалось следующим: в группе «ДПН+»: ТТ – 0,260, TG – 0,450, GG – 0,290; в группе «ДПН–»: ТТ – 0,221, TG – 0,416, GG – 0,363. Однако различия в распределении аллелей и генотипов в обеих исследованных группах были незначительными и носили недостоверный характер.
Таким образом, полученные нами результаты свидетельствуют об отсутствии ассоциации полиморфного маркера G(–605)T гена PEO1 с развитием ДПН при СД типа 1 в русской популяции г. Москвы.
8. Исследование ассоциации полиморфного маркера T(–365)C гена POLG1 с ДПН при СД типа 1.
Известно, что мутации в гене, кодирующем ДНК-полимеразу гамма 1 и в гене, кодирующем хеликазу мтДНК, могут проявляться весьма сходно фенотипически (Falkenberg et al., 2007). Продукты этих двух генов участвуют в репликации мтДНК. Кроме того, ДНК-полимераза гамма участвует также и в ее репарации (Hudson & Chinnery, 2006; Zheng et al., 2006; Graziewicz et al., 2006). Это позволило предположить, что ген POLG1 также может быть связан с развитием данного осложнения. Показано, что для этого фермента характерна довольно высокая частота ошибок при репликации, что приводит к появлению мутаций (Zheng et al., 2006).
ДНК-полимераза гамма – это гетеродимер, состоящий из трех субъединиц. Одна из них, каталитическая, обладает ДНК-полимеразной и 3'-5'-экзонуклеазной активностями. Две идентичные вспомогательные субъединицы взаимодействуют с ДНК (Lee et al., 2007). Вспомогательные субъединицы кодирует ген POLG2, расположеный на хромосоме 17q (Longley et al., 1998), а каталитическую – ген POLG1, находящийся на хромосоме 15q25 (Kaguni et al., 2004).
Как в самом гене POLG1, так и в прилегающих к нему участках обнаружено более ста полиморфных маркеров (Longley et al., 2005), в том числе полиморфизм T(–365)C (rs2856268), расположенный в области, предшествующей гену. Два других полиморфных маркера (A/G, rs758130) и (T/C, rs2238296), расположенные в интроне 1 гена POLG1, были использованы в проведенном недавно популяционном исследовании у народов, проживающих в Сибири (Буйкин и соавт., 2006).
Показано, что ряд мутаций гена POLG1 является причиной развития ряда заболеваний, в частности, таких заболеваний нервной системы, как паркинсонизм (Luoma et al., 2004) и синдром Альперса (Ferrari et al., 2005). Также было показано, что при ДПН имеет место апоптоз нейронов (Vincent et al., 2002) и, что для диабетической нейропатии характерно повреждение митохондрий (Honma et al., 2004).
Известно, что для диабетической гипергликемии характерен окислительный стресс, который может приводить к повреждению ДНК. Он гораздо более выражен в митохондриях, поэтому мтДНК повреждается существенно быстрее, нежели ядерная ДНК. Это приводит к снижению в митохондриях синтеза мРНК, кодирующих белки дыхательной цепи (DiMauro & Schon, 2003) и последующему снижению уровня синтеза АТФ (van Houten et al., 2006).
Таким образом, кодирующий каталитическую субъединицу ДНК-полимеразы гамма 1 ген POLG1 может являться одним из генов, определяющих генетическую предрасположенность к развитию ДПН у больных сахарным диабетом типа 1. Однако данные по ассоциации полиморфных маркеров гена POLG1 с ДПН при СД типа 1 отсутствуют, как для российских, так и для зарубежных популяций. Нами был выбран полиморфный маркер T(–365)C, так как он расположен в промоторной области гена POLG1 и может влиять на регуляцию его транскрипции.
В проведенных ранее исследованиях в основном использовали ряд мутаций, расположенных непосредственно в гене POLG1, причем наиболее хорошо были изучены те из них, которым соответствуют аминокислотные замены в районе полимеразного либо экзонуклеазного доменов. Одной из наиболее изученных мутаций G1399A соответствует аминокислотная замена Ala467Thr, расположенная между доменом с полимеразной активностью и экзонуклеазным доменом. Наличие этой мутации определяет развитие ряда заболеваний, в частности, синдрома Альперса (Nguyen et al., 2005). Другой мутацией, определяющей развитие этого синдрома, является мутация Glu873Stop (Naviaux et al., 2004). Также было показано, что одной из причин развития нейропатии (атаксии) является носительство мутации Trp748Ser (Nguyen et al., 2005).
Сравнительный анализ выявил достоверные различия частот аллелей и генотипов данного полиморфного маркера в группах «ДПН+» и «ДПН–». Частоты аллелей полиморфного маркера T(–365)C в группах «ДПН+» и «ДПН–» в нашей работе были следующими: аллель С – 0,545 и 0,425, аллель Т – 0,455 и 0,575 соответственно. Частоты генотипов: в группе «ДПН+»: СС – 0,400, СТ – 0,290, ТТ – 0,310; в группе «ДПН–»: СС – 0,274, СТ – 0,301, ТТ – 0,425. При этом риск развития патологии оказался связан с носительством аллеля С (OR = 1,62; CI = 1,11 – 2,38) и генотипа СС (OR = 1,76; CI = 0,99 – 3,13). Аллель Т, напротив, ассоциирован с пониженным риском развития ДПН при СД типа 1 (OR = 0,62; CI = 0,42 – 0,90), так же как и генотип ТТ (OR = 0,61; CI = 0,35 – 1,07) (табл. 9).
Полученные нами результаты позволяют говорить об ассоциации полиморфного маркера T(–365)C гена POLG1 с ДПН при СД типа 1 у русских пациентов, проживающих в г. Москве. Это может указывать на то, что вклад в развитие ДПН при СД типа 1 вносит нарушение репарации мтДНК, повреждение которой происходит в результате характерного для диабета окислительного стресса.
Таблица 9.
Распределение частот аллелей и генотипов полиморфного маркера Т(–365)С гена POLG1 в группах больных сахарным диабетом (СД) типа 1 с наличием («ДПН+») и отсутствием («ДПН–») диабетической полинейропатии
| Аллель и генотип | Частота аллелей и генотипов | Уровень значимости, p | OR | CI 95% | |
| «ДПН+» (n = 100) | «ДПН–» (n = 113) | ||||
| Аллель C | 0,545 | 0,425 | 0,0152 | 1,62 | 1,11 – 2,38 |
| Аллель T | 0,455 | 0,575 | 0,62 | 0,42 – 0,90 | |
| Генотип CC | 0,400 | 0,274 | >0,05 | 1,76 | 0,99 – 3,13 |
| Генотип CT | 0,290 | 0,301 | >0,05 | 0,95 | 0,53 – 1,71 |
| Генотип TT | 0,310 | 0,425 | >0,05 | 0,61 | 0,35 – 1,07 |
9. Исследование ассоциации полиморфных маркеров Val762Ala и Leu54Phe гена ADPRT1 с ДПН при СД типа 1.
Повреждения ДНК, обусловленные генотоксичным действием свободных радикалов, приводят к активации поли(ADP-рибозил)полимераз (PARP). Эти ферменты участвуют в репарации ДНК, катализируя поли(АДФ-рибозилирование) белков, связанных с ДНК (Новожилова, 1996; Oliver et al., 1999). Активность PARP возрастает в 500 раз и более при связывании с участками разрыва ДНК (Xiao et al., 2004)
Известно несколько видов белков PARP, кодируемых разными генами. Из них ассоциации с патогенезом ДПН при СД типа 1, прежде всего следует ожидать от PARP1, так как он ответственен за синтез до 90% поли(ADP-рибозы) в клетке (Суханова и соавт., 2004). Фермент PARP1 вовлечен в процессы репликации (Cesarone et al., 1990), транскрипции (Meisterernst et al., 1997) и репарации (Dantzer et al., 1999). Повышенная активация PARP1 может приводить к существенному снижению содержания внутриклеточного NAD+ и, как следствие, гибели клетки (Skaper et al., 2003).
Таким образом, ген ADPRT1, кодирующий поли(АДФ-рибозил)полимеразу, возможно является одним из генов, определяющих наследственную предрасположенность к развитию ДПН у больных сахарным диабетом типа 1.
Ген ADPRT1, картированный на участке 1q41-1q42 хромосомы человека, кодирует полипептид PARP1, состоящий из двух функционально различающихся частей: N-концевого ДНК-связывающего и C-концевого каталитического доменов. Между ними находится домен аутомодификации (Cherney et al., 1987).
Обнаружен ряд полиморфных участков, как в самом гене ADPRT1, так и в его фланкирующих областях. Полиморфный маркер Leu54Phe гена ADPRT1 представляет собой однонуклеотидный полиморфизм C/G, которому соответствует аминокислотный полиморфизм Leu/Phe в положении 54 полипептидной цепи. Полиморфный маркер Val762Ala гена ADPRT1 представляет собой однонуклеотидный полиморфизм Т/С, которому соответствует аминокислотный полиморфизм Val/Ala в положении 762 полипептидной цепи. Данные об изучении ассоциации этих двух полиморфных маркеров гена ADPRT1 с развитием ДПН при СД типа 1 или каких-либо других заболеваний в литературе отсутствуют.
В группах пациентов с СД типа 1 с диабетической полинейропатией («ДПН+») и без этого осложнения («ДПН–») были определены частоты аллелей и генотипов полиморфного маркера Val762Ala гена ADPRT1. Сравнительный анализ выявил достоверные различия частот аллелей и генотипов данного полиморфного маркера в этих группах. Частоты аллелей полиморфного маркера Val762Ala в группах «ДПН+» и «ДПН–» были следующими: аллель Val – 0,820 и 0,934, аллель Ala – 0,180 и 0,066 соответственно. Частоты генотипов: в группе «ДПН+»: Val/Val – 0,740, Val/Ala – 0,160, Ala/Ala – 0,100; в группе «ДПН–»: Val/Val – 0,876, Val/Ala – 0,115, Ala/Ala – 0,009 (табл. 10).
Таблица 10.
Распределение частот аллелей и генотипов полиморфного маркера Val762Ala гена ADPRT1 в группах больных сахарным диабетом (СД) типа 1 с наличием («ДПН+») и отсутствием («ДПН–») диабетической полинейропатии
| Аллель и генотип | Частота аллелей и генотипов | Уровень значимости, p | OR | CI 95% | |
| «ДПН+» (n = 100) | «ДПН» (n = 113) | ||||
| Аллель Val | 0,820 | 0,934 | 0,0005 | 0,32 | 0,17 – 0,61 |
| Аллель Ala | 0,180 | 0,066 | 3,09 | 1,63 – 5,83 | |
| Генотип Val/Val | 0,740 | 0,876 | 0,014 | 0,40 | 0,20 – 0,82 |
| Генотип Val/Ala | 0,160 | 0,115 | 0,424 | 1,47 | 0,67 – 3,22 |
| Генотип Ala/Ala | 0,100 | 0,009 | 0,0034 | 12,44 | 1,56 –99,05 |
Носители аллеля Val и генотипа Val/Val имели пониженный риск развития ДПН (OR = 0,32 и 0,40, соответственно; p = 0,0005 и 0,014, соответственно), в то время как носители аллеля Ala и генотипа Ala/Ala имели повышенный риск развития ДПН (OR = 3,09 и 12,44, соответственно; p = 0,0005 и 0,0034, соответственно). Аминокислотный полиморфизм Val762Ala расположен в каталитическом домене PARP1. Возможно, изоферменты, содержащие в положении 762 остатки Ala или Val, имеют разные удельные активности, что приводит и к разным скоростям синтеза поли-АДФ-рибозных цепей.
В группах пациентов с СД типа 1 с диабетической полинейропатией («ДПН+») и без этого осложнения («ДПН–») были определены частоты аллелей и генотипов полиморфного маркера Leu54Phe гена ADPRT1. В обеих группах наибольшая частота была у аллеля Phe (0,655 в группе «ДПН+» и 0,535 в группе «ДПН»). Наиболее распространенным генотипом были гомозиготы Phe/Phe. Гомозиготные генотипы Leu/Leu встречались наиболее редко – их доля в группах больных СД типа 1 с наличием («ДПН+») и отсутствием («ДПН») диабетической полинейропатии составила 0,150 и 0,301, соответственно. Гетерозиготные генотипы Leu/Phe встречались с частотой 0,390 и 0,327, соответственно (табл. 11).
Таблица 11.
Распределение частот аллелей и генотипов полиморфного маркера Leu54Phe гена PARP в группах больных сахарным диабетом (СД) типа 1 с наличием («ДПН+») и отсутствием («ДПН–») диабетической полинейропатии
| Аллель и генотип | Частота аллелей и генотипов | Уровень значимости, p | OR | CI 95% | |
| «ДПН+» (n = 100) | «ДПН» (n = 113) | ||||
| Аллель Leu | 0,345 | 0,465 | 0,0136 | 0,61 | 0,41 – 0,90 |
| Аллель Phe | 0,655 | 0,535 | 1,65 | 0,11 – 2,44 | |
| Генотип Leu/Leu | 0,150 | 0,301 | 0,0095 | 0,41 | 0,21 – 0,81 |
| Генотип Leu/Phe | 0,390 | 0,327 | 0,391 | 1,31 | 0,75 – 2,30 |
| Генотип Phe/Phe | 0,460 | 0,372 | 0,211 | 1,44 | 0,83 – 2,49 |
Носители аллеля Leu и генотипа Leu/Leu имели пониженный риск развития ДПН (OR = 0,61 и 0,41, соответственно; p = 0,0136 и 0,0095, соответственно), в то время как носители аллеля Phe имели повышенный риск развития ДПН (OR = 1,65; p = 0,0136), так же как и носители генотипа Phe/Phe (OR = 1,44; р = 0,211). Можно предположить, что данный полиморфизм влияет на эффективность распознавания белком PARP1 разрывов ДНК.
Таким образом, нам удалось показать, что два полиморфных маркера гена ADPRT1 ассоциированы с развитием ДПН при СД типа 1 в русской популяции г. Москвы. Первый из этих маркеров, аминокислотный полиморфный маркер Val762Ala, расположен в каталитическом домене PARP1 и, возможно, носительство определенных аллелей этого маркера коррелирует с уровнем синтеза поли-АДФ-рибозных цепей. Второй из этих маркеров, аминокислотный полиморфный маркер Leu54Phe, расположен в первом из «цинковых пальцев» домена белка PARP1, взаимодействующего с двухцепочечными разрывами в ДНК. Можно предположить, что данный полиморфизм влияет на эффективность распознавания белком PARP1 разрывов ДНК.
ВЫВОДЫ.
- Изучено распределение аллелей и генотипов полиморфных маркеров Pro72Arg и C(–594)CC гена TP53, T(–786)C гена NOS3, G4077A гена GPX3, С825T гена GNB3, G(–25)A гена ANT1, G(–605)T гена PEO1, T(–365)C гена POLG1, Val762Ala и Leu54Phe гена ADPRT1 с ДПН при СД типа 1 у русских пациентов, проживающих в г. Москве.
- Для полиморфных маркеров C(–594)CC гена TP53, T(–786)C гена NOS3, G4077A гена GPX3, G(–25)A гена ANT1, G(–605)T гена PEO1 показано отсутствие ассоциации с ДПН при СД типа 1 у русских пациентов г. Москвы.
- Обнаружена ассоциация полиморфного маркера Pro72Arg гена TP53 с развитием ДПН при СД типа 1 у русских пациентов г. Москвы. Носители аллеля Arg имеют повышенный риск развития ДПН при СД типа 1 и генотипа Arg/Arg. Аллель Pro, напротив, ассоциирован с пониженным риском развития ДПН при СД типа 1.
- Обнаружена ассоциация полиморфного маркера С825Т гена GNB3 с развитием ДПН при СД типа 1 у русских пациентов г. Москвы. Риск развития патологии связан с носительством аллеля T. Аллель C, напротив, ассоциирован с пониженным риском развития ДПН при СД типа 1.
- Обнаружена ассоциация полиморфного маркера T(–365)C гена POLG1 с развитием ДПН при СД типа 1 у русских пациентов г. Москвы. Риск развития патологии оказался связан с носительством аллеля С. Аллель Т, напротив, ассоциирован с пониженным риском развития ДПН при СД типа 1.
- Показана ассоциация полиморфных маркеров Val762Ala и Leu54Phe гена ADPRT1 с развитием ДПН при СД типа 1 у русских пациентов г. Москвы. В случае полиморфного маркера Val762Ala носители аллеля Val и генотипа Val/Val имели пониженный риск развития ДПН, в то время как носители аллеля Ala и генотипа Ala/Ala имели повышенный риск развития ДПН. В случае полиморфного маркера Leu54Phe носители аллеля Leu и генотипа Leu/Leu имели пониженный риск развития ДПН, в то время как носители аллеля Phe имели повышенный риск развития ДПН.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ.
- Спицина, Е.В., Якунина, Н.Ю., Чудакова, Д.А., Никитин, А.Г., Светлова, Г.Н., Солуянова, Т.Н., Строков, И.А., Носиков, В.В. (2007) Ассоциация полиморфных маркеров Pro72Arg и C(–594)CC гена TP53 с диабетической полинейропатией при сахарном диабете типа 1 в русской популяции г. Москвы. Молекулярная биология, 41(6), 989-993.
- Spitsina, E.V., Yakunina, N.Yu., Chudakova, D.A., Nikitin, A.G., Svetlova, G.N., Soluyanova, T.N., Strokov, I.A., Nosikov, V.V. Association of polymorphous markers Pro72Arg and C(–594)CC of TP53 gene with diabetic polyneuropathy in patients with type 1 diabetes mellitus living in Moscow. Abstracts of the conference for young scientists, PhD students and students on molecular biology and genetics, dedicated to 120th anniversary of M.I.Vavilov, p. 190, Kyiv, Ukraine (September 20 – 22, 2007).
- Nosikov, V.V., Strokov, I.A., Nikitin, A.G., Spitsina, E.V., Tritschler, H.J. Poly (ADP-ribose) polymerase-1 gene (PARP1) contributes to the genetic predisposition to diabetic polyneuropathy in Russian patients with type 1 diabetes. Abstracts of the 41th Annual Meeting of European Association for the Study of Diabetes, p.A357 (Abstract N 984), Athens, Greece (September 10 – 15, 2005). Diabetologia, 48 [Suppl. 1], p.A357.
- Nosikov, V.V., Strokov, I.A., Spitsina, E.V., Svetlova, G.N., Nikitin, A.G., Kuraeva, T.L. Association of cell cycle control gene TP53 and DNA repair genes (POLG and PEO1) with diabetic polyneuropathy in Russian patients with type 1 diabetes. Abstracts of the 16th Annual Scientific Meeting of the Diabetic Neuropathy Study Group of the EASD (NEURODIAB), p.57 (Abstract P4), Ystad, Sweden (September 10 – 13, 2006).
- Строков, И.А., Никитин, А.Г., Спицина, Е.В., Светлова, Г.Н., Кураева, Т.Л., Якунина, Н.Ю., Носиков, В.В. Генетическая предрасположенность к диабетической полинейропатии при сахарном диабете типа 1. Материалы V Всероссийского конгресса эндокринологов “Высокие медицинские технологии в эндокринологии”, стр. 50, Москва, Россия (30 октября – 2 ноября 2006 г.).
- Spitsina, E.V., Yakunina, N.Yu., Chudakova, D.A., Nikitin, A.G., Svetlova, G.N., Soluyanova, T.N., Strokov, I.A., Nosikov, V.V. Association of polymorphous markers Pro72Arg and C(-594)CC of TP53 gene with diabetic polyneuropathy in patients with type 1 diabetes mellitus living in Moscow. Abstracts of the Conference for young scientists, PhD students and students on molecular biology and genetics, dedicated to 120th anniversary of M.I. Vavilov, p. 190, Kyiv, Ukraine (September 20 - 22, 2007).