

Статистические потенциалы атомарной гидратации биополимеров
На правах рукописи
РАХМАНОВ СЕРГЕЙ ВИКТОРОВИЧ
СТАТИСТИЧЕСКИЕ ПОТЕНЦИАЛЫ
АТОМАРНОЙ ГИДРАТАЦИИ БИОПОЛИМЕРОВ
03.00.03 – молекулярная биология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Москва – 2008
Работа выполнена в лаборатории биоинформатики ФГУП «Государственный научно-исследовательский институт генетики и селекции промышленных микроорганизмов (ФГУП «ГосНИИгенетика»).
| |||||||||
Защита состоится 16 декабря 2008 г. в 14 часов на заседании Диссертационного совета Д.217.013.01 при Государственном научно-исследовательском институте генетики и селекции промышленных микроорганизмов по адресу: 117545, Москва, 1-й Дорожный проезд, 1.
С диссертацией можно ознакомиться в библиотеке ФГУП «ГосНИИ генетика».
втореферат разослан «___» _________ 200__ г.
| Учёный секретарь диссертационного совета, кандидат биологических наук | Г.Г. Заиграева |
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Изучение гидратации макромолекул биополимеров – белков и нуклеиновых кислот - необходимо для понимания механизмов функционирования этих молекул. Одним из путей решения этой задачи является вычислительное моделирование внутримолекулярных взаимодействий.
В виду практической невозможности провести прямое моделирование межатомных взаимодействия в макромолекулах, основываясь непосредственно на физических законах, основные усилия исследователей в этой области направлены на создание упрощённых эмпирических математических моделей взаимодействия атомов. Одним из классов моделей такого рода являются статистические модели взаимодействий, также часто называемые моделями, основанными на знаниях; при их построении производится статистическая обработка данных по большим количествам экспериментально определённых трёхмерных структурах биополимеров. При этом большое значение имеет нулевая гипотеза, на которой базируется статистическая обработка этих экспериментальных данных. В качестве таковой выстуает так называемое референсное состояние макромолекулы, в котором межатомные взаимодействия "выключаются". Ожидаемое распределение статистических параметров (в нашем случае межатомных расстояний) для такого референсного состояния используется для оценки интенсивности реальных межатомных взаимодействий.
В то время как основные усилия исследователей в последние десятилетия были в основном направлены на изучение взаимодействия собственно молекул биополимеров и составляющих их атомов между собой, описание функционирования белков и нуклеиновых кислот в водной среде оставалось сравнительно менее разработанной областью, несмотря на определяющую роль воды в определении укладки в и взаимодействия большинства биологических макромолекул.
В то же время успехи в описании белковой сольватации могут быть использованы для решения ряда прикладных задач, в частности предсказание расположения связанной воды вокруг белков. Знание местоположения молекул связанной воды позволяет повысить качество построения интерфейсов белковых молекул в задачах оценки взаимодействия белков с другими белками, нуклеиновыми кислотами, малыми молекулами и лигандами, а также другими макромолекулами.
Кроме того, определяющая роль гидратации в формировании пространственной структуры белка позволяет использовать статистические модели относительно взаимодействия атомов белка с молекулами воды для выделения правильной укладки белка среди множества неправильных вариантов укладки. Такая задача возникает в ряде алгоритмов моделирования пространственных структур белков, т.е. так называемой задачи моделирования фолдинга.
Цель и задачи работы.
При построении статистических моделей межатомных взаимодействий основные трудности заключаются в построении т.н. референсных, или невзаимодействующих состояний, т.е. состояний с нулевой энергией взаимодействия (СНЭВ), характеризующих ожидаемое распределение контактных частот в случае "выключенных внутримолекулярных взаимодействий". Имея в руках удачное СНЭВ, можно достаточно однозначно построить эмпирические потенциалы межатомного взаимодействия. В настоящей работе для построения СНЭВ предложено использовать метод стохастического моделирования, также называемый методом Монте-Карло.
- Разработка алгоритма построения СНЭВ с помощью метода Монте-Карло для получения СНЭВМК (состояний с нулевой энергией взаимодействий, полученных методом Монте-Карло).
- Программное воплощение и проверка метода СНЭВМК применительно к различным структурам белков и нуклеиновых кислот.
- Получение с помощью СНЭВМК потенциалов атомарной гидратации (ПАГ) белков и нуклеиновых кислот, основанных на статистике парного взаимодействия различных атомов с молекулами воды в 1776 трёхмерных структурах обучающей выборки.
- Проверка ПАГ в тестах по предсказанию расположения связанной воды в трёхмерных структурах белков
- Использование ПАГ для распознавания нативной укладки белков среди множества альтернативных неправильно уложенных структур – декоев.
Научная новизна и практическая значимость работы.
- Разработан принципиально новый способ задания невзаимодействующих состояний в статистико-механических атомных ансамблях – СНЭВМК. Этот способ учитывает размер и форму пространственной структуры, мономерный состав, число и распределение контактирующих атомов, и позволяет получать распределения ожидаемых межатомных контактных расстояний с любой требуемой точностью, на неограниченном диапазоне контактных расстояний.
- Впервые для статистических межатомных потенциалов создан способ детального количественного исследования атомарного отталкивания на малых расстояниях, сравнимых и меньших, чем Ван дер Ваальсовские радиусы атомов.
- Впервые получены потенциалы атомарной гидратации (ПАГ) белков и нуклеиновых кислот, основанные на статистиках парных взаимодействий различных атомов с молекулами воды в 1776 трёхмерных структурах обучающей выборки.
- Показана способность ПАГ предсказывать, с высокой точностью, расположение связанной воды в белках и белковых комплексах.
- Показана возможность, при помощи вычисления энергии сольватации с использованием ПАГ, различать нативную укладку белка, среди множества неправильно уложенных структур-обманок (декоев).
- Разработан широкий программный инструментарий для количественного анализа белковой сольватации, на уровне отдельных атомов и аминокислот, для визуализации результатов, применяемый при создании новых лекарственных средств и термостабильных форм ферментов.
Апробация работы. Диссертационная работа была представлена на заседании секции молекулярной биологии Ученого Совета ФГУП «ГосНИИ Генетика» 14 марта 2008 г. Материалы исследований по теме диссертации докладывались на межлабораторных семинарах в ГосНИИГенетики (2001-2008), МГУ, ИБХ, СПБГУ, ИМПБ; на международных конференциях и школах: BGRS’2004 (Bioinformatics of Genome Regulation and Structure, Новосибирск, Россия), MCCMB’05, (Moscow Conference on Computational and Molecular Bilogy Москва, Россия), FEBS-2006 (31st Congress of the Federation of European Biochemical Societies, Istanbul, Turkey), MCCMB’07, (Moscow Conference on Computational and Molecular Bilogy Москва, Россия), Erice-2008 (40th International Course in Erice, Italy – From Molecules To Medicine: Integrating Crystallography In Drug Discovery).
Публикации. По материалам диссертации опубликовано 10 печатных работ, включая три статьи, а также материалы докладов научных конференций.
Структура диссертации. Диссертационная работа изложена на 91 странице машинописного текста и состоит из трех основных разделов: введение, с обзором литературы, результаты и обсуждение, и выводов. Раздел «Результаты и обсуждение» состоит из четырех глав, каждая из которых начинается с краткого резюме и содержит описание выполненных автором исследований, изложение полученных результатов и их обсуждение. Список литературы, приведенный в конце диссертации, содержит 82 наименования. Работа содержит 23 рисунка и 3 таблицы.
СОДЕРЖАНИЕ РАБОТЫ
Введение и обзор литературы. Содержит мотивировку поставленных задач, а также аналитический обзор современной литературы по проблемам, рассмотренным в диссертации.
Результаты и обсуждение.
Глава I. Построение распределений ожидаемых плотностей вероятности атомарной гидратации трёхмерных структур белков.
Для построения атомных контактных потенциалов, необходимо получить ожидаемые плотности вероятности межатомных контактов на разных расстояниях, в данном случае – контактов с белок-связанными молекулами воды.
В общепринятой формуле для вычисления статистических потенциалов (1),
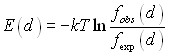 (1)
(1)
здесь значение fexp(d) представляет собой ожидаемую частоту контактов между рассматриваемой парой атомов типов a и b на расстоянии d в так называемом референсном состоянии, т.е. виртуальном состоянии с нулевой энергией взаимодействия (СНЭВ) между атомами данных типов. Наблюдаемая частота контактов fobs(d) определяется непосредственно из измеряемой статистики контактов атомов имеющихся типов в базе данных трёхмерных структур.
Для построения СНЭВ был предложен метод (Rahmanov and Makeev 2004), использующий технику Монте Карло, получивший название СНЭВМК. В качестве невзаимодействующих элементов в нём используются точки со случайными координатами насыщающие всё структурное пространство макромолекулы. Благодаря возможности использовать неограниченное число случайных проб, достигается любая заданная степень точности при вычислении ожидаемых плотностей вероятности парных контактов для любых атомов данной структуры. Также, при нормировке на число случайных точек, естественным образом учитывается размер и форма белковой структуры, а также число и расположение изучаемых контактирующих элементов. Поскольку случайные точки могут располагаться на сколь угодно малых расстояниях от любых элементов структуры, они могут быть использованы для детального количественного исследования атомарного отталкивания на малых расстояниях и расстояниях, сравнимых с ван дер Ваальсовскими радиусами атомов. Это позволило поднять детальность (уменьшить шаг гистограммы) при построении статистических потенциалов на один-два порядка, по сравнению с ранее существующими методами.
Глава II. Получение потенциалов гидратации белковых и ДНК атомов разных типов, из статистики их контактных расстояний с водой в обучающей выборке экспериментально полученных пространственных структур.
Мы применили метод СНЭВМК для построения потенциалов гидратации электронно-плотных (неводородных) белковых и нуклеиново-кислотных атомов различных типов, на основе статистики их непосредственных контактов с кристаллографически определёнными молекулами структурной воды (Rahmanov and Makeev 2006).
В качестве обучающей выборки была использована база данных из 1776 макромолекулярных (преимущественно белковых) трёхмерных структур высокого разрешения, с низкой (не более 25%) парной гомологией (Hobohm and Sander 1994).
Было введено ограничение на величину парной гомологии (по последовательности) между любыми двумя структурами, входящими в обучающую выборку, необходимое для того, чтобы близкие белки, представленные большим числом копий, не искажали среднюю картину атомных взаимодействий, внося в нее особенности, характерные для сильно представленных семейств.
Для каждой струкутры из обучающей выборки, для всех молекул структурной воды, производилась пре-фильтрация по следующему алгоритму: в соответствии с приведёнными в тексте файла структуры операторами пространственных трансформаций симметрии, генерировались симметричные положения молекул воды, принадлежащие соседним ячейкам кристаллографирования. В тех случаях, когда новое положение молекулы оказывалось ближе к белковой части исходной структуры (но при этом без перекрывания ван дер Ваальсовских радиусов атомов), происходило добавление воды в новое положение, с одновременным удалением из исходного. Необходимость данной процедуры объясняется тем, что довольно часто, в рентгеноскопированных структурах наблюдаются изолированные молекулы воды, отстоящие от макромолекулы на 10 ангстрем и более. Иногда, оказывается, что эти молекулы воды связаны с белком из соседней кристаллографической ячейки; в таких случаях вышеописанная процедура приводила к изменению положения соответствующих молекул относительно основной белковой структуры. Когда же вода в итоге не оказывалась ближе к белку, чем пороговое значение (обычно 4.5 ангстрема), она не использовалась в дальнейших вычислениях.
Для каждого типа атомов, из каждой структуры обучающей выборки, вычислялись и сохранялись расстояния до всех атомов кислорода молекул воды, указанных в.pdb-файле структуры. Типы атомов в данной работе определялись с учётом как химической природы, так и группы происхождения. Таким образом, CA_Val обозначает C-атом валина, отличный как от C-атома валина, так и, например, от C-атома тирозина.
Сохранённые контактные расстояния от атомов всех имеющихся типов (за исключением атомов водорода, имеющих очень маленькую электронную плотность, и с трудом определяемых на кристаллограммах белков), до воды, нормализовывались путём деления на ранее определённые величины fexp(d) (см. формулу 1).
После нормировки на число атомов данного типа, получающееся в результате описанной процедуры значения распределения нормированного отношения правдоподобия, как и следовало ожидать, колеблются в районе единицы на больших расстояниях, где взаимодействие практически отсутствует (см. рисунок 1).
На рисунке 1 приведены нормированные отношения правдоподобия для вазимодействия молекул связанной воды в белках между собой. Они показывают плотность распределения частот наблюдения атомов кислорода молекул воды в зависимости от расстояния между ними, относительно аналогичного среднего распределения плотности встречаемости атомов кислорода молекул воды вокруг случайной точки в пространстве структуры. Нормированное отношение правдоподобия на рис. 1 отражает, каким образом молекула связанной воды в белке в среднем влияет вокруг себя на плотность других молекул связанной с белком воды. Значения кривой выше единицы означают положительные контактные предпочтения (вероятности образования пар молекул воды на данном расстоянии), и наоборот. Приведённая на рисунке 1 аналогичная величина для жидкой воды общепринято носит обозначение gOO(r), или кислородно-кислородное радиальное распределение плотности, и может быть извлечена путём анализа экспериментальных данных о дифракции рентгеновских лучей либо измерений рассеяния нейтронных пучков (Head-Gordon and Hura 2002).
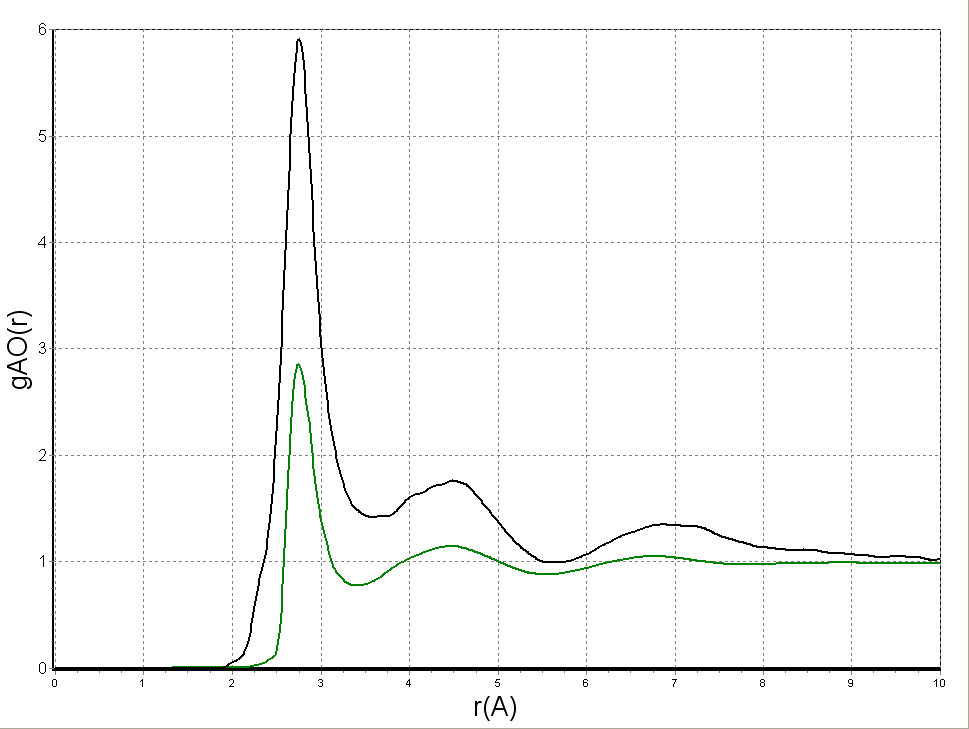
Рисунок 1. Нормированные отношения правдоподобия для взаимодействия атомов кислорода двух соседних молекул связанной воды в белковых кристаллах («водно-водный показатель контактных предпочтений»), вычисленный на основе статистики 54356851 парных контактов между 319024 молекул воды в структурах обучающей выборки. Нижняя кривая соответствует радиальной функции плотности водно-водных контактов (по кислородному атому) в жидкой воде, полученной экспериметально в работе (Head-Gordon and Hura 2002).
Полученная нами кривая усреднённой радиальной функции распределения близко повторяет расположение и относительные величины пиков функции gOO(r). Более высокие уровни распределения плотности водных контактов в кристаллах белков, по-видимому, связаны с наличием исключённого для воды объёма, занятого атомами белка, и с кластеризацией молекул воды преимущественно на поверхности белковой глобулы.
Наблюдаемые три пика контактной плотности при 2.75, 4.5 и ~7 отражают слои структурированной воды, также детектируемые в жидкой воде и во льду. Представляется заслуживаемым обсуждения тот факт, что до сих пор было распространено представление о характере водно-водных взаимодействий в окрестности белков, существенно отличном от классического, наблюдаемого в чистой воде (Makarov and Pettitt 2000). Наши данные (рис. 1) однозначно опровергают это представление.
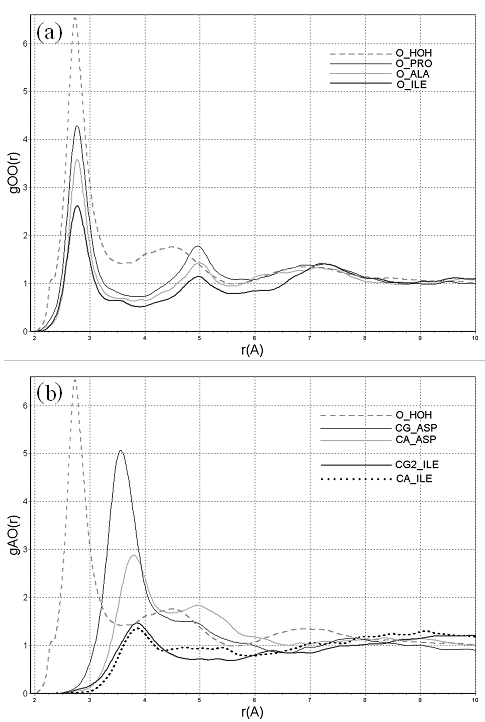
Рисунок 2. Нормированные отношения правдоподобия атомарной гидратации для некоторых белковых кислородных (a) и углеродных (b) атомов.
На рисунке 2 показаны примеры нормированных отношений правдоподобия и потенциалов атомарной гидратации для нескольких типов белковых атомов. Применительно к рис. 2(b) стоит особо отметить наблюдаемый для CG_Asp, углерода в гамма-положении боковой цепи аспартата, сдвиг первого пика потенциала в сторону меньших контактных расстояний, – как вершины, так и фронта. Проявленная в данном случае нехарактерная для атома углерода гидрофильность объясняется тем, что CG_Asp несёт частичный положительный заряд, вследствие оттягивания электронной плотности на сильно электроотрицательные кислородные атомы карбоксильной группы. Благодаря использованию СНЭВМК, количественные характеристики амфифильности всех типов тяжёлых атомов белков были впервые охарактеризованы с высокой детальностью и на всём диапазоне контактных расстояний, включая суб-ван-дер-Ваальсовские.
Рисунок 3. Нормированные отношения правдоподобия атомарной гидратации пар симметричных атомов углерода ароматических боковых цепей тирозина и фенилаланина.
Две пары симметричных атомов углерода ароматических боковых цепей тирозина и фенилаланина на рис. 3 представляют собой своего рода природный тест на величину статистической погрешности статистических потенциалов межатомного взаимодействия в белках: располагающиеся в симметричных положениях, химически идентичные атомы углерода находятся в неотличимом окружении. Атомы каждой пары находятся достаточно далеко друг от друга, чтобы иметь независимую статистику парных контактов с молекулами воды. При этом нет никаких физических причин для того, чтобы они имели различающиеся контактные потенциалы атомарной гидратации. Таким образом, все различия должны быть обусловлены статистической погрешностью процедуры, вследствие, например, ограниченного размера обучающей выборки и т.п. Видно, что их потенциалы, как и следовало ожидать, мало отличаются друг от друга, при этом значимые компоненты потенциала отчётливы на фоне статистических флуктуаций, вызванных погрешностью метода.
Глава III. Тестирование полученных потенциалов атомной гидратации в задачах предсказания расположения связанной воды в белках.
Применимость полученных потенциалов атомарной гидратации была проверена на нескольких практически значимых задачах, первая из которых – задача предсказания расположения молекул связанной (структурной) воды в белках. Нами было показано (Rahmanov and Makeev 2006), что полученные при помощи метода СНЭВМК статистические атомные потенциалы гидратации белков позволяют в несколько раз более точно предсказывать расположение молекул связанной воды в белках, нежели существующие методы, такие как молекулярная динамика и др. Это достигается при во много раз меньшем уровне пере-предсказания, и при этом требует значительно меньших вычислительных ресурсов, чем при использовании альтернативных методов.
Учёт индивидуальных молекул воды часто очень важен для анализа функции белка (Garczarek and Gerwert 2006). Специфическим образом связанные со структурой молекулы воды часто опосредуют взаимодействия макромолекул друг сдругом и с малыми молекулами, включая лиганды - фармакологические агенты (Schneider et al 1995). Известны случаи, когда вытеснение единственной молекулы воды из белок-пептидного интерфейса, вследствие мутации предназначенной увеличить аффинность комплекса, приводило наоборот, к её падению на два порядка. Особенную актуальность задача предсказания расположения индивидуальных молекул воды приобретает в связи с тем, что альтернативные методы моделирования макромолекулярной сольватации, такие, как вычисления переменного диэлектрического континуума путём решения уравнений Пуассона-Боцмана, не позволяют уверенно предсказывать участки гидратации.
Задача определения мест расположения связанной воды в белках решается в несколько этапов. На первом, происходит заполнение пространства гидратационной оболочки макромолекулы мелкоячеистой сеткой (с шагом ~0.1 – 0.4 ангстрема, в зависимости от потребности текущей задачи) из точек-проб, располагающихся в узлах. Гидратационная оболочка определяется здесь как область пространства, все точки которой отстоят от любого из атомов макромолекулы не ближе, чем 2 ангстрема, и не дальше, чем 4.5 ангстрема от хотя бы одного из атомов, принадлежащих структуре. Затем, для всех узлов полученной сетки, вычисляется локальная энергия гидратации, как сумма вкладов - энергий гидратации, от всех близлежащих атомов макромолекулы. На следующем этапе происходит определение подмножества проб, являющихся локальными минимумами гидратационной энергии, в некотором заданном объёме. Использовалось значение 1.4 ангстрема, т.е. половина ван дер Ваальсовского расстояния между соседними атомами кислорода воды. Необходимость данного этапа обусловлена тем, что для ускорения вычислений, необходимо уменьшить число рассматриваемых далее проб, за счёт наименее вероятных участков гидратации, которыми являются точки с большей оцененной энергией гидратации. На следующем, конечном, этапе, происходит уточнение расположения локальных минимумов энергии, за несколько шагов вычислений, по методу сопряжённых градиентов, т.е. путём последовательных сходящихся итераций блуждания проб в трёхмерном пространстве структуры. На каждом шаге, происходит смещение проб в положения, соответствующие меньшим (более оптимальным) значениям оценки свободной гидратации в данной точке. Эта процедура попеременно выполняется либо вдоль линий, соединяющих соседние пробы, обсчитанные на предыдущем шаге (способ сопряжённых градиентов), либо с использованием случайных точек, сгенерированных в заданной малой окрестности каждого локального минимума гидратационной энергии. В первом случае, смещение происходит по пространственному градиенту энергии, вдоль отрезка, соединяющего две близлежащие пробы, на расстояние, пропорциональное различию в их энергии. Новая точка, в которой будет произведена оценка энергии гидратации в контексте окружающей белковой структуры, делит отрезок между двумя соседними пробами, на части, соотносящиеся между собой, как отношение между отклонениями энергии в каждой из вершин отрезка, от текущего среднего значения энергии для всех проб. Во втором случае (с использованием случайных проб в окрестности локального минимума), симулируется процедура т.н. отжига, направленная на избегание «чересчур локальных» минимумов, на достижение действительных полных локальных минимов энергии.
Следует отметить, что исполняемая на данном этапе итеративная процедура является, во-первых, быстро сходящейся (к исчезающее малым смещениям положений локальных минимумов энергии), и, во-вторых, устойчивой к отклонениям как начальных параметров (положений проб и т.п.), так и деталей самой процедуры (величины шага покрытия гидратационной оболочки и т.д.).
Предсказания участков расположения связанной воды были получены для 16 белковых структур, для некоторых из которых были ранее сделаны аналогичные предсказания, с использованием других методов (van Gunsteren, Berendsen et al. 1983), (Henchman and McCammon 2002), (Gelpi et al 2001), (Marrone, Resat et al. 1998).
Эмпирически обнаружено, что в среднем при использовании количества проб, равного числу молекул воды в моделируемой структуре, достаточно хорошо (с ошибкой не более 1.4 ангстрема, т.е. половины расстояния между двумя соседними молекулами воды) воспроизводятся примерно 50% сайтов связанной воды в белках. При использовании вдвое большего количества проб, этот параметр возрастает в среднем до 70-90%. Таким образом, уровень перепредсказания при предсказании сайтов гидратации в белках, в этой работе в среднем равен двум, что существенно (порой в десятки раз) ниже, чем в других опубликованных исследованиях.
Сводная таблица, объединяющая результаты предсказания участков гидратации – мест расположения связанной воды в белке, для 16 структур различных белков, экспериментально полученных с высоким разрешением координат водной и белковой части, приведены в Таблице 1.
Таблица 1. Результаты предсказания участков гидратации – мест расположения связанной воды в белке, для 16 различных белковых структур.
| ## | Белок PDB ID | Амино-кислот | Сайтов гидратации | Среднее RMSD предсказаний | Z-score | |
| эксперимент. | предсказан. | |||||
| 1 | 1AJJ | 37 | 30 | 60 | 1.02 | 6.97 |
| 2 | 1ECA | 136 | 94 | 188 | 1.72 | 4.72 |
| 3 | 1ENH | 54 | 33 | 66 | 0.87 | 5.36 |
| 4 | 1FFO | 202 | 122 | 244 | 0.91 | 9.18 |
| 5 | 1IRD | 287 | 325 | 650 | 1.44 | 2.69 |
| 6 | 1KR7 | 110 | 106 | 212 | 0.86 | 9.60 |
| 7 | 1MAA | 540 | 187 | 374 | 1.32 | 5.41 |
| 8 | 1MOF | 53 | 39 | 78 | 0.90 | 7.34 |
| 9 | 1SBX | 103 | 79 | 158 | 0.86 | 10.41 |
| 10 | 1UOY | 64 | 126 | 252 | 1.03 | 7.73 |
| 11 | 4MT2 | 62 | 69 | 138 | 1.35 | 4.48 |
| 12 | 5PTI | 58 | 63 | 126 | 1.07 | 8.53 |
| 13 | 1IFC | 131 | 206 | 412 | 1.49 | 6.15 |
| 14 | 2PTH | 193 | 175 | 350 | 1.68 | 5.91 |
| 15 | 3EBX | 62 | 82 | 164 | 1.58 | 4.52 |
| 16 | 4PTI | 58 | 57 | 114 | 1.29 | 7.28 |
Для оценки качества предсказания был взят Z-критерий (Z-score), вычисляемый следующим образом:
z = (RMSDrandom - RMSDpredicted) / srandom,
где RMSDpredicted – это квадратный корень из среднего квадратичного отклонения (root mean square deviation) между предсказанными положениями сайтов гидратации, и определёнными экспериментально. RMSDrandom – аналогичная величина, вычисленная несколько раз и усреднённая, для равного числа случайных проб в объёме данной структуры, с наложенными стерическими ограничениями, эквивалентными тем, которые действуют на экспериментально определённые участки гидратации в данной структуре. srandom – величина разброса (дисперсии) отклонений положений случайных проб от равного числа экспериментальных точек гидратации, полученная в вычислительном эксперименте с использованием техники Монте Карло, и также усреднённая для нескольких симуляционных прогонов.
Глава IV. Тестирование полученных потенциалов атомной гидратации в задачах распознавания нативной укладки белка среди множества альтернативных неправильно уложенных структур.
Проблема проверки математических моделей трёхмерной полипептидной структуры требует набора стандартных тестов, которые бы позволили сравнивать результаты предсказания укладки с использованием различных методов. Распространённым тестом для алгоритмов, моделирующих укладку белков, является задача нахождения нативной, экспериментально определённой конформации, среди множества искусственно полученных неправильных структур-«обманок» (decoys). В открытом доступе в интернете находятся несколько наборов белковых структур такого рода, в англоязычной научной литературе носящих название decoy sets, созданных разными группами авторов. Тот факт, что водная среда играет определяющую роль в укладке и стабилизации структуры большинства белков, позволил нам применить атомарные потенциалы гидратации белков для анализа вариантов укладки полипептидной цепи и предсказания нативных конформаций.
Данный тест, т.е. вычисление полной сольватационной энергии структуры, является наиболее естественным применением полученных потенциалов. Важность этой задачи сложно переоценить. Достаточно сказать, что без её решения невозможно адекватное моделирование укладки и функционирования трёхмерной структуры белка, его взаимодействия с другими молекулами клетки. Тот факт, что по некоторым данным до 90% свободной энергии, стабилизирующей нативную конформацию большинства глобулярных белков, обеспечивается гидрофобными взаимодействиями (Makarov, Andrews et al. 2000), (Li X, Liang 2006), говорит о важности методов определения этой энергии.
Применяя потенциалы гидратации белков к задаче распознавания нативной укладки белка, мы использовали наиболее простой критерий средней энергии гидратационной оболочки вокруг структуры. Гидратационная оболочка определяется нами как область пространства вокруг и внутри структуры, все точки которой лежат не далее 4.5 ангстрем и не ближе 2.5 ангстрем от любого не-водородного атома белка. Доля точек, падающих внутрь области ГО, от общего количества проб, сгенерированных в прямоугольнике, описывающем белок (плюс 4.5 ангстрема со всех сторон), позволяет приблизительно определить объём сложной формы ГО. Сольватационная энергия в каждой точке вычислялась так же, как и в предыдущих разделах, т.е как сумма вкладов энергий от всех близлежащих атомов белка.
Основной набор структур-«обманок», использованный для тестирования – это “Improved Rosetta decoy set” (Tsai, Baker et al 2003), является общепризнанно сложным тестом, в связи с большим количеством декоев (в среднем около 1800) для каждой нативной структуры, и повышенной частотой структур-обманок с высоким сходством с нативной структурой.
Полученные нами результаты различения нативных струтур при помощи вычисления свободной энергии сольватации структуры, приведены в таблице 2.
Таблица 2. Распознавание нативной укладки белка среди множества структур-обманок, при помощи вычисления полной энергии гидратации структуры, с использованием набора структур «Rosetta».
| # | Код PDB структуры | Средняя энергия гидратации структуры, kT | Отличие нативной структуры, G, Z-крит. | Ранг нативной структуры, % | |
| нативная | декои | ||||
| 1 | 1res | -11.30 | -9.63 | 5.10 | 100 |
| 2 | 1ptq | -10.21 | -9.70 | 0.68 | 74 |
| 3 | 1uxd | -10.04 | -8.27 | 5.08 | 100 |
| 4 | 2pdd | -8.68 | -8.73 | 0.04 | 51 |
| 5 | 1uba | -9.01 | -8.28 | 1.24 | 88 |
| 6 | 1gab | -7.71 | -5.98 | 3.25 | 100 |
| 7 | 1vif | -10.05 | -8.70 | 2.37 | 98.5 |
| 8 | 1bq9 | -10.22 | -8.01 | 3.55 | 100 |
| 9 | 5pti | -11.12 | -8.31 | 4.70 | 100 |
| 10 | 1aa3 | -9.94 | -7.76 | 3.78 | 100 |
| 11 | 1bw6 | -7.962 | -7.564 | 0.80 | 79 |
| 12 | 1orc | -8.944 | -9.693 | -1.04 | 15 |
| 13 | 1am3 | -10.857 | -9.126 | 3.67 | 100 |
| 14 | 1pgx | -10.211 | -8.262 | 3.40 | 100 |
| 15 | 1tif | -10.283 | -8.416 | 3.33 | 100 |
| 16 | 1msi | -12.712 | -10.401 | 3.69 | 100 |
| 17 | 2ptl | -10.982 | -7.708 | 6.89 | 100 |
| 18 | 1r69 | -10.316 | -7.670 | 4.69 | 100 |
| 19 | 1tuc | -10.564 | -8.614 | 2.28 | 100 |
| 20 | 1dol | -10.890 | -8.611 | 3.68 | 100 |
| 21 | 1utg | -7.998 | -7.792 | 0.42 | 67 |
| 22 | 1csp | -11.524 | -8.227 | 4.30 | 100 |
| 23 | 1a32 | -9.844 | -10.077 | -0.50 | 28 |
| 24 | 2ezh | -11.496 | -9.431 | 3.74 | 100 |
| 25 | 1nre | -8.198 | -7.288 | 1.92 | 97 |
| 26 | 1sro | -11.319 | -9.443 | 2.85 | 99.5 |
| 27 | 2fow | -9.442 | -7.466 | 5.24 | 100 |
| 28 | 1ail | -10.957 | -10.734 | 0.38 | 63 |
| 29 | 1ctf | -8.758 | -7.123 | 3.30 | 100 |
| 30 | 1lfb | -9.387 | -7.908 | 3.18 | 100 |
| 31 | 1nkl | -11.289 | -10.048 | 2.21 | 99 |
| 32 | 1pou | -11.890 | -9.495 | 4.40 | 100 |
| 33 | 1mzm | -12.126 | -9.837 | 4.93 | 100 |
| 34 | 1afi | -10.691 | -9.655 | 2.28 | 99.5 |
| 35 | 5icb | -9.006 | -6.895 | 5.80 | 100 |
| 36 | 1kjs | -10.521 | -9.063 | 2.90 | 100 |
| 37 | 1hyp | -14.662 | -12.513 | 2.63 | 98.5 |
| 38 | 1cc5 | -10.227 | -9.895 | 0.64 | 77 |
| 39 | 1vcc | -13.279 | -11.533 | 2.71 | 100 |
| 40 | 2fxb | -11.126 | -8.699 | 4.37 | 100 |
| 41 | 1cei | -9.810 | -9.589 | 0.43 | 65 |
Пятый столбец в Таблице 2 приводит различие в энергии гидратации между нативной структурой и средним для структур-обманок по Z-критерию (см. ниже). Последний столбец содержит величины доли декоев с оценкой свободной энергии выше (менее оптимальной) чем нативная структура.
Для оценки качества предсказания был, как и ранее, использован Z-критерий, вычисляемый как:
z = (Edecoy – Enative) / sdecoy,
где Edecoy – это вычисленная средняя энергия сольватации для набора декоев, Enative – энергия нативной структуры, sdecoy – величина разброса (дисперсии) энергий декоев.
Из таблицы 2 видно, что применение ПАГ позволяет различать нативную структуру белков среди множества структур-обманок. Для 26 из 41 структур из набора декоев «Rosetta», нативная структура оказалась абсолютно лучшей по оцененной средней энергии сольватации. Такие показатели дискриминации находятся на уровне лучших опубликованных систем моделирования структур белков. Полученный результат достигнут даже без учёта внутренних взаимодействий в белковой макромолекуле, только за счёт оценки энергии взаимодействия белка с водой. Среди тех нативных структур, которые не были распознаны при помощи ПАГ в данном тесте, большинство заведомо не годятся для применения ПАГ. Так, например, белок 1utg имеет форму полусферы, вогнутая внутренняя поверхность которой образована практически полностью гидрофобными аминокислотами. Представляется невероятным, чтобы подобная структура могла существовать как нативная в водной среде. В действительности, 1utg существует в виде димера, в котором гидрофобные стороны плотно состыкованы. Другая нераспознанная структура, рибосомальный белок 1a32, представляет собой некомпактную, штопорообразную структуру, очевидно стабилизируемую в первую очередь контактами с множеством других рибосомальных белков и РНК, а не с растворителем.
Успешное использование ПАГ для выделения правильной конформации полипептида среди множества неправильных конформаций, предполагает, что различные варианты укладки белка, могут быть количественно проранжированы. При этом каждой конформации, схожей с нативной, сопоставляется некоторая вероятность её возникновения, что в некоторых случаях может быть проинтерпретировано, как доля времени которую данный полипептид проводит в рамках данной конформации. Полученные результаты позволяют надеяться, в перспективе, на построение динамической модели белковой структуры, в том числе и описывающей её взаимодействие с другими макромолекулами и малыми молекулами клетки.
ВЫВОДЫ.
- Разработан программный комплекс для анализа водной сольватации биологических макромолекул на основе вычисления свободной энергии атомарной гидратации.
- Разработан метод стохастического моделирования для построения состояния невзаимодействующих атомов при определении статистических потенциалов межатомного взаимодействия.
- При помощи нового метода получены потенциалы атомной гидратации белковых макромолекул, обладающие высокой разрешающей способностью и широким контактным диапазоном.
- Продемонстрирована возможность использования потенциалов гидратации для предсказания расположения связанной воды в белках.
- Продемонстрирована эффективность использования полученных потенциалов для распознавания нативной укладки белка среди набора альтернативных конформаций.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ.
Статьи в научных журналах:
- С.В. Рахманов и В.Ю. Макеев. Использование невзаимодействующих проб в пространстве белковой структуры для построения статистических потенциалов межатомного взаимодействиях. Биофизика, 2008, том 53, вып. 3, с. 389-396.
- Rakhmanov S.V., Makeev V.J. Atomic hydration potentials using a Monte Carlo Reference State (MCRS) for protein solvation modeling. BMC Structural Biology 2007, 7:19.
- S.V. Rahmanov, V.J. Makeev. Atomic hydration potentials using Monte Carlo reference state advance protein solvation modeling. FEBS Journal 2006, 273 (s1), 62-64.
Материалы научных конференций:
- Modeling of protein crystal hydration in topologically closed asymmetric unit using 3D motif empirical atomic potentials. Sergei V. Rahmanov and VsevolodYu. Makeev, 40th Erice Crystallographic Conference “From Molecules to Medicine: Integrating Crystallography in Drug Discovery”, Erice, Italy 2008.
- Knowledge-based potentials for protein atom interaction based on Monte Carlo reference state. Sergei V. Rahmanov and Vsevolod J. Makeev. Moscow Conference on Computational Molecular Biology MCCMB-2007. Moscow 2007.
- Статистические потенциалы взаимодействия белковых атомов. В.Ю. Макеев, С.В. Рахманов. IV Московский международный конгресс «Биотехнология: состояние и перспективы развития». (Мир БиоТехнологии-2007).
- Atomic hydration potentials using Monte Carlo reference state advance protein solvation modeling. Sergei V. Rahmanov and Vsevolod J. Makeev. FEBS-2006, Istanbul, Turkey.
- Энергетические потенциалы гидратации белков. Рахманов С.В., Макеев В.Ю. Российская школа - конференция «Генетика микроорганизмов и биотехнология», посвящённая 100-летию со дня рождения С. И. Алиханяна. Москва 2006.
- Новый метод построения межатомных потенциалов для моделирования структуры и взаимодействий биополимеров. Рахманов С.В. Международная школа-конференции молодых ученых «Системная биология и биоинженерия» Звенигород, 2005.
- Constructing detailed knowledge-based atomic potentials for water in proteins. Rahmanov S.V., Makeev V.Yu., The Fourth International Conference on Bioinformatics of Genome Regulation and Structure (BGRS'2004), Novosibirsk, 2004.