

Биологическая активность эндофитных штаммов bacillus subtilis, перспективных в качестве основы новых препаратов для растениеводства
На правах рукописи
Егоршина Анна Александровна
БИОЛОГИЧЕСКАЯ АКТИВНОСТЬ ЭНДОФИТНЫХ ШТАММОВ BACILLUS SUBTILIS, ПЕРСПЕКТИВНЫХ В КАЧЕСТВЕ ОСНОВЫ НОВЫХ ПРЕПАРАТОВ ДЛЯ РАСТЕНИЕВОДСТВА
03.02.03 – микробиология
03.01.06 – биотехнология (в том числе бионанотехнологии)
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Саратов – 2012
| Работа выполнена в ФГБОУ ВПО Башкирский государственный аграрный университет и Федеральном государственном бюджетном учреждении науки Институте биохимии и генетики Уфимского научного центра Российской академии наук |
| Научный руководитель: | доктор биологических наук, профессор Хайруллин Рамиль Магзинурович, Федеральное государственное бюджетное учреждение науки Институт биохимии и генетики Уфимского научного центра Российской академии наук |
| Официальные оппоненты: | доктор биологических наук Антонюк Людмила Петровна, Федеральное государственное бюджетное учреждение науки Институт биохимии и физиологии растений и микроорганизмов Российской академии наук кандидат биологических наук Актуганов Глеб Эдуардович, Федеральное государственное бюджетное учреждение науки Институт биологии Уфимского научного центра Российской академии наук |
| Ведущая организация | Федеральное государственное бюджетное учреждение науки Казанский институт биохимии и биофизики Казанского научного центра Российской академии наук |
Защита диссертации состоится "4" апреля 2012 г. в 14:00 ч на заседании диссертационного совета Д 002.146.01 при Институте биохимии и физиологии растений и микроорганизмов РАН (410049, г. Саратов, просп. Энтузиастов, 13).
Тел. / факс (8452) 970383.
Автореферат диссертации размещен на официальном сайте Минобрнауки РФ и на сайте ИБФРМ РАН: http://ibppm.ru/dissertacionnyy-sovet/
С диссертацией можно ознакомиться в научной библиотеке ИБФРМ РАН.
Автореферат разослан "2" марта 2012 г.
Ученый секретарь диссертационного совета
доктор биологических наук, профессор В.Е. Никитина
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Долгое время единственными непатогенными для растений микроорганизмами, способными обитать внутри растительных тканей, считались клубеньковые бактерии и эндомикоризные грибы. Многочисленные доказательства способности других видов бактерий заселять внутренние растительные ткани, не вызывая какие-либо симптомы у растений (Sturz, Nowak, 2000; Bacon, Hinton, 2002; Antoun, Prvost, 2005; Wu et al., 2009), изменили традиционное мнение о стерильности последних, что позволило сформировать представление о растениях, как сложных микроэкосистемах, являющихся местом обитания различных эндофитных микроорганизмов, как паразитов, так и мутуалистов. Последние привлекают особое внимание исследователей, и именно их, как правило, называют эндофитами, исключая паразитов и комменсалов (Antoun, Prvost, 2005; Hardoim et al., 2008). Научный интерес представляют механизмы взаимодействия растений с их эндофитами. Одновременно изучение эндофитов интересно и практикам, так как такие микроорганизмы, как стимуляторы роста или защитные агенты, открывают новые возможности для создания безопасных биопрепаратов. Недостаточные сведения о свойствах эндофитных бактерий и их взаимоотношениях с хозяевами не позволяет успешно создавать на их основе новые более эффективные биофунгициды, что подтверждается использованием в растениеводстве России пока лишь одного препарата «Фитоспорин-М» на основе эндофитного штамма Bacillus subtilis 26Д (Государственный каталог пестицидов…, 2010). При этом многие авторы относят эндофитов к более перспективным бактериям по сравнению с обитателями ризосферы, поскольку первые лучше защищены от неблагоприятного воздействия многих средовых факторов (Менликиев и др., 1996; Недорезков, 2003; Мубинов, 2007). Бациллам, как потенциальной основе биофунгицидов, посвящено значительное число работ (Смирнов и др., 1982; Недорезков, 2003 Мелентьев, 2007; Choudhary, Johri, 2009; Prez-Garca et al., 2010). Однако в большинстве из них эти бактерии рассматривают, в основном, как антагонистов для биоконтроля фитопатогенов, в то время как другим механизмам стимуляции роста, устойчивости и продуктивности растений, особенно эндофитными представителями рода Bacillus, на наш взгляд, уделяется недостаточное внимание.
Цель работы – выявить комплекс биологической активности эндофитных штаммов Bacillus subtilis, позволяющий использовать бактерии в качестве основы новых полифункциональных препаратов для растениеводства.
Задачи работы. 1) Провести сравнительные исследования культурально-морфологических и физиолого-биохимических свойств, определить генетическую гетерогенность новых эндофитных штаммов B. subtilis, выделенных из растений яровой пшеницы. 2) Определить эндофитность одного из перспективных штаммов – B. subtilis 11ВМ с помощью рекомбинантного штамма B. subtilis 11ВМgfp. 3) Выявить участие фитогормонов в механизмах регуляции роста растений эндофитными представителями B. subtilis. 4) Изучить способность штаммов переводить труднорастворимые фосфаты в подвижные формы и определить некоторые её механизмы. 5) Охарактеризовать взаимоотношения исследуемых штаммов B. subtilis с фитопатогенными грибами, а также азотфиксирующими бактериями. 6) Оценить безопасность и технологичность штамма B. subtilis 11ВМ как перспективной основы для разработки полифункционального биофунгицида.
Научная новизна. Дана подробная сравнительная характеристика культурально-морфологических, физиолого-биохимических свойств семнадцати новых эндофитных штаммов B. subtilis, выделенных из растений мягкой яровой пшеницы, показана их генетическая гетерогенность. Впервые показано, что вовлечение фитогормонов – как активаторов (ИУК и цитокинины), так ингибиторов роста (АБК) во взаимоотношения эндофитных штаммов B. subtilis с растениями может быть одним из общих механизмов проявления этими бактериями рострегулирующей активности по отношению к хозяину. Выявлен один из механизмов повышения содержания ИУК в корнях растений под влиянием эндофитных представителей B. subtilis, включающий снижение активности ИУК-оксидазы. Впервые показано, что стимуляция роста растений эндофитными штаммами B. subtilis может быть опосредована их способностью переводить труднорастворимые фосфаты в подвижные формы за счет продукции бактериями органических кислот, спектр которых зависит от биологических особенностей штамма. Показано, что при наличии у изученных штаммов бактерий общего свойства проявлять антагонизм к фитопатогенным грибам рода Fusarium, эти штаммы различаются по способности угнетать рост хозяйственно полезных азотфиксирующих бактерий. Установлено, что отличительным признаком новых штаммов бацилл от депонированных (B. subtilis 26Д, 24D, 11В, М1) является преимущественное ингибирование роста Azospirillum brasilense и A. lipoferum. Показано, что бактерии Azotobacter vinelandii отличаются от других изученных представителей азотфиксаторов устойчивостью к действию антибиотиков всех исследованных штаммов бацилл.
Практическая значимость. Подробно охарактеризованная коллекция из 17 эндофитных штаммов B. subtilis представляет практическую ценность, как для выявления новых свойств этих бактерий, так и разработок биопрепаратов для сельского хозяйства. Штамм бактерий B. subtilis 11ВМ, трансформированный плазмидой pRL-GFP765 может быть использован для изучения механизмов взаимоотношений растений и эндофитов. Показано, что выявление фосфат-мобилизующей активности бактерий и её регистрируемый уровень зависят от консистенции среды, на которой культивируются микроорганизмы, также как и спектр продуцируемых ими органических кислот. Для повышения надежности оценки рострегулирующей активности эндофитных бактерий предложена методика, в которой тест-объекты получают из растений, инокулированных эндофитами. В коллекции ВНИИСХМ депонирован штамм B. subtilis 11ВМ (№519) перспективный в качестве основы нового полифункционального биофунгицида.
Связь работы с научными программами. Исследования по теме диссертации поддержаны грантами РФФИ №05-04-08156-офи_а, №05-04-97935-р_агидель_а, №11-04-97044-р_поволжье_а, Фонда содействию развитию МПНТС №4822р/7290 («СТАРТ-07»), №5490р/7971 («У.М.Н.И.К.»). Часть работы выполнена в рамках заказа Минсельхоза России в соответствии с планом НИР Башкирского ГАУ за счет средств федерального бюджета по теме «Разработка полифункциональных биофунгицидов для растениеводства» (№ госрегистрации 01200853490).
Основные положения, выносимые на защиту. 1. 17 новых эндофитных штаммов отличаются друг от друга по ряду свойств и относятся к виду B. subtilis. 2. Штамм B. subtilis 11ВМ способен проникать во внутренние растительные ткани, не проявляя патогенности по отношению к растению-хозяину. 3. Стимуляция роста растений пшеницы эндофитными штаммами B. subtilis опосредована комплексом их биологической активности: способностью проявлять фитогормон-подобные свойства по отношению к растениям, изменять баланс фитормонов в растительных тканях, переводить труднорастворимые фосфаты в подвижную форму, ингибировать рост фитопатогенных грибов, повышать устойчивость растений к действию фитотоксинов, избирательно подавлять рост азотфиксирующих бактерий. 4. Штамм B. subtilis 11ВМ (ВНИИСХМ №519) безопасен для животных, технологичен и может быть использован в качестве основы полифункционального биофунгицида.
Апробация работы. Основные результаты работы были представлены на: IV Межрегиональной конференции молодых ученых «Стратегия взаимодействия микроорганизмов и растений с окружающей средой» (Саратов, 2008); V международном конгрессе «Биотехнология: состояние и перспективы развития» (Москва, 2009); XII съезде общества микробиологов Украины им. С.М. Виноградского (Ужгород, 2009); II Всероссийской школе-конференции молодых ученых УНЦ РАН и Волго-Уральского региона по физико-химической биологии и биотехнологии «Биомика – наука XXI века» (Уфа, 2011).
Публикации. По теме диссертации опубликовано 11 работ, из них 3 статьи в журналах, рекомендованных ВАК РФ.
Структура и объем диссертации. Диссертация состоит из введения, обзора литературы, экспериментальной части, включающей описание материалов и методов исследования, а также полученных результатов и их обсуждения, заключения, выводов, практических рекомендаций и библиографического списка из 283 наименований, в том числе 249 на иностранном языке. Работа изложена на 145 страницах машинописного текста, включает 8 рисунков и 16 таблиц, а также приложение на 16 страницах, содержащее 5 таблиц.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
Объектами исследования были 23 штамма B. subtilis Cohn. Штаммы 26Д, 24D, М1 (деп. в коллекции ВНИИСХМ, №128, №129, №111Д24 соотв.), 11В (коллекция ИБФМ РАН, №ВКМ В-2218Д) были получены из коллекции ДП «Биофаг» ГУП «Иммунопрепарат» (г. Уфа), остальные 19 (НТ, НТ2, 11ВМ, 11РН, 49РН, 89РН, 112РН, 118РН, 121РН, 122РН, 141РН, 161РН, 162РН, 171РН, 811РН, 832РН, 871РН, 922РН, 962РН) выделены в лаборатории биотехнологии Башкирского ГАУ из поверхностно стерилизованных тканей растений пшеницы (Хайруллин Р.М. и др., 2009). Бактерии культивировали на мясо-пептонном агаре/бульоне (МПА/Б), картофельно-глюкозном агаре (КГА) и жидкой полусинтетической среде (ПСС) (Недорезков, 2003). Свойства штаммов характеризовали согласно определителю Берджи (1998). Молекулярно-генетические различия штаммов оценивали методом RAPD, видовую принадлежность – ПДРФ-анализом гена 16S рРНК. Плазмиду pRL-GFP765 с генами зеленого флуоресцентного белка и устойчивости к канамицину в клетки бактерий вводили электропорацией протопластов (Romero et al., 2006) на приборе MicroPulser (Bio-Rad, США). Для оценки ростстимулирующей активности семена мягкой пшеницы (Triticum aestivum L.) сорта Казахстанская 10, капусты (Brassica oleracea L.) сорта Грибовская обрабатывали суспензией (20 л препарата на 1 т семян) спор бактерий. Через 24 ч семена проращивали 3 дня на влажной фильтровальной бумаге в темноте при 24-25°С и затем измеряли размеры проростков. Фитогормон-подобную активность штаммов определяли с помощью биотестов на ауксин- (Бояркин, 1966), цитокинин- (Кулаева, 1973) и гиббереллин-подобную (Агнистикова, 1966) активности регуляторов роста растений. Экстракцию АБК, ИУК и цитокининов из проростков, оценку концентрации фитогормонов непрямым конкурентным иммуноферментным анализом (ИФА) проводили согласно методам, предложенным Г.Р. Кудояровой с соавт. (Кудоярова и др., 1990) и Ф.М. Шакировой с соавт. (Shakirova et al., 2003), с использованием кроличьих антител, специфичных к АБК, ИУК и зеатинрибозиду. Активность ИУК-оксидазы в корнях пшеницы определяли по методу Гамбурга (1966). Мобилизацию бактериями нерастворимых неорганических фосфатов выявляли на плотной среде Муромцева по образованию зон гало (Сэги, 1983) и в жидкой среде NBRIP (Mehta, Nautiyal, 2001), оценивая количество освобожденного фосфора фотоколориметрически. рН плотной среды NBRIP оценивали по изменению окраски бромфенолового синего (Mehta, Nautiyal, 2001). Фосфатазную активность выявляли по методу Payne и Jackson (1991). Спектр органических кислот определяли ВЭЖХ на хроматографе Waters Breeze (Waters, США), используя колонку Luna C18 (Phenomenex, США) и набор органических кислот фирмы Supelco (США). Антагонизм штаммов B. subtilis к грибам и бактериям оценивали методом агаровых блоков. В качестве тест-объектов использовались фитопатогены Fusarium sporotrichioides (Sherb.) и F. poae (Peck.) Wollenw. (выделены и идентифицированы в лаборатории биотехнологии Башкирского ГАУ) и бактерии-азотфиксаторы Azospirillum brasilense SR15, A. lipoferum Sp59b, A. irakense KBC1 (из коллекции ИБФРМ РАН, г. Саратов), Azotobacter chroococcum B1616, A. vinelandii ИБ-1 (получены из коллекции микроорганизмов ИБ УНЦ РАН, г. Уфа) и Rhizobium leguminosarum bv. viciae 1078 (из коллекции ВНИИСХМ). Штамм B. subtilis 11ВМ в газо-вихревом биореакторе «БИОК» культивировали при условиях, описанных М.А. Лукьянцевым (2010). Полевые опыты проводили в Уфимском районе Башкортостана по методике госсортоиспытания полевых культур. Исследования фитотоксичности метаболитов грибов рода Fusarium проведены совместно с к.б.н. Кутлубердиной Д.Р., полевые опыты – совместно с сотрудниками Башкирского ГАУ к.б.н. Уразбахтиной Н.А. и м.н.с. Мининой Т.С. Автор выражает благодарности д.б.н., проф. Турковской О.В. (ИБФРМ РАН, г. Саратов) и д.б.н., проф. Логинову О.Н. (ИБ УНЦ РАН, г. Уфа) за предоставленные штаммы микроорганизмов, к.б.н. Баймиеву Ан.Х. (ИБГ УНЦ РАН, г. Уфа) за помощь при проведении молекулярно-генетического анализа бактерий и трансформации штамма.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Регуляции роста растений эндофитами
С целью разработки новых полифункциональных препаратов для растениеводства сотрудниками лаборатории биотехнологии Башкирского ГАУ был проведен поиск эндофитных штаммов бактерий, обитающих во внутренних тканях растений пшеницы. Основным критерием отбора служил антагонизм к широкому спектру фитопатогенных грибов. В результате было выделено 19 новых штаммов, предварительно отнесенных к роду Bacillus. Полевые испытания препаратов на основе самых сильных антагонистов показали, что этот критерий, является важным, но не достаточным для отбора перспективных бактерий, способных эффективно повышать урожайность зерна (Хайруллин и др., 2009). Этот вывод подтвержден также исследованиями М.А. Лукьянцева (2010). В связи с этим мы оценили способность штаммов стимулировать рост проростков с.-х. культур. Многие из эндофитных штаммов B. subtilis стимулировали рост проростков пшеницы и капусты (табл. 1, из 23 штаммов приведены наиболее активные стимуляторы роста корней пшеницы). Стимулирующая активность бактерий зависела от концентрации спор. Больший эффект наблюдался при обработке семян бациллами в концентрации 108 КОЕ/мл. При увеличении концентрации в 10 раз ростстимулирующая активность по отношению к корням, например, пшеницы не проявлялась, а в случае таких штаммов, как 49РН, 89РН, 118РН, 161РН, 162РН, 112РН рост корней и/или побегов угнетался. Проростки капусты были менее чувствительны к изменению концентрации спор в препарате, при этом ни один из исследованных штаммов не ингибировал рост корней. Наибольшую стимулирующую активность проявили штаммы 11ВМ, 11В, 121РН, 922РН.
На основании полученных результатов и других лабораторных испытаний штамм 11ВМ был предложен в качестве перспективного в качестве основы биофунгицида для защиты растений.
Таблица 1 – Влияние обработки семян некоторыми эндофитными штаммами
B. subtilis на рост проростков с.-х. культур
| Штамм | Длина проростков, мм | |||||
| 108 КОЕ/мл | 109 КОЕ/мл | |||||
| Пшеница | Капуста, корень | Пшеница | Капуста, корень | |||
| корень | побег | корень | побег | |||
| Контроль | 45,8±3,0 | 12,3±1,1 | 16,8±1,2 | 45,8±2,8 | 12,3±0,5 | 16,8±0,9 |
| 11ВМ | 60,0±5,4 | 18,3±0,8 | 21,8±1,3 | 47,8±4,7 | 18,5±0,9 | 22,9±1,9 |
| 11В | 52,7±3,8 | 16,2±1,2 | 21,1±2,0 | 46,2±3,3 | 15,6±0,8 | 21,3±1,1 |
| 121РН | 55,9±2,6 | 19,0±0,8 | 23,4±1,0 | 45,7±2,2 | 16,4±1,4 | 24,9±1,7 |
| 922РН | 55,5±3,5 | 17,2±1,0 | 24,2±1,5 | 45,8±3,7 | 16,1±1,2 | 19,8±1,7 |
| 26Д (эталон) | 51,4±3,7 | 14,4±0,9 | 23,1±0,7 | 46,3±3,8 | 14,4±0,9 | 22,9±1,1 |
В полевых испытаниях 2007 г выяснилось, что штамм 11ВМ способен защищать растения от корневых гнилей и повышать их урожайность (табл. 2), не уступая по этим показателям биофунгициду «Фитоспорин-М». Следует особо подчеркнуть, что этот препарат кроме клеток штамма B. subtilis 26Д содержит регулятор роста растений в виде гумата, а также микроэлементы. Таким образом, опытный препарат, содержащий только споры B. subtilis 11ВМ проявлял такую же активность, как и комплексный коммерческий препарат.
Таблица 2 – Влияние обработки семян штаммом B. subtilis 11ВМ на поражение пшеницы корневыми гнилями и её урожайность (Уфимский район, 2007 г)
| Вариант | Корневые гнили, % | Урожайность зерна, г/м2 | |
| Распространение | Развитие | ||
| Контроль (вода) | 13±4 | 31±8 | 243±6 |
| B. subtilis 11ВМ (1 л/т) | 6±2 | 17±3 | 286±13 |
| Фитоспорин-М | 5±1 | 15±2 | 274±19 |
Позже, в 2008 г, Иванчиной Н.В. (2010) было выявлено также, что штамм B. subtilis 11ВМ при обработке семян гороха способен снижать поражение растений корневыми гнилями, стимулировать развитие симбиотического аппарата при совместной обработке с Rh. leguminosarum bv. viceae, повышать урожайность зерна гороха. В связи с этим встала задача анализа биологической активности этого штамма в сравнении с «эталонным» 26Д (основа коммерческого препарата "Фитоспорин-М"), а также другими эндофитными штаммами B. subtilis.
Культурально-морфологические, физиолого-биохимические признаки и генетическая гетерогенность новых эндофитных штаммов B. subtilis, выделенных из растений пшеницы
Колонии новых штаммов, растущие на МПА, имели как общие морфологические признаки, так и отличающие некоторых из них. Большинство штаммов формировали плоские непрозрачные колонии, белого или бежевого цвета, с фестончатым краем, с шероховатой поверхностью, матовые, имеющие не выделяющийся центр, зернистой структуры. Некоторые штаммы различались по способности утилизировать лактозу и мочевину, наличию липолитической активности и другим свойствам. Генетический полиморфизм между штаммами четко выявлялся методом RAPD-анализа с двумя праймерами Opa1 и Lmbd8, при этом большинство новых штаммов отличались от «эталонного».
По совокупности свойств все 23 штамма различались между собой. По изученным физиолого-биохимическим свойствам новые штаммы отнесены к виду B. subtilis, что подтвердилось также данными ПДРФ-анализа (рис. 1), в ходе которого показано полное совпадение рестриктазных профилей, полученных с рестриктазами AluI и RsaI, новых штаммов с депонированными.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
 |
Рисунок 1. Электрофореграмма рестрикционных фрагментов гена 16S рРНК эндофитных штаммов B. subtilis с использованием рестриктазы AluI: 1 – 26Д, 2 – НТ, 3 – НТ2, 4 – М1, 5 – 11В, 6 – 11РН, 7 – 49РН, 8 – 89РН, 9 – 118РН, 10 – 24D, 11 – 112PH, 12 – 121PH, 13 – 122PH, 14 – 141PH, 15 – 171PH, 16 – 922PH, 17 – 962PH, 18 – 161PH, 19 – 162PH, 20 – 832PH, 21 – 811PH, 22 – 871PH, 23 – 11ВМ
Эндофитность штамма B. subtilis 11ВМ
Все новые штаммы были отнесены к эндофитным, исходя из выделения бактерий из тканей тщательно поверхностно стерилизованных органов. Для детализации возможных механизмов взаимоотношений растений с эффективным штаммом B. subtilis 11ВМ нужны были более веские доказательства. Поэтому бактерии трансформировали плазмидой pRL-GFP765, содержащей гены устойчивости к канамицину и зеленого флуоресцентного белка (gfp), в результате чего получили штамм B. subtilis 11ВМgfp. Прямая микроскопия тканей растений не позволила нам выявить локализацию бактерий из-за автофлуоресценции растительных клеток. Поэтому в асептических условиях были проведены следующие эксперименты. На корни проклюнувшихся поверхностно стерилизованных семян кукурузы, фасоли, гороха и тыквы наносили суспензию суточной культуры штамма B. subtilis 11ВМgfp и инкубировали ночь. Семена переносили в стаканы и проращивали в темноте при 25°С в течение трех (кукуруза, горох) или пяти (тыква, фасоль) суток на фильтровальной бумаге. После этого проростки тщательно промывали в стерильной воде и последовательно стерилизовали 70%-ным этанолом (10 мин), 6%-ной перекисью водорода (10 мин) и 0,1%-ным раствором нитрата серебра (3 мин), затем трижды промывали стерильной водой, отделяли побег и корень и разрезали их поперек на кусочки длиной 5 мм. После этого кусочки выдерживали 5 мин в чашках Петри №1 на МПА с канамицином (контроль стерилизации) и переносили их на аналогичную среду в чашку Петри №2, проверяя, таким образом, степень поверхностной стерилизации (если бы в чашке №1 вырастали колонии траснформированных бактерий, то поверхность кусочков была бы не стерильной). Через 2-4 дня вокруг всех кусочков, полученных из инокулированных штаммом B. subtilis 11ВМgfp растений на поверхности МПА чашки №2 появлялись колонии флуоресцирующих клеток эндофита. В чашке №1 ни в одном из случаев не было роста бактерий. Таким образом, можно объективно говорить об эндофитности исследованного штамма и отсутствии у него строгой специфичности к хозяину.
Некоторые механизмы стимуляция роста растений эндофитами
Нами выявлено, что одним из механизмов стимуляции роста проростков эндофитами является наличие у бактерий фитогормон-подобной активности, которая проявлялась у штаммов в зависимости от способа постановки эксперимента. Ауксин- и цитокинин-подобная активность выявлялись только при предобработке бактериями семян растений, из которых получали объекты биотестов, тогда как гиббереллин-подобная активность наблюдалась при обработке клетками штаммов непосредственно объектов биотеста (гипокотилей салата) (табл.3). Интересно, что у большинства штаммов рострегулирующая активность проявлялась в биотестах с объектами, в тканях которых, вероятно, уже находились клетки бактерий. Таким образом, внедрение эндофитов в растение могло способствовать усилению их регуляторного действия через клетки хозяина.
Таблица 3 – Фитогормон-подобная активность эндофитов
| Вариант | Активность | ||
| ауксин-подоб-ная, длина от-резков коле-оптилей пшеницы | цитокинин-подобая, содержание хлорофилла в отрезках листьев ячменя | гиббереллин-подобная, длина гипокотилей салата | |
| Вода | 100% | 100% | 100% |
| Фитогормон | ИУК, 132% | БАП, 260% | ГК3, 194% |
| Объект биотеста получен из необработанных семян | |||
| B. subtilis 11ВМ | 104% | 100% | 118% |
| B. subtilis 121РН | 105% | 92% | 107% |
| B. subtilis 26Д | 106% | 98% | 110% |
| Объект биотеста получен из проростков, предобработанных спорами эндофитов | |||
| B. subtilis 11ВМ | 119% | 135% | Не определяли |
| B. subtilis 121РН | 108% | 141% | |
| B. subtilis 26Д | 111% | 105% | |
ИФА содержания АБК, ИУК и цитокининов в проростках, выращенных из семян, обработанных спорами B. subtilis 11ВМ, свидетельствует об участии фитогормонов во взаимоотношениях между эндофитами и растением. Обработка семян бактериями вызывала стойкое увеличение уровня АБК как в корнях, так и в побегах, причем в побегах уже на вторые сутки (рис. 2).
Концентрация ауксинов в инокулированных эндофитом проростках постепенно возрастала так, что в корнях таких растений количество ИУК на пятые сутки было все еще больше, чем в контрольных, в то время как в побегах оно выравнивалось с показателями контроля. Содержание цитокининов в корнях инокулированных бактериями двухсуточных проростков было примерно в 3 раза ниже, чем у контрольных. К четвертым суткам содержание цитокининов в
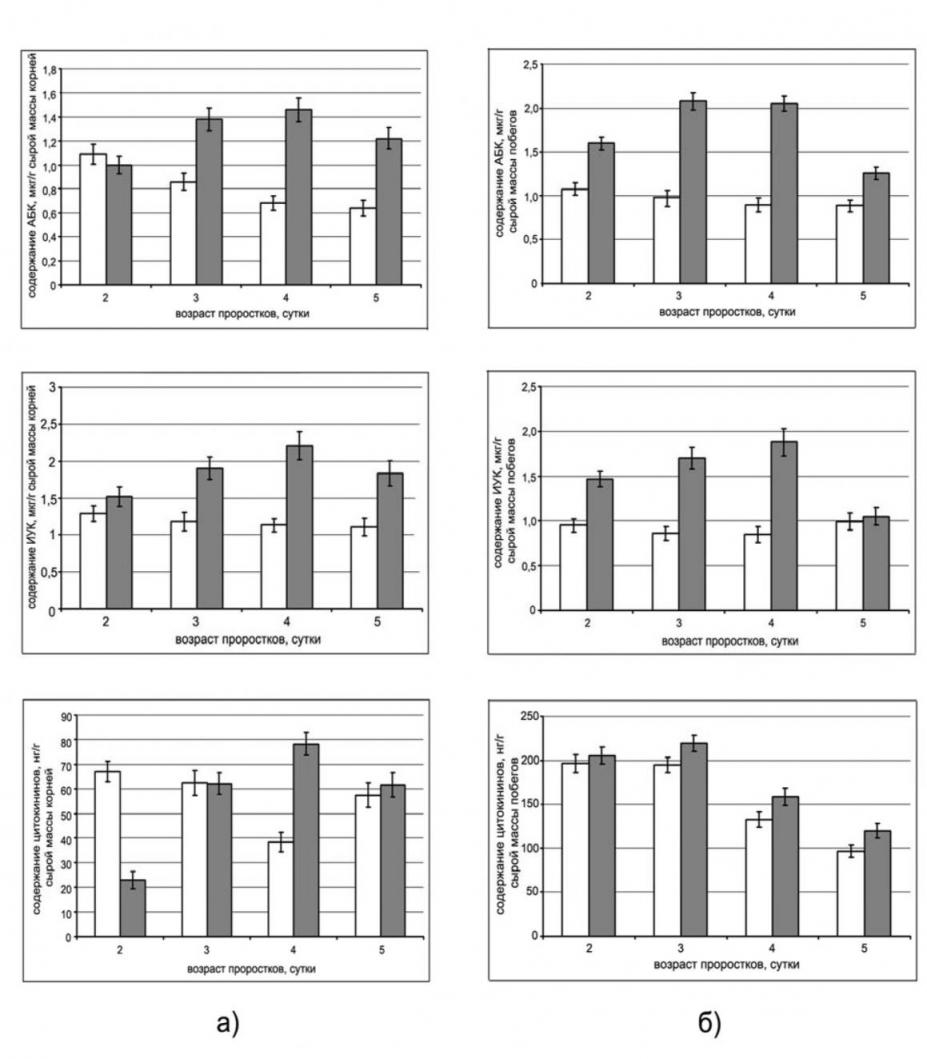
Рисунок 2. Динамика содержания АБК, ИУК и цитокининов в корнях (а) и побегах (б) проростков пшеницы: ![]() – контроль,
– контроль, ![]() – семена обработаны спорами B. subtilis 11ВМ
– семена обработаны спорами B. subtilis 11ВМ
корнях обработанных проростков повышалось, в то время как в побегах их содержание незначительно превышало контроль. Если накопление АБК в инокулированных растениях можно объяснить защитным ответом растительных клеток на внедрение бактерий в ткани, то повышенное содержание ИУК, наряду с другими причинами, могло быть вызвано снижением активности ферментов, участвующих в деградации ауксинов, например, оксидазы ИУК. Это предположение подтвердилось тем, что первоначальная активация фермента в корнях трехсуточных проростков, инокулированных бактериями, в сравнении с контрольными, сменялась более чем двукратным падением активности ИУК-оксидазы к четвертым суткам. Таким образом, максимальное накопление ауксинов в корнях под действием эндофита совпало со снижением активности фермента в них в сравнении с контрольными.
Мобилизация эндофитами фосфатов кальция, железа и алюминия
Способность PGPB к повышению биодоступности элементов минерального питания растений, особенно фосфора, также может вносить существенный вклад в стимуляцию роста с.-х. культур. Оценка способности штаммов мобилизовать фосфаты на плотной среде Муромцева показала, что все они могут растворять фосфат железа, а большинство – и фосфат кальция (кроме штаммов М1, НТ, 118РН, 962РН).
Ни один штамм не растворял фосфат алюминия. Фосфат кальция активнее растворяли штаммы 89РН, 922РН, 49РН (по убыванию активности), фосфат железа – штаммы 121РН, 922РН, 171РН. При этом отсутствие способности некоторых штаммов к мобилизации фосфата кальция может являться артефактом, поскольку известно, что осаждение цитрата и оксалата кальция мешает обнаружению зоны гало, несмотря на растворение фосфата (Fankem et al, 2006). В связи с этим можно полагать, что применение описанного метода недостаточно для выявления фосфат-мобилизующих микроорганизмов, поскольку многие изоляты, не формирующие зоны гало на агаризованных средах, могут растворять фосфаты в жидкой среде. Действительно, при культивировании штаммов в жидкой среде NBRIP мобилизация фосфата кальция происходила активнее, чем фосфатов железа и алюминия, а растворение AlPO4 – в целом лучше, чем FePO4, что согласуется с данными литературы (Ahn, 1993). Данные по мобилизации Ca3(PO4)2 в жидкой среде позволили выделить группу «штаммов-лидеров» (11ВМ, 118РН, 162РН, 24Д, 11В), состав которой отличался от такового на плотной питательной среде. Фосфат алюминия активнее других растворяли штаммы 871РН, 962РН. В отличие от способности к растворению минеральных фосфатов, фосфатазная активность наблюдалась, примерно, у половины изученных штаммов.
Известно, что основным механизмом растворения фосфатов микроорганизмами является выделение ими органических кислот. Органические кислоты, продуцируемые некоторыми из штаммов, представлены в табл. 4. Набор кислот зависел от штамма и консистенции среды. Все бациллы
Таблица 4 – Органические кислоты в составе экзометаболитов некоторых штаммов при культивировании на среде NBRIP с добавлением Ca3PO4, нг/мл
| Кислоты | Штаммы | ||||||
| 26Д | 832РН | 11РН | 922РН | 11ВМ | |||
| Лимонная | Ж* | сл** | - | - | сл** | - | |
| П*** | -**** | - | - | - | |||
| Пропионовая | Ж | 161 | 100 | ||||
| П | - | - | - | - | - | ||
| Малоновая | Ж | - | - | 316 | 576 | 1106 | |
| П | 177 | 447 | 447 | 382 | 508 | ||
| Янтарная | Ж | 255 | 289 | - | - | 426 | |
| П | 114 | сл** | - | - | |||
| Щавелевая | Ж | сл** | - | сл** | сл** | сл** | |
| П | - | - | - | - | - | ||
| Винная | Ж | - | сл** | 483 | сл** | 158 | |
| П | - | - | - | - | - | ||
| Изолимонная | Ж | 326 | 1006 | 2190 | - | 719 | |
| П | - | 125 | 112 | 210 | 207 | ||
| Молочная | Ж | 737 | 794 | 1076 | 735 | 935 | |
| П | 269 | 276 | 236 | 305 | 636 | ||
* – жидкая среда; ** – следы, менее 100 нг/мл; *** – плотная среда;****– кислота не была обнаружена
продуцировали монокарбоновую молочную кислоту на обеих средах. Присутствие в составе метаболитов бацилл ди- и трикарбоновых кислот позволяет предполагать, что мобилизация фосфатов может происходить не только благодаря снижению рН среды, но и за счет хелатирования ионов металлов.
Интересным, на наш взгляд, является отсутствие идентичности в спектрах кислот, выделяемых штаммами на средах с разной консистенцией. У большинства штаммов среди метаболитов в плотной среде удалось идентифицировать меньше видов кислот, по сравнению с жидкой. Интересен также факт продукции малоновой кислоты всеми штаммами на плотной среде, тогда как в жидкой среде эту кислоту продуцировали только штаммы 11ВМ, 11РН и 922РН.
Антагонизм эндофитов по отношению к фитопатогенным грибам и защитное действие при воздействии фитотоксинов на проростки пшеницы
Одними из наиболее опасных фитопатогенов из-за их способности продуцировать микотоксины, в т.ч. и фитотоксины, являются грибы рода Fusarium. Ранее Кутлубердиной Д.Р. (2010) детально были охарактеризованы взаимоотношения штамма B. subtilis 11РН с грибами F. sporotrichioides, F. poae, F. graminearum, F. avenaceum, в результате чего он был охарактеризован как самый сильный антагонист к этим фитопатогенам, способный также повышать устойчивость растений к действию фузариотоксинов. Нами показано, что исследованные штаммы B. subtilis также способны ингибировать рост F. sporotrichioides и F. poae, выделенных из зерна озимой ржи. При этом самыми сильными антагонистами были штаммы 49РН, 89РН, 11ВМ и 832РН в случае F. sporotrichioides и 121РН, 11ВМ и 112РН в случае F. poae.
Оценка способности штамма B. subtilis 11ВМ проявлять защитный эффект, подобный ранее изученному штамму 11РН (Кутлубердина, 2010), показала, что он эффективнее снижает фитотоксичность метаболитов F. avenaceum в сравнении с «эталонным» штаммом 26Д (табл. 5). В случае действия фитоксинов F. poae протекторная активность этого штамма была слабее «эталонного». Факт наличия защитного эффекта у этих двух эндофитных штаммов, а также штамма B. subtilis 11РН (Кутлубердина, 2010), независящего от вида гриба, свидетельствует, что протекторный механизм этих эндофитных бактерий по отношению к растениям при действии на них фузариотоксинов может быть опосредован через хозяина.
Таблица 5 – Сравнительный защитный эффект штамма B. subtilis 11ВМ при действии фузариотоксинов на проростки пшеницы
| Вариант | Длина, мм | ||||||||
| F. sporotrichioides | F. poae | F. graminearum | F. avenaceum | ||||||
| корень | побег | корень | побег | корень | побег | корень | побег | ||
| контроль | |||||||||
| Вода | 65±4 | 35±2 | 62±4 | 31± | 67±5 | 39±4 | 63±4 | 34±2 | |
| B.s. 11ВМ | 76±4 | 51±4 | 71±6 | 40±3 | 75±5 | 53±4 | 72±3 | 43±2 | |
| B.s. 26Д | 70±4 | 37±2 | 65±5 | 33±2 | 73±3 | 42±2 | 68±3 | 35±2 | |
| + фитотоксины (культуральный фильтрат) | |||||||||
| Контроль | 16±1 | 18±1 | 17±1 | 22±1 | 15±1 | 25±2 | 20±1 | 12±1 | |
| B.s. 11ВМ | 18±1 | 25±1 | 22±1 | 23±2 | 20±2 | 29±2 | 30±2 | 26±2 | |
| B.s. 26Д | 19±2 | 25±2 | 23±1 | 23±1 | 31±2 | 27±2 | 24±2 | 14±1 | |
| + фитотоксины (культуральный фильтрат) в разбавлении с водой 1:1 | |||||||||
| Контроль | 25±2 | 29±1 | 44±2 | 25±2 | 38±3 | 30±1 | 31±2 | 27±2 | |
| B.s. 11ВМ | 31±2 | 33±2 | 49±3 | 29±1 | 49±4 | 36±3 | 45±4 | 31±2 | |
| B.s. 26Д | 27±1 | 25±1 | 62±3 | 31±1 | 47±4 | 36±3 | 41±3 | 29±1 | |
Взаимоотношения эндофитных бацилл с азотфиксирующими бактериями
В природе не исключен антагонизм PGPB не только к фитопатогенам, но и к хозяйственно полезным видам бактерий, например, ассоциативным и симбиотическим, обеспечивающим растения азотом. Выявлено, что из 23 штаммов 19 подавляли рост азотфиксаторов, указанных в табл. 6. (приведены свойства самых активных штаммов). Наиболее чувствительными к бациллам оказались бактерии A. brasilense, рост которых угнетали 17 штаммов, и A. lipoferum, размеры зон отсутствия роста которых были наибольшими. Рост A. irakense подавляли 3 штамма, Azotobacter vinelandii – не подавлял ни один из эндофитов. Интересно, что депонированные штаммы (26Д, 24D, 11В, М1) не ингибировали рост азотфиксаторов или проявляли антагонизм лишь к одному из видов азотфиксирующих бактерий.
Таблица 6 – Штаммы B. subtilis, наиболее антагонистичные по отношению к бактериям-азотфиксаторам
| Штамм | Площадь зоны подавления роста бактерий, мм2 | |||||
| Azospirillum brasilense | Azospirillum lipoferum | Azospirillum irakense | Azotobacter chroococcum | Azotobacter vinelandii | Rh. legumi-nosarum | |
| НТ2 | - | - | - | - | - | 31±2 |
| 26Д | - | - | - | - | - | 38±3 |
| М1 | 133±9 | - | - | - | - | - |
| 162РН | 143±7 | - | - | - | - | - |
| 871РН | 97±4 | - | 27±2 | - | - | - |
| 11ВМ | 82±4 | 123±9 | -* | - | - | 36±2 |
| 832РН | 106±7 | 168±12 | - | - | - | 36±2 |
| 11РН | 165±8 | 295±18 | - | 605±34 | - | - |
| 118РН | 133±5 | 244±14 | - | 207±9 | - | 27±1 |
| 171РН | 113±7 | 165±13 | 25±2 | - | - | 51±4 |
| 49РН | 87±4 | 165±7 | 25±1 | 371±29 | - | 56±2 |
* – отсутствие подавления роста
Безопасность и технологичность эндофитного штамма B. subtilis 11ВМ
Безопасность штамма B. subtilis 11ВМ оценивали на основе договора с Уфимским НИИ МТиЭЧ Роспотребнадзора РФ. При введении клеток штамма различными способами мышам и крысам смертности среди них не наблюдалось в течение 15 дней ни от одной из доз препарата, что позволяет отнести изучаемый штамм к группе невирулентных. Штамм не проявлял токсичных свойств. Клеточный состав крови подопытных животных не имел достоверных различий от контрольных. Диссеминации во внутренние органы животных не обнаружено. Скармливание клеток штамма достоверно не изменяло частоту встречаемости и количество полезной микрофлоры кишечника в сравнении с контрольными. Таким образом, штамм B. subtilis 11ВМ безвреден для животных.
Перспектива производства препарата на основе микроорганизмов определяется не только их полезной биологической активностью и безопасностью, но и возможностью промышленного культивирования. В связи с этим испытывали возможность и эффективность культивирования штамма 11ВМ, используя газо-вихревой биореактор «БИОК». Изменения определяемых параметров биомассы штамма при культивировании, в целом, были сходными с таковыми у ранее культивировавшихся штаммов B. subtilis 26Д и 11В (Лукьянцев, 2010). Однако в отличие от них переход культуры в стационарную фазу развития у штамма 11ВМ наблюдался уже через 6 ч роста, в то время как штаммы 26Д и 11В достигали этой стадии через 8-10 ч. Спорообразование у штамма 11ВМ также начиналось раньше в сравнении с указанными. Концентрация жизнеспособных спор в препарате составляла 3109 КОЕ/мл. Таким образом, показана возможность культивирования штамма B. subtilis 11ВМ в биореакторе в одинаковых для коммерческого штамма B. subtilis 26Д условиях.
ВЫВОДЫ
1. 17 исследованных новых штаммов бактерий относятся к виду B. subtilis.
2. Один из механизмов стимуляции роста пшеницы эндофитными штаммами B. subtilis, обитающими в ее внутренних тканях, опосредован через фитогормон-подобное действие клеток бактерий на растения, а также изменение уровня фитогормонов АБК, ИУК и цитокининов в растительных тканях, характеризующееся на первоначальном этапе взаимоотношений растение-эндофит одновременным повышением в корнях и побегах содержания как ИУК, так и АБК.
3. Повышение содержания ауксинов в проростках пшеницы, инокулированных эндофитными штаммами бактерий B. subtilis, может происходить благодаря снижению активности оксидазы ИУК под влиянием клеток этих бактерий.
4. Все исследованные штаммы B. subtilis способны растворять in vitro фосфаты кальция и железа, и большинство из них – фосфат алюминия. Активность растворения фосфатов одним и тем же штаммом зависит от консистенции среды и может быть обусловлена выделением органических кислот в среду культивирования.
5. Исследованные виды азотфиксирующих бактерий Azospirillum brasilense, A. lipoferum, A. irakense, Azotobacter chroococcum, A. vinelandii, Rh. leguminosarum bv. viceae отличаются между собой по чувствительности in vitro к ростингибирующему действию метаболитов B. subtilis. Большинство изученных штаммов бацилл подавляют рост исследованных представителей A. brasilense и A. lipoferum; ни один из штаммов не ингибирует рост представителя A. vinelandii.
6. Повышение урожайности с.-х. культур и их устойчивости к болезням при инокуляции растений эндофитными штаммами B. subtilis происходит благодаря комплексу их полезной биологической активности, в том числе: способности стимулировать рост растений, растворять неорганические фосфаты, повышая их подвижность в почве, продуцировать органические кислоты, хелатирующие ионы микроэлементов, проявлять антагонизм к фитопатогенным грибам и повышать устойчивость растений к действию их фитотоксинов, избирательно влиять на рост различных видов ассоциативных и симбиотических азотфиксирующих бактерий.
ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ
1. Эндофитный штамм B. subtilis 11ВМ, депонированный в коллекции ВНИИСХМ под номером 519, безопасен для животных, технологичен и может быть использован для разработки препаратов с комплексной биологической активностью, повышающих урожайность с.-х. культур и их устойчивость к болезням.
2. Для выявления ауксин- или цитокинин-подобной активностей эндофитов рекомендуется использовать методику, в которой, соответственно, отрезки колеоптилей или листьев получают из растений, предварительно инокулированных этими микроорганизмами, используя в качестве контроля неинокулированные растения.
СПИСОК ОПУБЛИКОВАННЫХ РАБОТ
Статьи в журналах, рекомендованных ВАК РФ
1. Хайруллин Р.М. Биологические особенности эндофитных штаммов Bacillus subtilis как перспективной основы новых биопрепаратов / Р.М. Хайруллин, А.А. Егоршина, М.А. Лукьянцев, Н.А. Уразбахтина, Р.Ш. Иргалина, А.Р. Сахабутдинова // Аграрная Россия. – 2011. – №1. – С. 49-53.
2. Егоршина А.А. Фосфат-мобилизующая активность эндофитных штаммов Bacillus subtilis и их влияние на степень микоризации корней пшеницы / А.А. Егоршина, Р.М. Хайруллин, М.А. Лукьянцев, З.М. Курамшина, Ю.В. Смирнова // Журнал Сибирского Федерального Университета. Серия Биология. – 2011. – №2. – С. 172-182.
3. Егоршина А.А. Участие фитогормонов в формировании взаимоотношений проростков пшеницы с эндофитным штаммом Bacillus subtilis 11ВМ / А.А. Егоршина, Р.М. Хайруллин, А.Р. Сахабутдинова, М.А. Лукьянцев // Физиология растений. – 2012. – Т.59. – № 1. – С. 148-154.
Статьи в других изданиях
1. Егоршина А.А. Стимуляция роста и продуктивности растений эндофитными бактериями: разработка алгоритмов поиска эффективных штаммов и создания препаратов / А.А. Егоршина // Биомика – наука XXI века: материалы школы-семинара молодых ученых Уфимского научного центра РАН и Волго-Уральского региона по физико-химической биологии и биотехнологии, 9-15 сентября, 2007. – Уфа, 2007. – С. 44-46.
2. Хайруллин Р.М. Механизмы повышения адаптационного потенциала и продуктивности растений в сообществе с микроорганизмами / Р.М. Хайруллин, А.А. Егоршина // Фундаментальные и прикладные аспекты исследования симбиотических систем: материалы всероссийской конференции с международным участием, 25-27 сентября, 2007. – Саратов, 2007. – С. 41.
3. Кутлубердина Д.Р. Антагонистические взаимоотношения эндофитных бактерий Bacillus subtilis с грибами рода Fusarium / Д.Р. Кутлубердина, Р.М. Хайруллин, А.А. Егоршина // Стратегия взаимодействия микроорганизмов и растений с окружающей средой: материалы IV межрегиональной конференции молодых ученых, 14-16 октября, 2008. – Саратов, 2008. – С.16.
4. Хайруллин Р.М. Разработка полифункционального биофунгицида «СТАРТ» на основе эндофитного штамма Bacillus subtilis 11ВМ / Р.М. Хайруллин, Н.А. Уразбахтина, А.А. Егоршина, М.А. Лукьянцев // Биотехнология: состояние и перспективы развития, часть 1: материалы пятого московского международного конгресса, 16-20 марта, 2009. – Москва, 2009. – С. 320-321.
5. Егоршина А.А. Влияние эндофитного штамма Bacillus subtilis 11ВМ на фитогормональный статус проростков пшеницы / А.А. Егоршина, А.Р. Сахабутдинова, М.А. Лукьянцев, Р.М. Хайруллин // Симбиоз Россия 2009: материалы II Всероссийского с международным участием конгресса студентов и аспирантов-биологов, 25-29 мая 2009. – Пермь, 2009. – С. 25-26.
6. Егоршина А.А. Мобилизация нерастворимых неорганических фосфатов эндофитными штаммами Bacillus subtilis / Егоршина А.А., Хайруллин Р.М., Лукьянцев М.А. // XII з’зд Товариства мiкробiологiв Украни iм. С.М. Виноградського: тези доповiдей, 25-30 травня 2009. – Ужгород, 2009. – С. 302.
7. Лукьянцев М.А. Эндофитный штамм Bacillus subtilis 49РН как перспективная основа нового биофунгицида / М.А. Лукьянцев, А.А. Егоршина, Р.М. Хайруллин // Проблемы региональной экологии в условиях устойчивого развития, часть 2: материалы Всероссийской научно-практической конференции, 1-2 декабря, 2009. – Киров, 2009. – С. 42-43.
8. Егоршина А.А. Защитный эффект эндофитных штаммов Bacillus subtilis при воздействии фузариотоксинов на проростки злаков / А.А. Егоршина, Д.Р. Кутлубердина, Р.М. Хайруллин // Биомика – наука XXI века: материалы II Всероссийской школы-конференции молодых ученых Уфимского научного центра РАН и Волго-Уральского региона по физико-химической биологии и биотехнологии, 27-29 сентября, 2011. – Уфа, 2011. – С. 35-36.