

Роль сенсорных рнк в регуляции биосинтеза рибофлавина у escherichia coli и bacillus subtilis
На правах рукописи
Еремина
Светлана Юрьевна
Роль сенсорных РНК в регуляции биосинтеза рибофлавина у
Escherichia coli и Bacillus subtilis
03.00.15 – Генетика
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Москва
2008
Работа выполнена в лаборатории биохимической генетики Федерального государственного унитарного предприятия Государственного научно-исследовательского института генетики и селекции промышленных микроорганизмов «ФГУП ГосНИИгенетика»
Научный руководитель:
доктор биологических наук, профессор А.С. Миронов
ФГУП «ГосНИИгенетика»
Официальные оппоненты:
доктор биологических наук, профессор C.В. Машко
Научно-исследовательский институт
«Аджиномото-Генетика» (ЗАО «АГРИ»)
кандидат биологических наук И.В. Манухов
ФГУП «ГосНИИгенетика»
Ведущая организация: кафедра генетики биологического факультета
МГУ им. М.В. Ломоносова
Защита диссертации состоится «______»______________ 2008 года
на заседании Диссертационного совета Д 217.013.01 при ФГУП «Государственный научно-исследовательский институт генетики и селекции промышленных микроорганизмов»
по адресу: 117545, г. Москва, 1-й Дорожный проезд, д.1
С диссертацией можно ознакомиться в библиотеке ФГУП «ГосНИИгенетика»
Автореферат разослан «______»______________ 2008 года
Ученый секретарь
диссертационного совета
кандидат биологических наук Г.Г. Заиграева
ВВЕДЕНИЕ
Актуальность темы. Важной характеристикой жизнедеятельности микроорганизмов является использование широкого спектра регуляторных механизмов, обеспечивающих многообразие экспрессии генов в ответ на изменение условий внешней среды. Несколько лет назад был обнаружен новый тип регуляторных лидерных мРНК, выступающих в качестве сенсоров специфических метаболитов (малых молекул) и напрямую, без белковых посредников, способных регулировать транскрипцию и трансляцию соответствующих мРНК [Mironov A.S. et al., 2002, Winkler W. et al., 2002]. Такие сенсорные РНК представляют собой природные аптамеры и способны с высокой специфичностью и селективностью распознавать и связывать природные метаболиты, а также формировать альтернативные вторичные структуры в зависимости от присутствия специфического метаболита, включать или выключать процесс элонгации транскрипции мРНК или инициации трансляции этих мРНК.
Открытие нового механизма регуляции экспрессии генов, основанного на специфическом узнавании молекулой сенсорной РНК низкомолекулярных эффекторов, позволяет начать разработку принципиально новой стратегии получения штаммов-продуцентов целевых продуктов, так как эта регуляция осуществляется без участия регуляторных белков, путем прямого взаимодействия сенсорной РНК с низкомолекулярными соединениями, которые могут добавляться непосредственно в ростовую среду и модулировать экспрессию целевых генов. Это обстоятельство существенно упрощает задачу направленного изменения активности целевых генов в связи с тем, что молекула сенсорной РНК является значительно более компактной мишенью для модификации по сравнению со сложными молекулами регуляторных белков, действие которых приходится устранять для обеспечения высокого выхода желаемых продуктов в классических работах по конструированию штаммов-продуцентов. Можно использовать модифицированные сенсорные РНК для направленной реконструкции метаболизма бактерий. Это совершенно новый подход к управлению активностью генов, который позволит включать или выключать целевые гены с помощью сенсорных РНК путем использования простых метаболитов типа аминокислот, витаминов или даже ионов металлов, а в более далекой перспективе сможет послужить основой для создания нового поколения лекарственных препаратов на базе искусственно сконструированных молекул сенсорных РНК.
Несмотря на то, что биосинтез рибофлавина у Bacillus subtilis на протяжении многих лет был объектом интенсивных генетических и биохимических исследований, некоторые аспекты структурно-функциональной организации rib-оперона остаются неясными. С другой стороны, представлялось интересным сравнить роль сенсорных РНК в регуляции биосинтеза рибофлавина у Escherichia coli и Bacillus subtilis.
Цель и задачи исследования. Настоящая работа выполнялась с целью проведения исследований по расшифровке механизма регуляции с участием сенсорных РНК. В качестве объектов исследования использовались сенсорные РНК, кодируемые лидерными областями рибофлавинового (rib) оперона Bacillus subtilis и гена ribB Escherichia coli.
Достижение указанной цели было связано с решением следующих задач:
— компьютерный анализ вторичной структуры лидерных областей модельных сенсорных РНК;
— проведение сайт-направленного мутагенеза лидерных участков rib-оперона В. subtilis и гена ribB E. coli для получения модифицированных сенсорных РНК, позволяющих изучить механизм функционирования этих областей;
— конструирование транскрипционных и трансляционных фьюзов лидерной области rib-оперона B. subtilis и гена ribB E. coli с реперным геном lacZ;
— изучение экспрессии in vivo и in vitro мутантных вариантов лидерных областей rib-оперона B. subtilis и гена ribB E. coli c нуклеотидными заменами, приводящими к изменению конформации соответствующих сенсорных РНК.
Научная новизна. Результаты проведенного мутационного анализа лидерной области rib-оперона B. subtilis и лидерной области гена ribB E. coli позволили получить новую информацию для расшифровки механизма регуляции биосинтеза рибофлавина с участием сенсорных РНК. Показано, что специфические мутации в лидерной области rib-оперона B. subtilis, стабилизирующие формирование терминирующей шпильки, приводят к полной репрессии этого оперона. С другой стороны, мутации, нарушающие образование шпильки, обеспечивают конститутивную ФМН-независимую экспрессию rib-оперона. Эти данные уточняют модель регуляции rib-оперона B. subtilis с участием специфической ФМН-зависимой сенсорной РНК. В опытах in vitro показано, что ФМН модулирует вторичную структуру сенсорной РНК и влияет на уровень терминации транскрипции rib-оперона.
Впервые проведен структурно-функциональный анализ гена ribB E. coli. На основании данных сайт-направленного мутагенеза и анализа поведения полученных мутантов установлено, что лидерная область гена ribB E. coli кодирует специфическую ФМН-зависимую сенсорную РНК. Из результатов опытов по гибридизации 30S-субъединицы рибосомы с сенсорной РНК гена ribB можно сделать вывод о том, что ФМН участвует в формировании шпильки, перекрывающей сайт связывания рибосомы. На основе полученных данных была построена модель регуляции гена ribB E. coli, основанная на изменении конформации сенсорной РНК в зависимости от присутствия эффектора – ФМН. Согласно этой модели, связывание ФМН с сенсорной РНК стабилизирует шпилечную структуру, перекрывающую сайт связывания рибосомы, и тем самым блокирует инициацию трансляции гена ribB.
В ходе проведенной работы было установлено, что главное отличие механизмов регуляции экспрессии генов биосинтеза рибофлавина у B. subtilis и E. coli с участием ФМН-специфической сенсорной РНК состоит в том, что в rib-опероне B. subtilis контроль осуществляется на уровне терминации транскрипции, в то время как в гене ribB E. coli – на уровнe инициации трансляции.
Практическая значимость. Методом сайт-направленного мутагенеза получена коллекция мутантных вариантов лидерной области rib-оперона B. subtilis и лидерной области гена ribB E. coli с нуклеотидными заменами в участках лидерной мРНК, которые влияют на формирование вторичной структуры или на связывание мРНК с эффектором. Коллекция может быть использована для дальнейших исследований в этой области и для конструирования высокоактивных штаммов-продуцентов рибофлавина.
Структура работы. Диссертация изложена на 118 листах машинописного текста, включая 42 рисунка и 9 таблиц. Работа состоит из введения, обзора литературы, экспериментальной части, включающей описание материалов и методов, изложения и обсуждения результатов, выводов и списка используемой литературы. Список литературы включает 150 работ отечественных и зарубежных авторов.
Апробация работы. Материалы исследования по теме диссертации опубликованы в трех статьях. Диссертационная работа была апробирована на семинаре секции «Генетика микроорганизмов» Ученого Совета ФГУП «ГосНИИгенетика» 25 сентября 2008 года.
Материалы и методы
Бактериальные штаммы и плазмиды. Использованные в работе бактериальные штаммы и плазмиды, а также их генетические характеристики представлены в Таблице 1.
Таблица 1. Использованные в работе бактериальные штаммы и плазмиды.
| Штаммы, плазмиды | Генотип | Происхождение |
| B. subtilis: | ||
| B. subtilis 168 | Дикий тип | ВКПМ |
| B. subtilis RK99 | lys42 | Д.А. Перумов |
| RK998 ribB110 | lys42 ribB | « |
| RK996 ribC | lys42 ribC | « |
| E.coli K-12: | ||
| TG1 | thi supE hsd5 (lac-proAB)/F'tra 36 proAB+ lacIq lacZ M15 | ВКПМ |
| MG1655 | Дикий тип | ВКПМ |
| AM4001 | Как MG 1655, но ribB::Tn5, KmR | ВКПМ |
| AM4003 | Как MG 1655, но lacZ::Tn10, TcR | ВКПМ |
| AM4002 | Как MG 1655, но lacZ::Tn10ribB::Tn5, TcR, KmR | Данная работа |
| Плазмиды: | ||
| pDG268 | amyE lacZ AmpR CmR, присутствует SD последовательность | ВКПМ |
| pLZ | pDG268, содержащая EcoRI-BamHI лидерную область rib-оперона B. subtilis дикого типа | Данная работа |
| pLZ34, 35, 40, 49, 59, 87, 121, 128, 158, 161, 235/265, 79/189 | Как pLZ, но содержат нуклеотидные замены или делеции в лидерной области rib-оперона | » |
| рJEL250 | Малокопийный вектор, AmpR, присутствует SD последовательность | П. Валентин-Хансен (Дания) |
| рJEL246 | Малокопийный вектор, AmpR | П. Валентин-Хансен (Дания) |
| pBZ | pJEL246, содержащая EcoR1-BamH1 лидерную область гена ribB+ длиной 400 пн (от –120 до +280) | Данная работа |
| pBZ250 | pJEL250, содержащая EcoR1-BamH1 лидерную область гена ribB+ длиной 400 пн (от –120 до +280) | » |
| pBZM1, 2, 3, 4, 5, 6, 7, 8, 9, M6/M7 | Как pBZ, но содержат нуклеотидные замены или делеции в лидерной области гена ribB+ | » |
| pBZM9250 | Как pBZ250, но содержит делецию 8-235 | » |
Манипуляции с ДНК. Трансдукцию бактериофагом Р1 проводили по стандартной методике [Миллер Дж., 1979]. Выделение плазмидной ДНК, клонирование, трансформацию и анализ рекомбинантных плазмид проводили с использованием стандартных методов, изложенных в [Sambrook J. et al., 1989]. Компетентные клетки готовили по методу Манделя [Mandel M., Higa A., 1970].
Сайт-направленный мутагенез. Мутации в лидерной области rib-оперона B. subtilis и в лидерной области гена ribB E. coli получали методом сайт-направленного мутагенеза с помощью ПЦР, используя специфические праймеры.
Полимеразную цепную реакцию проводили на приборе Gene Amp PCR System 2400 (фирма "Perkin-Elmer Cetus"). Температурный режим подбирали с учетом длины амплифицированного фрагмента, длины и состава используемых праймеров. Выделение и очистку ПЦР продуктов проводили с использованием набора "Gene clean" (фирма "Bio 101", La Jolla, США) и GFXTM PCR DNA and Gel Band Purification Kit (фирма "GE Healthcare"). Присутствие соответствующих мутаций проверяли секвенированием по методу Сэнгера [Sanger F. et al., 1977].
Компьютерный анализ вторичной структуры лидерных мРНК. Потенциальные вторичные структуры мРНК, формирующиеся в процессе транскрипции, были смоделированы с помощью алгоритма Цукера-Тернера [Zuker D.H. et al., 1999].
Выделение суммарной РНК проводили методом гуанидинтиоционатной экстракции [Wickieser J.K, 2005].
Определение старта транскрипции проводили методом «достройки праймера» с использованием AMV ревертазы, меченного [P32] праймера и 70-80 мкг суммарной РНК согласно [Sambrook J. et al., 1989].
Транскрипция in vitro. Синтез лидерной мРНК rib-оперона дикого типа и ее мутантных вариантов, а также специфической лидерной мРНК гена ribB проводили по методике, описанной в работе [Franklund S.V., Kadner R.J., 1997] с использованием препарата холофермента РНК-полимеразы E. coli ("GE Healthcare").
Гибридизацию РНК с олигонуклеотидами проводили по методике, описанной в работе [Mironov A.S. et al., 2002].
Анализ формирования 30S-мРНК-тРНКfMet инициаторного комплекса in vitro (тоупринтинг) проводили по методике согласно [Hartz D. et al., 1988].
Измерение активности -галактозидазы. Бактериальные культуры выращивали при 30oС (для E. coli) или 37оС (для B. subtilis) в жидкой среде Спицайзена или M9 с необходимыми добавками и глюкозой (0,4%). Ночные культуры штаммов дважды отмывали, разводили в 20 50 раз в свежей минимальной среде, содержащей необходимое количество рибофлавина или его производных (ФАД, ФМН), и выращивали при 30оС или 37оС в течение 23 часов до экспоненциальной фазы роста. Активность -галактозидазы определяли по методу Миллера [Миллер Дж. 1979] и выражали в условных единицах, рассчитанных по формуле:
E = ОП420/ОП450·t.
Приведенные значения активности – средние из 4 независимых определений.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
1 Исследование регуляции рибофлавинового оперона Bacillus subtilis
К моменту начала настоящей работы появились первые данные, свидетельствующие о том, что негативная регуляция рибофлавинового (rib) оперона B. subtilis конечными продуктами биосинтеза рибофлавина осуществляется с помощью принципиально нового механизма аттенюации транскрипции. Настоящая работа является продолжением исследований по расшифровке механизма регуляции с участием сенсорных РНК и посвящена детальному анализу структурно-функциональной организации лидерной области rib-оперона B. subtilis, в частности, изучению влияния отдельных нуклеотидных замен в лидерной области rib-оперона на процесс фолдинга лидерной мРНК и на эффективность связывания с регуляторным метаболитом – ФМН.
1.1 Клонирование и определение нуклеотидной последовательности лидерной области rib-оперона B. subtilis
Прежде чем приступить к получению мутаций в лидерной области rib-оперона B. subtilis, необходимо было осуществить её клонирование. Лидерная область rib-оперона была амплифицирована с хромосомной ДНК штамма B. subtilis 168 методом ПЦР с использованием фланкирующих праймеров. Нуклеотидная последовательность данного фрагмента по обеим нитям была определена методом секвенирования по Сэнгеру. Полученная в результате секвенирования нуклеотидная последовательность представлена на Рис. 1. Нуклеотиды в области от +25 до +162 входят в состав так называемого rfn-элемента и обнаруживают высокий уровень гомологии при сравнении лидерных областей генов биосинтеза рибофлавина у различных таксономических групп бактерий. Кроме того, в пределах rfn-элемента содержатся, по крайней мере, 6 инвертированных повторов, способных формировать потенциальные шпилечные структуры. Важно подчеркнуть, что один из повторов (I) способен формировать шпильку либо с левым плечом терминатора (VII), способствуя образованию антитерминатора транскрипции, либо с последовательностью I', что может приводить к образованию альтернативной структуры анти-антитерминатора и, соответственно, терминатора транскрипции. Формирование шпильки I – I', вероятно, является решающим событием для последующего фолдинга лидерной мРНК и приводит к образованию терминирующей шпильки VII:VII', что, в конечном счете, обусловливает преждевременную терминацию транскрипции мРНК rib-оперона.
1.2 Сайт-направленный мутагенез лидерной области rib-оперона B. subtilis
Для дальнейшего подтверждения модели регуляции rib-оперона с участием ФМН проводился мутационный анализ лидерной области оперона. Сайт-направленный мутагенез проводили методом ПЦР с использованием специфических праймеров, содержащих нуклеотидные замены в участках лидерной мРНК, которые предположительно влияют на формирование ее вторичной структуры или на связывание сенсорной РНК с эффекторами (ФМН, ФАД). Расположение всех полученных мутаций в лидерной области rib-оперона показано на Рис. 1. Таким образом, были получены мутанты L34 (G34C) и L35 (G35C), содержащие такие замены в левом плече антитерминатора, которые нарушают формирование шпильки I:VII и одновременно обеспечивают дополнительную гомологию между нуклеотидами, формирующими шпильку I:I'. Мутанты L158 (158GА) и L161 (161GА), напротив, содержат замены, нарушающие формирование шпильки I:I' анти-антитерминатора, и тем самым увеличивают вероятность образования антитерминатора транскрипции (шпилька I:VII). Кроме того, получены три пары нуклеотидных замен, нарушающих формирование шпилек: II:II' – мутанты L40
RS1 -70 -50 -30
cccgaattc......
agggtgatgttttgtttttctcaaattgtaagtttatttcattgcgtactttaaaaagga
tcccactacaaaacaaaaagagtttaacattcaaataaagtaacgcatgaaatttttcct
I II
-10 +10 +30 CC A
....
tcgctataataaccaataaggacaaatgaataaagattgtatccttcggggcagggtgga
agcgatattattggttattcctgtttacttatttctaacataggaagccccgtcccacct
II’ III III’ IV
T+50 A +70 T +90
. ... ..
aatcccgaccggcggtagtaaagcacatttgctttagagcccgtgacccgtgtgcataag
ttagggctggccgccatcatttcgtgtaaacgaaatctcgggcactgggcacacgtattc
_________79-189_________
IV’ V V' VI VI’ I’
+110 A T +130 +150 A A
.. #2 .. . . #1
cacgcggtggattcagtttaagctgaagccgacagtgaaagtctggatgggagaaggatg
gtgcgccacctaagtcaaattcgacttcggctgtcactttcagacctaccctcttcctac
___________________________79-189__________________________
+170 +190 +210
......
atgagccgctatgcaaaatgtttaaaaatgcatagtgttatttcctattgcgtaaaatac
tactcggcgatacgttttacaaatttttacgtatcacaataaaggataacgcattttatg
_____79-189___________
VII VII’
+230 +250 +270
......
ctaaagccccgaattttttataaattcggggcttttttgacggtaaataacaaaagaggg
gatttcggggcttaaaaaatatttaagccccgaaaaaactgccatttattgttttctccc
235/265
+290 M E E Y Y M K L A L D L A K Q G E
......
gagggaaacaaatggaagagtattatatgaagctggccttagatcttgcgaagcagggcg
ctccctttgtttaccttctcataatatacttcgaccggaatctagaacgcttcgtcccgc
ctaggccc RS2
Рисунок 1. Нуклеотидная последовательность лидерной области rib-оперона B. subtilis.
Обычным прописным шрифтом выделены –10 и –35 последовательности промотора. Красными буквами обозначены консервативные основания rfn-элемента. Потенциальные шпилечные структуры – пунктирными стрелками и римскими цифрами. Голубым цветом выделен терминатор транскрипции (VII – VII'). Зеленым цветом – анти-антитерминатор (I – I'). Заглавными жирными буквами обозначены нуклеотиды, формирующие антитерминатор транскрипции (I – VII). Желтым цветом выделены последовательности инвертированных повторов. Заглавными жирными буквами и стрелками над основной последовательностью обозначены нуклеотидные замены, содержащиеся в геноме исследуемых мутантов. Стрелкой указан старт транскрипции. Рамками выделены сайт связывания рибосомы (SD) и старт трансляции. Делеции показаны нижним подчеркиванием и вертикальными стрелками. Подчеркнуты последовательности, комплементарные олигонуклеотидам #1 и #2. Нумерация нуклеотидов приведена, начиная со старта транскрипции rib-оперона. Обычными малыми буквами обозначены нуклеотидные последовательности праймеров RS1 и RS2.
(G40A) и L49 (C49T), III:III' – мутанты L59 (G59A) и L87 (C87T) и V:V' – мутанты L121 (G121A) и L128 (С128Т). Наконец, были получены два делеционных мутанта: L235/265, у которого практически полностью удален терминатор транскрипции (шпилька VII:VII'), и L79/189, содержащий делецию протяженного участка лидерной области от +79 до +189 нуклеотида (Рис. 1).
Далее было осуществлено клонирование мутаций в лидерной области rib-оперона в составе экспрессионного вектора pDG268 и их интеграция в хромосомный локус amyE штаммов B. subtilis RK99, B. subtilis RK998 ribB110 и B. subtilis RK996 ribC.
1.3 Изучение экспрессии транскрипционных фьюзов ribPL-lacZ в клетках штаммов B. subtilis
различного генотипаБыл исследован эффект полученных мутаций в лидерной области в составе транскрипционных фьюзов ribPL-lacZ. В качестве реципиента использовали ауксотрофные по рибофлавину штаммы: RK998 ribB110 с мутацией в структурном гене ribB, кодирующем рибофлавинсинтазу, и RK996 ribC с мутацией в гене ribC, контролирующем реакцию синтеза ФМН из рибофлавина.
Из данных Табл. 2, прежде всего, следует, что использование для компенсации ауксотрофности штамма RK998 ribB110 экзогенного рибофлавина в концентрации 0,1 мкM обеспечивает высокий уровень дерепрессии активности -галактозидазы в контрольном штамме с интактной лидерной областью (по сравнению с прототрофным штаммом B. subtilis RK99 – данные не приводятся). В то же время добавление флавинов приводит к существенному подавлению экспрессии этого гибридного оперона: 12-кратному в присутствии рибофлавина, 17-кратному в присутствии ФАД и максимальному – почти 20-кратному – на фоне экзогенного ФМН. Как и ожидалось, в клетках штамма RK998 ribB110 ярко проявляются свойства мутаций в лидерной области в отношении их реакции на добавление экзогенных флавинов. Так, в случае штаммов, содержащих делеции лидерной области (L235/265 и L79/189) или точковые мутации (L158 и L161), добавление экзогенных флавинов практически никак не сказывается на уровне экспрессии соответствующих фьюзов. Остальные мутанты также характеризуются существенным (10 15-кратным) снижением уровня регуляции под действием экзогенных флавинов по сравнению с контрольным штаммом.
С другой стороны, уже само по себе присутствие мутации ribC в геноме штамма RK996 приводит к конститутивной экспрессии -галактозидазы под контролем лидерной области дикого типа. Более того, у этого штамма не наблюдается какого-либо подавления экспрессии фьюза при добавлении высоких концентраций экзогенного рибофлавина, тогда как в присутствии ФМН и ФАД происходит примерно 5-кратное снижение активности -галактозидазы.
Таблица 2. Влияние рибофлавина и его производных на экспрессию транскрипционных фьюзов ribPL-lacZ в клетках штаммов
B. subtilis RK998 ribB110 и B. subtilis RK996 ribC
Мутант | Нуклео-тидная замена | Активность -галактозидазы в присутствии добавок | |||||||
| B. subtilis RK998 ribB110 | B. subtilis RK996 ribC | ||||||||
| Риб (0,1 мкM) | Риб (5 мкM) | ФМН (5 мкM) | ФАД (5 мкM) | Риб (1 мкM) | Риб (50 мкM) | ФМН (50 мкM) | ФАД (50 мкM) | ||
| wt | – | 550 | 44 (12,5) | 28 (19,6) | 32 (17,1) | 450 | 440 (1,0) | 88 (5,1) | 95 (4,7) |
| L34 | G34C | 11 | 10 (1,1) | 9 (1,2) | 10 (1,1) | 15 | 14 (1,1) | 12 (1,2) | 13 (1,1) |
| L35 | G35C | 16 | 14 (1,2) | 13 (1,2) | 15 (1,0) | 17 | 16 (1,0) | 15 (1,1) | 16 (1,0) |
| L40 | G40A | 340 | 218 (1,5) | 222 (1,5) | 234 (1,4) | 388 | 369 (1,0) | 272 (1,4) | 295 (1,3) |
| L49 | C49T | 398 | 290 (1,4) | 310 (1,3) | 327 (1,2) | 414 | 397 (1,0) | 328 (1,3) | 346 (1,2) |
| L59 | G59A | 457 | 265 (1,7) | 270 (1,7) | 263 (1,7) | 461 | 458 (1,0) | 313 (1,5) | 324 (1,4) |
| L87 | C87T | 413 | 235 (1,8) | 214 (1,9) | 228 (1,8) | 474 | 450 (1,0) | 288 (1,6) | 296 (1,6) |
| L121 | G121A | 216 | 156 (1,4) | 145 (1,5) | 161 (1,4) | 356 | 349 (1,0) | 207 (1,7) | 253 (1,4) |
| L128 | C128T | 238 | 169 (1,4) | 168 (1,4) | 171 (1,4) | 338 | 327 (1,0) | 225 (1,5) | 249 (1,4) |
| L158 | G158A | 198 | 164 (1,2) | 165 (1,2) | 160 (1,2) | 235 | 228 (1,0) | 198 (1,2) | 216 (1,1) |
| L161 | G161A | 190 | 173 (1,1) | 179 (1,1) | 181 (1,1) | 210 | 198 (1,0) | 165 (1,3) | 170 (1,2) |
| L235/265 | 235–265 | 478 | 445 (1,1) | 452 (1,1) | 459 (1,1) | 512 | 505 (1,0) | 484 (1,1) | 499 (1,0) |
| L79/189 | 79–189 | 165 | 161 (1,0) | 163 (1,0) | 159 (1,0) | 265 | 256 (1,0) | 260 (1,0) | 262 (1,0) |
Ночные культуры штамма B. subtilis, содержащие в составе хромосомы транскрипционные фьюзы ribP-lacZ, разводили в 50 раз в свежей среде Спицайзена и растили при 37оС до OD600=0.2, затем в соответствующие пробы добавляли рибофлавин, ФМН или ФАД в указанных концентрациях и растили еще 2 часа перед определением активности -галактозидазы. Активность -галактозидазы выражена в единицах Миллера. В скобках приведены значения уровня репрессии активности -галактозидазы в присутствии флавинов.
Приведенные значения активности – средние из 4 независимых определений.
Качественно такая же картина наблюдается в опытах с мутантами по лидерной области: экзогенный рибофлавин не оказывает сколько-нибудь заметного ингибирующего действия на их экспрессию, в то время как ФМН и ФАД ингибируют экспрессию у мутантов L40, L59, L87, L121, L128. Эти данные свидетельствуют о том, что негативными эффекторами регуляции rib-оперона служат фосфорилированные производные ФМН или ФАД, но не сам рибофлавин.
Таким образом, характер экспрессии реперного гена lacZ под контролем мутантных лидерных последовательностей полностью согласуется с моделью формирования альтернативных вторичных структур лидерной мРНК в зависимости от присутствия или отсутствия регуляторного метаболита ФМН.
Наиболее показательно в этом отношении поведение мутантов L34 и L35, содержащих нуклеотидные замены, которые нарушают формирование шпильки I:VII и одновременно обеспечивают дополнительную гомологию между нуклеотидами, формирующими шпильку I:I'. Именно у этих мутантов наблюдается существенное снижение базального уровня экспрессии реперного гена независимо от добавления экзогенных флавинов (Табл. 2). Из этого следует, что стабилизация шпилечной структуры I:I', которая обусловлена нуклеотидными заменами у мутантов L34 и L35, имеет решающее значение в метаболической регуляции лидерной области rib-оперона, поскольку необратимо приводит к формированию терминатора транскрипции. Анализ фенотипического проявления других мутаций в лидерной области показывает, что нарушение любой из исследованных шпилечных структур (I:I', II:II', III:III', V:V') приводит к снижению уровня метаболической регуляции rib-оперона производными рибофлавина. У мутантов с делециями (мутант L235/265 – делеция, перекрывающая терминатор транскрипции или мутант L79/189 – делеция, удаляющая существенную часть rfn-элемента) происходит устранение негативного действия флавинов. Таким образом, полученные данные позволяют заключить, что метаболическая регуляция rib-оперона осуществляется за счет изменения конфигурации вторичной структуры лидерной мРНК в зависимости от присутствия метаболитов-эффекторов – производных рибофлавина. Однако окончательное заключение о природе метаболита-эффектора на основании опытов in vivo сделать трудно из-за неконтролируемых реакций взаимопревращения флавинов в клетке. Для решения этого вопроса на следующем этапе работы проводили опыты по изучению транскрипции лидерной мРНК rib-оперона в системе in vitro.
1.4 Транскрипция лидерной мРНК rib
-оперона дикого типа и ее мутантных вариантов в системе in vitroБыло изучено влияние рибофлавина и его производных ФМН и ФАД на характер транскрипции лидерной мРНК, синтезируемой с ПЦР-фрагмента, содержащего лидерную область rib-оперона дикого и мутантных типов. В предварительных опытах было показано, что максимальное воздействие на терминацию транскрипции rib-оперона дикого типа обнаруживает ФМН, причем даже в небольших концентрациях (от 1 до 3 мкМ). Аналогичные опыты были выполнены на матрицах, содержащих полученные нами мутации в лидерной области rib-оперона. Как следует из данных, представленных на Рис. 2, в присутствии ФМН 65% мРНК, синтезированной с матрицы, содержащей интактную лидерную область, подвержено преждевременной терминации транскрипции (Рис. 2, дорожка 2). В полном соответствии с данными in vivo у мутантов L34 и L35 наблюдается приблизительно такой же процент преждевременной терминации транскрипции как у дикого типа в присутствии ФМН, причем добавление ФМН в транскрипционную смесь не приводит к заметному увеличению эффективности терминации (Рис. 2, дорожки 3 6).
| Мутант | wt | L34 | L35 | L158 | L161 | L235/265 | L79/189 |
| ФМН | – + | – + | – + | – + | – + | – + | – + |
| 360 255 |  |  |  |  | 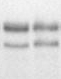 |  | |
| % T | 20 65 | 63 67 | 60 62 | 32 35 | 25 30 | 10 10 | 30 30 |
| № дорожки | 1 2 | 3 4 | 5 6 | 7 8 | 9 10 | 11 12 | 13 14 |
Рисунок 2. Эффект мутаций, затрагивающих формирование шпилечной структуры I:I', и делеций в лидерной области rib-оперона на уровень терминации транскрипции в системе in vitro (концентрация ФМН в пробах 5 мкМ).
Зеркальная картина наблюдается у мутантов L158 и L161: у обоих мутантов эффективность терминации транскрипции как в присутствии, так и в отсутствие ФМН составляет 25-35% (Рис. 2, дорожки 7 10), что примерно соответствует таковой у дикого типа в отсутствие ФМН. Примерно таким же уровнем терминации характеризуется мутант L79/189 с делецией, перекрывающей почти весь rfn-элемент, но не затрагивающей терминирующую шпильку VII:VII' (Рис. 2, дорожки 13, 14). Наконец, мутант L235/265 характеризуется наиболее сильным снижением эффективности терминации транскрипции (до 10%), что вполне объяснимо полным отсутствием в его лидерной области терминирующей шпильки VII:VII' (Рис. 2, дорожки 11, 12).
Результаты транскрипционных опытов с остальными мутантами представлены на Рис. 3. Как видно из представленных электрофореграмм, существенным отличием мутантных вариантов является их сниженная чувствительность к добавлению в транскрипционную систему ФМН по сравнению с диким типом.
| Мутант | wt | L40 | L49 | L59 | L87 | L121 | L128 |
| ФМН | – + | – + | – + | – + | – + | – + | – + |
| 360 255 |  | 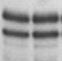 |  | 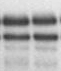 | 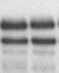 | 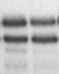 |  |
| % T | 18 68 | 45 46 | 36 32 | 24 37 | 31 35 | 39 46 | 41 51 |
| № дорожки | 1 2 | 3 4 | 5 6 | 7 8 | 9 10 | 11 12 | 13 14 |
Рисунок 3. Эффект мутаций, затрагивающих формирование различных шпилечных структур в лидерной области rib-оперона, на уровень терминации транскрипции в системе in vitro (концентрация ФМН в пробах 5 мкМ).
1.5 Опыты по гибридизации специфических олигонуклеотидов с лидерной мРНК rib-оперона
Таким образом, на основании полученных данных можно сделать важное заключение о том, что ФМН является ключевым метаболитом-эффектором rib-оперона и оказывает прямое действие на уровень терминации транскрипции rib-оперона, вероятно модулируя вторичную структуру лидерной мРНК. Для проверки этого предположения были проведены опыты in vitro по гибридизации специфических олигонуклеотидов с меченной [Р32] лидерной мРНК rib-оперона в присутствии ФМН, ФАД и рибофлавина или в их отсутствие. Один из олигонуклеотидов #1 комплементарен правому плечу шпильки I:I', а другой #2 – левому плечу шпильки V:V' (Рис. 1). Препараты РНК обрабатывали РНКазой H, которая расщепляет только двунитевые участки гибридов РНК/олигонуклеотид, а затем проводили электрофорез продуктов реакции в полиакриламидном геле.
| РНК-аза Н | – + + + + + + |
| ФМН | – – + – + – + |
| олигонуклеотид | – – – #1 #1 #2 #2 |
| 230 162 116 | 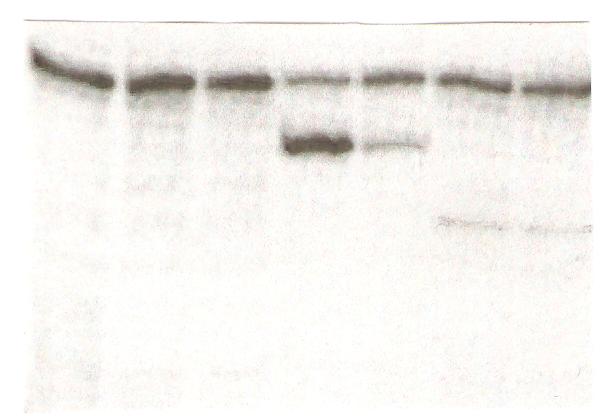 |
| % расщепления | – 0 0 69 33 10 10 |
| № дорожки | 1 2 3 4 5 6 7 |
Рисунок 4. Опыты по гибридизации лидерной мРНК rib-оперона со специфическими олигонуклеотидами в присутствии или отсутствие ФМН.
Из результатов этих опытов (Рис. 4), прежде всего, следует, что в отсутствие олигонуклеотидов не происходит расщепления меченой лидерной мРНК (размер 230 н.) под действием РНКазы H (дорожки 2, 3). При гибридизации мРНК с олигонуклеотидом #2 наблюдается появление дополнительного фрагмента мРНК (~116 н. – дорожки 6, 7), интенсивность которого примерно одинакова (~10% от контроля) независимо от присутствия или отсутствия в системе ФМН. Эти данные согласуются с моделью вторичной структуры лидерной мРНК. В случае олигонуклеотида #1 появляется дополнительный фрагмент (~162 н.), интенсивность которого существенно выше в отсутствие ФМН (69% от контроля – дорожка 4), чем в его присутствии (33% от контроля – дорожка 5). Из этого следует, что ФМН стабилизирует шпильку I:I', правое плечо которой должно гибридизоваться с праймером #1, тогда как в отсутствие ФМН эта шпилька разрушается и становится более доступной для отжига этого праймера.
Таким образом, совокупность полученных данных in vivo и in vitro подтверждают функциональную значимость нуклеотидных замен в лидерной области rib-оперона в формировании альтернативных структур терминатора и антитерминатора транскрипции и, кроме того, выявляют роль ФМН в качестве главного регуляторного метаболита в экспрессии rib-оперона B. subtilis.
2 Исследование регуляции гена ribB Escherichia coli
В отличие от rib-оперона B. subtilis, у E. coli за синтез рибофлавина отвечают шесть генов. Они не сцеплены на хромосоме, и об их регуляции известно очень немного. Высказывалось предположение, что экспрессия этих генов осуществляется конститутивно [Bacher A. et al., 1996]. Было интересно узнать, участвует ли rfn-элемент в регуляции биосинтеза рибофлавина у E. coli. С помощью компьютерного анализа нуклеотидных последовательностей регуляторных участков rib-генов E. coli выявили присутствие упомянутого выше консервативного rfn-элемента лишь перед одним геном – ribB, который кодирует синтез фермента 3,4-дигидрокси-2-бутанон-4-фосфатсинтазы [Vitreschak A.G. et al., 2002]. Эта часть работы посвящена мутационному анализу лидерной области гена ribB E. coli с целью выяснения механизма регуляции экспрессии гена ribB флавинами с участием специфической сенсорной РНК.
2.1 Клонирование и определение нуклеотидной последовательности лидерной области гена ribB Escherichia coli
Согласно предварительным данным [Mironov A.S. et al., 2002] лидерная область гена ribB кодирует сенсорную РНК, которая по своей структуре похожа на сенсорную РНК rib-оперона B. subtilis, но, в отличие от последней, не содержит Rho-независимого терминатора транскрипции (Рис. 5).
Сравнение нуклеотидной последовательности лидерной области ribB c последовательностью лидерной области rib-оперона B. subtilis выявляет высокий уровень гомологии в районе rfn-элемента (нуклеотиды от +3 до +147) (Рис. 5). Более того, лидерная область содержит три пары антипараллельных последовательностей, которые способны формировать потенциальные альтернативные шпилечные структуры: I:I' и III:III', либо II:II'. Расположение этих потенциальных шпилек практически полностью совпадает с таковым в лидерной области rib-оперона B. subtilis. Важное отличие состоит в том, что шпилечная структура III:III' в лидерной области гена ribB E. coli не включает характерного для rib-оперона B. subtilis Rho-независимого терминатора транскрипции, а вместо этого перекрывает сайт связывания рибосомы (SD) и сайт инициации трансляции (ATG).
Таким образом, можно предположить, что лидерная область гена ribB кодирует сенсорную РНК, которая в присутствии флавинов (ФМН, ФАД) формирует шпилечную структуру III:Ш', подавляющую трансляцию гена ribB, а в отсутствие этих эффекторов происходит образование альтернативной конфигурации лидерной мРНК с разрушением шпильки III:Ш', при которой осуществляется нормальная трансляция мРНК.
Модель двух наиболее вероятных альтернативных вторичных структур лидерной мРНК гена ribB представлена на Рис. 6. Одна из конфигураций (в присутствии эффектора – ФМН) действительно образуется за счет комплементарного взаимодействия последовательностей I и I' (обозначены зелеными стрелками на Рис. 5, шпилька анти-антисеквестора на Рис. 6) и последовательностей III и III' (фиолетовые стрелки на Рис. 5, шпилька секвестора трансляции на Рис. 6). Образование альтернативной конфигурации (в отсутствие ФМН) происходит за счет формирования шпильки II:II' (синие стрелки на Рис. 5, шпилька антисеквестора на Рис. 6). В соответствии с моделью, формирование антисеквестора (шпильки II:II') приводит к разрушению секвестора (III и III') и обеспечивает доступность сайта SD для связывания рибосомы и, следовательно, возможность последующей инициации трансляции структурного гена ribB.
2.2 Сайт-направленный мутагенез лидерной области гена ribB E. coli
Для проверки модели регуляции гена ribB, приведенной на Рис. 6 и основанной на изменении конформации сенсорной РНК, был проведен сайт-направленный мутагенез лидерной области гена ribB. Сайт-направленный мутагенез проводили методом ПЦР с использованием специфических праймеров, содержащих нуклеотидные замены в участках лидерной мРНК, которые предположительно влияют на формирование ее вторичной структуры или на связывание сенсорной РНК с эффекторами (ФМН, ФАД). Расположение всех полученных мутаций в лидерной области гена ribB показано на Рис. 5 и 6.

+ 1 cCc (M1)
-35 -10.. . ttgctgccgctaatcattagcgttatagtgaatccgcttattctcagggcggggcgaaat +25
#1
I
II
......
tccccaccggcggtaaatcaactcagttgaaagcccgcgagcgctttgggtgcgaactca +85
#2
A A A (M2)
.....
aaggacagcagatccggtgtaattccggggccgacggttagagtccggatgggagagagt +145
#4 #3
I’
. .. . ..
aacgattctgtcgggcatggacccgctcacgttattttggctatatgccgccactcctaa +205
AA A(M4)
GAC(M5) CC (M6) CC C(M3) GG(M7)
. .. .. M N Q T RibB gactgccctgattctggtaaccataattttagtgaggtttttttaccatgaatcagacg +265
<<<<<<<<<<<<<<<<<<<<<<<<<< M8
III III’
II’
Рисунок 5. Нуклеотидная последовательность лидерной области гена ribB E. coli.
Заглавными буквами выделены – 35 и – 10 последовательности промотора. Красными буквами обозначены консервативные основания rfn-элемента. Желтым цветом выделены последовательности, способные формировать шпилечные структуры. Заглавными буквами и стрелками над основной последовательностью обозначены нуклеотидные замены, полученные с помощью сайт-направленного мутагенеза. Подчеркнуты последовательности, комплементарные олигонуклеотидам #1, #2, #3, #4. Стрелкой указан старт транскрипции. Рамками выделены сайт связывания рибосомы (SD) и старт трансляции. Зелеными стрелками (I и I') обозначен анти-антисеквестор, синими стрелками (II и II') – антисеквестор, фиолетовыми стрелками (III и III') – секвестор трансляции. Нумерация нуклеотидов приведена, начиная со старта транскрипции гена ribB E. coli.
I:I’ III:III’ II:II’
G
A U
A C U:G
+ФМН M4,3 U-A
A C U:G SD -ФМН
A C U:G
A-U
A-U
U:U
G+17 G+133 A-U
G A СC:U
C U A-U G+21 A+204
C G G A-U C A
M1 C G G U-A G G
C G G C G-C G G A
A A M6 C G-C G M7 G-C
C-G A U-AU G:U
U-A C-G C-G
C-G A M2 U-A C G-C
U-A U-A M1 C G-C
U:G A A-U C G-C G
A-U C G-C A-U A M5
U-A M5 A U-A C-G C AA A M4
U-A+147 G C-G U-A+216 SD
+1GC CG GACUGCC+212 ACGC+265 +1GCUUAUUC+8 UUCUGGUAACCAUAAUUUUAGUGAGGUUUUUUUACCAUGAAUCAGACGC
CC M6 CC C M3 GG M7
Рисунок 6. Модель регуляции гена ribB, основанная на изменении конформации сенсорной РНК в зависимости от присутствия
эффектора – ФМН.
Нуклеотиды, входящие в состав анти-антисеквестора, обозначены зеленым цветом; антисеквестора – синим, секвестора – фиолетовым. Стрелками показаны нуклеотидные замены использованных в работе мутантов.
Исходя из этой модели, в присутствии ФМН стабилизируется шпилечная структура I и I' (анти-антисеквестор), что влечет за собой формирование шпилечной структуры III и III' (секвестор трансляции), перекрывающей сайт связывания рибосомы (SD) и стартовый кодон трансляции. В отсутствие ФМН происходит формирование альтернативной шпилечной структуры II и II' (антисеквестор), которая приводит к разрушению секвестора (шпилька III и III') и, тем самым, делает сайт SD доступным для связывания рибосомы и последующей инициации трансляции.
Таким образом, мутант M1 (G12C, G13C, G14C) содержит замены в области антисеквестора (левое плечо шпильки II:II'), тогда как мутант M2 (G140A, G142A, G144A) – в области анти-антисеквестора (правое плечо шпильки I:I'). Кроме того, получены пять мутантов, содержащих замены в области секвестора трансляции, причем четыре из них расположены в левом плече шпильки III:III': М3 (T232C, T233C, T235C); М4 (T232A, T233A, T235A); М5 (C213G, T214A, G215C) и М6 (G221C, G222C). Один мутант М7 содержит замены C251G, C252G в правом плече шпильки III:III'.
Важно подчеркнуть, что мутант М7 содержит симметричные нуклеотидные замены, компенсирующие эффект мутаций G221C, G222C (мутант М6), т.е. у двойного мутанта М6/М7 должна восстанавливаться шпилечная структура III:III' секвестора трансляции (Рис. 6). Наконец, были получены два делеционных мутанта: M8, у которого удалено 26 п.н. (210-235), т.е. практически все левое плечо (III) секвестора, а также делеционный мутант М9 с полностью делетированной лидерной областью. Полученные мутантные варианты лидерной области гена ribB клонировались в составе векторов pJEL250 и pJEL246 для изучения влияния полученных мутаций на экспрессию гена ribB.
2.3 Изучение экспрессии транскрипционных и трансляционных фьюзов ribB-lacZ в клетках E. coli
Экспрессионные векторы pJEL250 и pJEL246, содержащие, соответственно, транскрипционные и трансляционные фьюзы ribB-lacZ (Табл. 1) с помощью трансформации переносили в реципиентный штамм AM4002 ribB::Tn5 (KmR) lacZ::Tn10 (TcR). У полученных рекомбинантов определяли активность -галактозидазы в условиях репрессии активности гена ribB (среда с 300 мкM рибофлавина) и его дерепрессии (среда с 3 мкM рибофлавина). Прежде всего мы сравнили уровень репрессии активности -галактозидазы рибофлавином у штаммов, содержащих плазмиды pJEL250 и pJEL246, в которые был клонирован один и тот же фрагмент интактной лидерной области гена ribB. В условиях репрессии (300 мкМ рибофлавина) активность -галактозидазы у транскрипционного фьюза ribB-lacZ снижается примерно в 1,5 раза (Рис. 7), в то время как у трансляционного фьюза наблюдается почти 40-кратное падение активности фермента (Рис.7, Табл. 3). В то же время присутствие делеции, перекрывающей всю лидерную область гена ribB (М9), приводит к примерно одинаковой конститутивной экспрессии как транскрипционного, так и трансляционного фьюзов ribB-lacZ даже при наличии высоких концентраций рибофлавина в среде (Рис. 7, Табл. 3). Эти данные указывают на то, что репрессирующее действие рибофлавина (или его производных) осуществляется посредством их взаимодействия с лидерной областью гена ribB и реализуется, вероятней всего, на трансляционном уровне. Поэтому в дальнейших опытах по изучению влияния полученных мутаций в лидерной области на экспрессию гена ribB мы использовали только трансляционные фьюзы ribB-lacZ.
Анализ данных Табл. 3 подтверждает модель регуляции гена ribB, основанную на изменении конформации сенсорной РНК в зависимости от присутствия эффекторов – рибофлавина или его производных (Рис. 6). В соответствии с этой схемой у мутанта М1 присутствие нуклеотидных замен в левом плече антисеквестора (шпилька I) приводит к резкому (на несколько порядков) снижению активности -галактозидазы независимо от присутствия или отсутствия рибофлавина в среде (Табл. 3). Важно подчеркнуть, что эти нуклеотидные замены (G12C, G13C, G14C) не только нарушают гомологичное спаривание нуклеотидов в шпильке II:II' антисеквестора, но одновременно увеличивают область спаривания оснований в шпильке I:I анти-антисеквестора. В свою очередь, формирование более протяженной шпильки I:I' способствует стабилизации шпильки III:III', перекрывающей сайт SD и кодон ATG, что, в конечном счете, практически полностью подавляет трансляцию гена ribB (Рис.6).
Рисунок 7. Сравнение уровня активности -галактозидазы у штаммов, содержащих транскрипционные (250) и трансляционные (246) фьюзы ribB-lacZ с лидерной областью дикого типа (wt) и ее делеционного варианта (М9).
К противоположному эффекту приводят нуклеотидные замены мутанта М2 (G140A, G142A, G144A), которые, в отличие от замен мутанта М1, не способствуют, а препятствуют формированию шпильки I:I' анти-антисеквестора и, следовательно, секвестора трансляции (шпилька III:III'). В результате у мутанта М2 увеличивается вероятность формирования шпильки II:II' антисеквестора и повышается ее стабильность. В соответствии с этим, у мутанта М2 наблюдается хотя и более низкий по сравнению с диким типом, но существенно более высокий по сравнению с мутантом М1, уровень экспрессии ribB. При этом репрессирующее действие рибофлавина на экспрессию ribB у мутанта М2 существенно снижается (в 1,5 раза против 37 у дикого типа – Табл. 3).
Таблица 3. Активность -галактозидазы у штаммов, содержащих мутации в
лидерной области гена ribB, в зависимости от концентрации рибофлавина в среде.*
Мутация | Нуклеотидные замены | Активность -галактозидазы | Уровень репрессии | |
| Риб (3мкМ) | Риб (300мкМ) | |||
| wt | – | 2600143 | 704 | 37 |
| M1 | G12C,G13C,G14C | 7,00,6 | 6,00,4 | 1,1 |
| M2 | G140A, G142A, G144A | 23010 | 1507 | 1,5 |
| M3 | T232C, T233C, T235C | 6,0 0,5 | 6,00,4 | 1,0 |
| M4 | T232A, T233A, T235A | 154062 | 864 | 18 |
| M5 | C213G, T214A, G215C | 140057 | 55018 | 2,5 |
| M6 | G221C, G222C | 5500176 | 90039 | 6,1 |
| M7 | C251G, C252G | 4500155 | 80043 | 5,6 |
| M6/M7 | G221C, G222C C251G, C252G | 80032 | 403 | 20 |
| M8 | (210-235) | 5400170 | 213096 | 2,5 |
| M9 | (8-235) | 5580237 | 5470223 | 1,0 |
* Сравнение уровня активности -галактозидазы у штаммов, содержащих трансляционные фьюзы ribB-lacZ с лидерной областью дикого типа (wt) и ее мутантных вариантов. Приведенные значения активности – средние из 4 независимых определений.
Функциональная роль шпильки III:III' в качестве секвестора трансляции подтверждается поведением мутанта М8, содержащего делецию левого плеча этой шпильки (Рис. 6). Разрушение шпильки III:III' у этого мутанта приводит к тому, что уровень активности -галактозидазы достигает почти максимального уровня и репрессируется рибофлавином всего в 2,5 раза (Табл. 3). Примерно такой же уровень репрессии наблюдается у мутанта М5, содержащего нуклеотидные замены в основании шпильки III:III', которые и приводят к сокращению ее протяженности (Рис. 6). Несколько более высокий уровень репрессии наблюдается у мутантов М6 и М7, содержащих симметричные замены в плечах III (G221C, G222C) и III' (C251G, C252G) секвестора. Примечательно, что комбинация этих мутаций, в результате которой восстанавливается комплементация в соответствующем участке шпильки III:III' (мутант М6/М7), приводит к существенному восстановлению уровня репрессии рибофлавином (с 6 до 20 раз – Табл. 3). Эти данные еще раз подчеркивают решающую роль шпильки III:III' в подавлении экспрессии гена ribB флавинами.
Особого внимания заслуживает поведение мутанта М3, содержащего нуклеотидные замены T232C, T233C, T235C, которые обеспечивают более жесткое спаривание антипараллельных цепей шпильки III:III' в сайте связывания рибосомы SD (Рис. 6). Активность -галактозидазы у этого мутанта характеризуется минимальным значением и свидетельствует о практически полном подавлении трансляции даже в отсутствие рибофлавина (Табл. 3). В то же время у мутанта М4, содержащего замены T232A, T233A, T235A, которые, напротив, снижают эффективность спаривания оснований в районе сайта SD, уровень репрессии рибофлавином ниже, чем у дикого типа. Таким образом, полученные данные согласуются с моделью регуляции гена ribB, основанной на формировании альтернативных вторичных структур сенсорной РНК в зависимости от присутствия эффекторов – рибофлавина или его производных. В случае rib-оперона B. subtilis таким эффектором является ФМН и в меньшей степени ФАД, но не рибофлавин. Для выяснения природы эффектора в случае гена ribB E. coli были проведены опыты in vitro.
2.4 Опыты in vitro по идентификации эффекторов сенсорной РНК гена ribB
Для выявления эффектора, модулирующего экспрессию гена ribB E. coli, проводили опыты in vitro по гибридизации специфических олигонуклеотидов, комплементарных различным участкам лидерной мРНК (Рис. 5), с меченной [Р32] лидерной мРНК гена ribB в присутствии ФМН, ФАД и рибофлавина или в их отсутствие. Препараты РНК обрабатывали РНКазой H, которая расщепляет только двунитевые участки гибридов РНК/олигонуклеотид, а затем проводили электрофорез продуктов реакции в полиакриламидном геле.
Из данных электрофореграммы, представленной на Рис. 8, прежде всего следует, что добавление ФМН (или рибофлавина и ФАД – данные не представлены) не приводит к появлению преждевременно терминированных транскриптов: на дорожках 1 и 2 видны транскрипты (~275 н.), соответствующие полноразмерной лидерной РНК. Эти данные подтверждают сделанное ранее заключение о том, что регуляция гена ribB с участием сенсорной РНК осуществляется не на уровне терминации транскрипции, а путем подавления инициации трансляции. При гибридизации РНК с олигонуклеотидами #1 (дорожки 3, 4) и #4 (дорожки 9, 10) также не наблюдается появления дополнительных транскриптов – продуктов расщепления РНКазой Н. Из этого следует, что области лидерной мРНК, комплементарные этим олигонуклеотидам, скорее всего, сильно структурированы как в присутствии, так и в отсутствие ФМН и поэтому недоступны для отжига этих олигонуклеотидов.
| РНКаза Н | |||||
| олиг | – | #1 | #2 | #3 | #4 |
| ФМН | – + – + – + – + – + | ||||
| 275 140 65 | 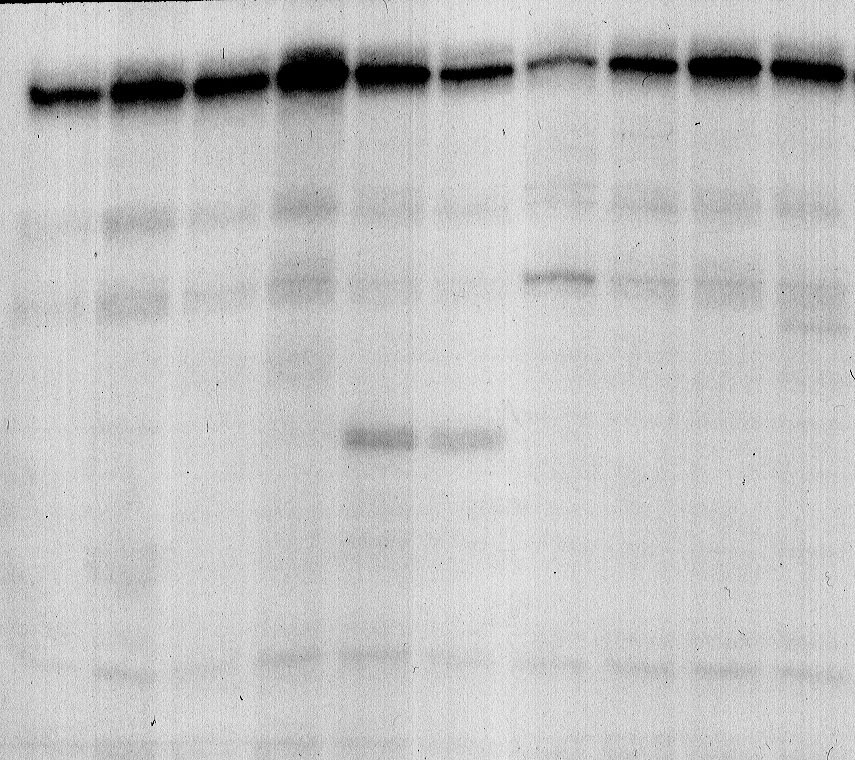 | ||||
| % расщепления | 0 0 0 0 27 25 43 5 0 0 | ||||
| № дорожки | 1 2 3 4 5 6 7 8 9 10 | ||||
Рисунок 8. Опыты по гибридизации сенсорной РНК со специфическими олигонуклеотидами: #1, #2, #3, #4 в присутствии ФМН (дорожки 4, 6, 8, 10) или в отсутствие ФМН (дорожки 3, 5, 7, 9) и последующей обработкой РНКазой Н.
Дорожки 1, 2 – контрольные пробы без добавления олигонуклеотидов (в отсутствие и в присутствии ФМН, соответственно).
Цифрами перед стрелками указаны размеры фрагментов РНК, получаемых после обработки препаратов РНКазой Н.
При гибридизации РНК с праймером #2 наблюдается появление дополнительного фрагмента РНК (~65 н. – дорожки 5, 6), интенсивность которого примерно одинакова (25-27% от контроля) независимо от того, присутствует ли в системе ФМН или нет. В случае праймера #3 появляется дополнительный фрагмент (~140 н.), интенсивность которого существенно выше в отсутствие ФМН (43% от контроля – дорожка 7), чем в его присутствии (5% от контроля – дорожка 8). Из этого следует, что ФМН стабилизирует шпильку I:I, левое плечо которой должно гибридизоваться с праймером #3, тогда как в отсутствие ФМН эта шпилька разрушается и становится более доступной для отжига этого праймера. Опыты по гибридизации с использованием в качестве эффектора ФАД дали похожую картину, но при значительно больших концентрациях ФАД, тогда как добавление рибофлавина не приводило к снижению интенсивности полосы транскрипта размером 140 н. в случае праймера #3 (данные не представлены).
Таким образом, результаты гибридизации свидетельствуют о том, что основным эффектором сенсорной РНК гена ribB является ФМН и, кроме того, указывают на прямое взаимодействие ФМН со шпилечной структурой I:I' в соответствии с моделью регуляции, представленной на Рис. 6.
2.5 Влияние флавинов на формирование 30S-инициаторного комплекса на лидерной мРНК гена ribB
Формирование 30S-инициаторного комплекса на лидерной мРНК гена ribB и его зависимость от присутствия флавинов анализировались методом тоупринт-анализа. В качестве праймера использовали олигонуклеотид R63, комплементарный участку (+283) (+303) ribB мРНК. Как следует из данных Рис. 9, тоупринт-сигнал в районе +16 нуклеотида мРНК появляется только при одновременном присутствии в реакционной смеси 30S-субъединицы и тРНКfMet (дорожка 3). Добавление ФМН, начиная с концентрации 40 мкМ (дорожка 8), снижает интенсивность тоупринт-сигнала (дорожки 7 10), а при концентрации 60-100 мкМ (дорожки 9, 10) ФМН практически полностью подавляет формирование инициаторного комплекса. Добавление ФАД приводит к снижению интенсивности тоупринт-сигнала только в высокой концентрации – 100 мкМ (дорожка 16), тогда как в присутствии рибофлавина (дорожки 17 22) ослабления тоупринт-сигнала вообще не наблюдается.
Таким образом, полученные данные однозначно свидетельствуют о том, что ФМН и в меньшей степени ФАД, но не рибофлавин подавляют формирование 30S-инициаторного комплекса на лидерной мРНК гена ribB. Эти данные полностью подтверждают модель регуляции гена ribB, в соответствии с которой связывание эффекторов ФМН и ФАД с сенсорной РНК стабилизирует шпилечную структуру, перекрывающую сайт связывания рибосомы, и тем самым блокирует инициацию трансляции.
| Флавины | – | ФМН | ФАД | Риб | |
| Конц.(мкМ) | – – – | 50 5 10 20 40 60 100 | 50 20 40 60 80 100 | 50 20 40 60 80 100 | |
| S30 | + – + | + + + + + + + | + + + + + + | + + + + + + | |
| тРНКfMet | – + + | – + + + + + + | – + + + + + | – + + + + + | |
| C U A G | 1 2 3 | 4 5 6 7 8 9 10 | 1112 13 14 15 16 | 17 18 19 20 2122 | |
A C C 0 A U G A A U C A G A C G C U A C +16U U U | 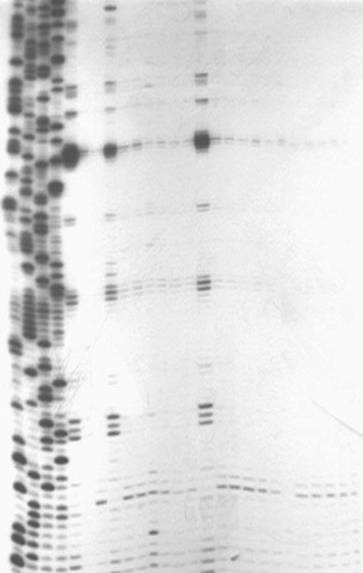 | ||||
Рисунок 9. Зависимость формирования 30S-инициаторного комплекса на лидерной мРНК гена ribB от присутствия флавинов.
C, U, A, G – сиквенсные дорожки. Слева приведена последовательность мРНК, перекрываемая 30S-субъединицей рибосомы. Стрелками показано положение А0 и С+16 нуклеотидов. Пунктирной стрелкой показано положение тоупринт-сигнала. Контрольные дорожки (№ 1, 4, 11, 17) не содержат тРНКfMet. Концентрации флавинов указаны в верхней части рисунка.
ВЫВОДЫ
- Лидерные области rib-оперона B. subtilis и гена ribB E. coli кодируют сенсорные РНК, способные специфически связываться с производными флавинов и модулировать экспрессию прилегающих структурных генов.
- ФМН является главным эффектором, специфически связывающимся с сенсорными РНК rib-оперона B. subtilis и гена ribB E. coli.
- Связывание ФМН с сенсорными РНК приводит к изменению их конформации, что, в свою очередь, обусловливает подавление экспрессии прилегающих генов.
- Регуляция экспрессии rib-оперона B. subtilis с участием специфической сенсорной РНК и ФМН осуществляется на уровне терминации транскрипции.
- Регуляция экспрессии гена ribB E. coli с участием специфической сенсорной РНК и ФМН осуществляется на уровне инициации трансляции.
Основные результаты работы отражены в следующих публикациях:
1. Миронов А.С., Еремина С.Ю. 2007. Фундаментальные и прикладные аспекты исследования регуляции оперона биосинтеза рибофлавина у бацилл. Биотехнология, №5, с.7 – 13.
2. Миронов А.С., Карелов Д.В., Соловьева И.М., Еремина С.Ю., Эрраис Лопес Л., Кренева Р.А., Перумов Д.А. 2008. Исследование связи вторичной структуры и регуляторной активности лидерного транскрипта оперона биосинтеза рибофлавина у Bacillus subtilis. Генетика, т.44, №4, с.1 – 7.
3. Еремина С.Ю., Золотухина М.А., Эрраис Лопес Л., Миронов А.С. 2008. Мутационный анализ лидерной области гена ribB Escherichia coli. Биотехнология, №2, с.14 – 22.