

Японский (grus japonensis p.l.s. m ller, 1776 ) и даурский (grus vipio pallas, 1811 ) журавли архаринской низменности (среднее приамурье)
На правах рукописи
АНДРОНОВ Владимир Андреевич
Японский (Grus japonensis P.L.S. Mller, 1776) и даурский
(Grus vipio Pallas, 1811) журавли Архаринской низменности
(Среднее Приамурье)
03.00.08 - зоология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Владивосток
2008
Работа выполнена в лаборатории орнитологии Биолого-почвенного института ДВО РАН и в Государственном природном заповеднике «Хинганский» Росприроднадзора
Научный руководитель: доктор биологических наук,
старший научный сотрудник
Нечаев Виталий Андреевич
Официальные оппоненты: доктор биологических наук,
профессор
Бочарников Владимир Николаевич
кандидат биологических наук,
доцент
Глущенко Юрий Николаевич
Ведущее учреждение: Институт водных и экологических проблем
ДВО РАН, г. Хабаровск
Защита состоится «4» декабря 2008 года в «14.30» часов на заседании диссертационного совета Д 005.003.03 при Биолого-почвенном институте ДВО РАН по адресу: 690022, Владивосток, проспект 100-лет Владивостоку, 159.
Факс: (4232) 310-193
С диссертацией можно ознакомиться в Центральной научной библиотеке ДВО РАН
Автореферат разослан « » ноября 2008 г.
Ученый секретарь
диссертационного совета,
кандидат биологических наук Е.М.Саенко
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Редкие и находящиеся под угрозой исчезновения виды животных – самая хрупкая, но очень важная часть биоразнообразия. Видовое разнообразие, обусловленное длительным процессом эволюции, составляет основу целостности экосистем и биосферы в целом (Стратегия..., 2004). Без комплексного изучения биологии редких видов, как основы для разработки мер по защите и восстановлению численности, невозможно представить стратегическую и тактическую линию на спасение любого редкого или исчезающего вида животных (Флинт, 2000). Журавли – одна из наиболее уязвимых групп птиц. Актуальность проблем охраны редких видов журавлей давно стала предметом самого пристального внимания исследователей. Будущее журавлей зависит от правильности выработанных мер по сохранению популяций и среды их обитания. Многолетнее и разноплановое биологическое исследование наиболее полно и достоверно помогает выяснить причины неблагополучного состояния популяций в природе.
Японский (Grus japonensis P.L.S. Mller, 1776) и даурский (Grus vipio Pallas, 1811) журавли – исчезающие виды птиц (Красная книга РФ, 2001). Оба вида обитают на Дальнем Востоке. Распространение японских и даурских журавлей на территории России приурочено к бассейну Амура и Архаринская низменность, расположенная в Амурской области, входит в гнездовую часть ареала этих видов.
С одной стороны, наличие на Архаринской низменности различных категорий особо охраняемых природных территорий и включение Архаринской низменности в 1994 году в Список территорий, имеющих международное значение, главным образом в качестве местообитаний водоплавающих и околоводных видов птиц (Рамсарская конвенция), подчеркивает ее высокий природоохранный статус, который способствует сохранению журавлей. С другой, высокая плотность населения и сельскохозяйственное освоение территории создают как благоприятные, главным образом, кормовые, условия для жизни журавлей, так и угрозы – это пожары, браконьерство, фактор беспокойства и др.
Все вместе предопределило важность и своевременность наших исследований по изучению биологии исчезающих видов журавлей на данной территории.
Целью исследований являлось выяснение современного состояния популяций японского и даурского журавлей на Архаринской низменности и определение негативных факторов, влияющих на журавлей и среду их обитания.
Для достижения цели определены следующие задачи:
- Изучить территориальное распределение японских и даурских журавлей на модельных участках Архаринской низменности.
- Определить экологические параметры обитания этих видов.
- Определить демографические показатели двух видов журавлей и факторы, их лимитирующие.
- Провести сравнительное изучение биологии двух видов журавлей.
- Выявить пути миграций и места зимовок японских и даурских журавлей гнездящихся на Архаринской низменности.
- Выработать рекомендации по сохранению японских и даурских журавлей на Архаринской низменности.
Научная новизна. Впервые на территории России проведен многолетний мониторинг численности, распределения, успеха гнездования и других особенностей биологии. двух исчезающих видов журавлей – японского и даурского. Впервые проведен анализ территориального распределения и динамики численности. Определены показатели размножения и факторы, влияющие на состояние популяций двух видов журавлей в современных условиях обитания. Получены новые сведения о биологии и распространении японских и даурских журавлей на Архаринской низменности, о местах их зимовок и миграционных остановках. Предложены научно обоснованные меры для сохранения и восстановления популяций японского и даурского журавлей на Архаринской низменности.
Теоретическая и практическая ценность. Теоретическая ценность работы заключается в возможности использования результатов многолетнего мониторинга в анализе влияния глобальных природных факторов и антропогенных воздействий на изучаемые виды в пределах всего ареала видов. Сведения о распределении гнездовых участков журавлей помогут в организации мониторинговых работ на Архаринской низменности. Данные о биотопическом распределении журавлей будут способствовать резервированию местообитаний в других частях ареала. Информация по миграциям важна для организации охраны мест остановок и зимовок журавлей. Полученные сведения помогают в разработке природоохранных мероприятий по сохранению журавлей и среды их обитания в Хинганском заповеднике и других заповедниках юга Дальнего Востока. Материалы исследований использованы при подготовке обоснований к созданию Станции реинтродукции редких видов птиц Хинганского заповедника, по присоединению государственного природного заказника «Ганукан» к Хинганскому заповеднику, при написании видовых очерков о японском и даурском журавлях для последнего издания Красной книги Российской Федерации.
К предложенным мерам по сохранению и восстановлению популяций японского и даурского журавлей на Архаринской низменности относятся: реинтродукция журавлей в природу; методика проведения контролируемых отжигов по защите гнездовий журавлей; предложение экологического страхования среды обитания и популяций редких видов птиц в зоне возможного ущерба; разработки программ экопросвещения населения, посвященным редким видам («День журавля», «Озеро белых птиц» и др.). Одним из реализованных автором природоохранных проектов по сохранению японских и даурских журавлей стало создание особого структурного подразделения в Хинганском заповеднике – Станции реинтродукции редких видов птиц.
Апробация работы. Материалы исследований доложены на советско-японском симпозиуме (Хабаровск 1984), совещаниях Рабочей группы по журавлям СССР (Матсалуский заповедник – 1984, Хинганский заповедник 1986, Караганда 1988), заседаниях Амуро-Уссурийского отделения Орнитологического общества СССР (Владивосток – 1986, 1987, 1988, 1989), Всесоюзной орнитологической конференции (Ленинград – 1986), Всесоюзном совещании по проблемам зоокультуры (Москва – 1986), Международном симпозиуме по журавлям Палеарктики (Таллинн – 1989), региональной конференции «Состояние и охрана Зейско-Буреинской равнины» (Благовещенск – 1991), Международном симпозиуме «Журавли и аисты бассейна р. Амур» (Хабаровск- Благовещенск – 1992), рабочих заседаниях Международного фонда охраны журавлей (США – 1990, 1998), международных конференциях по водно-болотным угодьям (КНР – 1992, 1997, 1998), заседаний Сети важнейших журавлиных территорий Юго-Восточной Азии (Пекин 1997, Благовещенск – 1998, Сеул 2001, Саппоро 2002), на международных встречах экспертов по охране журавлей и аистов (Япония – 1993, 1994, 1996, 1998), а также на отдельных заседаниях российско-японской Конвенции по охране перелетных птиц и среды их обитания (2003, 2006, 2007), на заседаниях и конференциях Рабочей группы по журавлям Евразии (2000, 2001, 2003) и других.
Публикации. По теме диссертации опубликовано 26 работ.
Структура и объем работы. Диссертация состоит из введения, обзора литературы, 6 глав, выводов, списка литературы и приложения. Работа изложена на 211 страницах и содержит 27 таблиц, 42 рисунка. Список использованной литературы включает 194 наименования, из которых 92 на иностранных языках.
Благодарности. Благодарю всех, кто за время моего длительного научного пути, оказывал помощь и ценные советы – И.А. Нейфельдт, [В.Е.Флинта], С.М.Смиренского, В.Ю.Ильяшенко, [Ю.Б.Пукинского], Ю.В.Шибаева, С.В. Винтера, а в период работы в Хинганском заповеднике – всех сотрудников заповедника.
Благодарю научного руководителя В.А.Нечаева и всех сотрудников лаборатории орнитологии БПИ ДВО РАН (А.А. Назаренко, Ю.В. Шибаева, [Н.М.Литвиненко], С.Г. Сурмача, О. П. Вальчук, В.Н. Чернобаеву) всегда оказывавших мне моральную поддержку в моих исследованиях.
Большую помощь в сборе полевого материала мне оказали студенты- дипломники. Постоянную и разнообразную поддержку на протяжении всего периода исследований мне оказывал Ю.А. Дарман, которому я искренне признателен.
Отдельно благодарю Р.С.Андронову за особую помощь при обсуждении, сборе, анализе и написании данной рукописи, а также за ее огромный вклад в многолетнюю деятельность Станции реинтродукции, которую мне довелось создать.
Глава 1. МАТЕРИАЛ И МЕТОДИКИ
Практическая работа выполнена в основном в период 1982-2001 гг., последний авиаучет – 2003 г. Исследования осуществлялись на двух модельных участках Хинганского государственного природного заповедника – Антоновское (филиал) и Лебединское лесничества (54969 га). Кроме того, учетные работы проводились в государственном природном заказнике «Ганукан» (64000 га). Заповедник и заказник представляют Архаринскую низменность.
Полевые исследования основывались на наземных учетах всей равнинной территории заповедника – фактически абсолютный учет территориальных пар журавлей. Постоянным мониторингом охвачены главные периоды гнездовой жизни журавлей. Авиаучеты на территории низменности проводились регулярно до 4 раз в год в 1983-1998 гг., а также в 2000 и 2003 гг. Общий период полевых исследований – 129 месяцев, протяженность авиаучетных маршрутов – свыше 42000 км (389 летных часов), пеших 9300 км, авто 17900 км. Суммарно учтено 139 гнездящихся пар даурских и 253 пары японских журавлей. Найдено 167 гнезд и гнездовых участков двух видов. Описано 27 гнезд и 37 кладок. Учтено 72 выводка даурских и 155 выводков японских журавлей. Объем суточных наблюдений составил: в гнездовой период 269 ч, в послегнездовой – 376 ч. Материал полевых работ дополнен сведениями из научного архива заповедника за 1977-2000 гг., главным образом, для уточнения сроков миграций журавлей. При расчете площадей водно-болотных угодий Архаринской низменности использованы материалы Проектов лесоустройства заповедника и Лесохозяйственного регламента лесничества «государственный природный заповедник «Хинганский».
Изучение территориального поведения основывалось на наблюдениях за особями, помеченными цветными ножными кольцами и спутниковыми передатчиками. Всего на территории Амурской области окольцовано 192 даурских и 173 японских журавля; спутниковыми передатчиками – 8 журавлей.
При анализе влияния климатических факторов на состояние популяций даурского и японского журавлей использованы многолетние данные метеостанций пос. Архара (Амурская область) и г. Облучье (Еврейская АО). Математический анализ проведен в программах “Statistica 5” и “Microsoft Exel”.
Глава. 2. АРХАРИНСКАЯ НИЗМЕННОСТЬ: ТЕРРИТОРИЯ
ИССЛЕДОВАНИЯ И МЕСТА ОБИТАНИЯ ЖУРАВЛЕЙ
В главе приведена физико-географическая характеристика Архаринской низменности, описаны природных условий, обеспечивающие обитание журавлей.
Архаринская низменность включена в Рамсарскую конвенцию и входит в число основных мест гнездования японских журавлей материковой популяции, является важным участком гнездования даурских журавлей. Площади болот на Архаринской низменности составляют 27,64% и площади прочих нелесных земель – 20,81%.
Глава 3. ДАУРСКИЙ ЖУРАВЛЬ НА АРХАРИНСКОЙ НИЗМЕННОСТИ
Даурский журавль – редкий гнездящийся вид Архаринской низменности. Места обитания журавлей находятся в равнинных лесничествах Хинганского заповедника и на территории заказника «Ганукан».
Сроки прилета и отлета. Появление первых журавлей в Лебединском лесничестве, которое соседствует с Еврейской автономной областью, регистрируют, по данным многолетних наблюдений, в среднем 12.04. (n = 23). Самая ранняя встреча в лесничестве – 27.03.1990 г. В Антоновском лесничестве среднемноголетняя дата прилета – 09.04. (n = 24); самая ранняя встреча – 20.03.1990 г.
Несмотря на удаленность двух мониторинговых участков друг от друга на расстояние более 70 км, прилет даурских журавлей отмечается практически одновременно в разных местах низменности Средняя дата прилета для всей низменности соответствует 09.04. (n = 26). Наиболее благоприятные погодные условия для прилета даурских журавлей в места гнездования определяются среднесуточными температурами воздуха первой и второй декад апреля в интервале +0,2…4,1С. Прилет журавлей в места гнездования имеет корреляцию (p = 0,035) с переходом среднесуточных температур воздуха через +5С, среднемноголетняя дата температурного перехода на низменности – 14.04.
Отлет происходит с августа по октябрь. Средняя дата отлета на низменности соответствует 28.09. (n = 16). Отлет журавлей совпадает с переходом положительных среднесуточных температур приземного воздуха через 10°С к более низким значениям; среднемноголетняя дата температурного перехода на низменности соответствует 20.09. Завершающая стадия отлета (последняя декада сентября и первая декада октября) приходится на период установления среднесуточных наземных температур воздуха в пределах +12,4…7,9С.
Сезонная ритмика миграционной активности. Большинство птиц – 59,0%, возвращается в места гнездования в апреле. Осенняя миграция активно начинается уже во второй половине августа – 39,4%, в сентябре отмечается максимальное количество групп и стай – 50%. Массовый отлет происходит в сентябре, после чего темпы отлета постепенно снижаются ко второй половине октября. В октябре число стай сокращается почти в 5 раз по сравнению с сентябрем.
Число особей в стаях. В весенней миграции выделяются две волны движения журавлей. Первыми, в марте и апреле, регистрируют одиночных птиц (17,1%), и пары (60,8%). По видимому это местные птицы, которые остаются в заповеднике на гнездовой сезон. Прилет территориальных пар происходит в сжатые сроки. В следующие 5-7 дней со дня первой встречи журавлей на низменности, территориальные участки уже заселенны. Во второй половине апреля и в мае встречаются небольшие группы в среднем по 6 ± 2 птиц (n = 43), что отражает вторую волну миграции. На группы в 3-8 птиц в апреле приходится 17,7%, в мае – 33,0%. С конца мая стаи в 4 и более особей перестают отмечаться в заповеднике и вблизи его границ. Стаи свыше 20 птиц весной не встречаются.
Осенью даурских журавлей чаще регистрируют в парах (30,3%), и маленьких, обычно семейных, группах (33,3%). В сентябре встречи парных птиц становятся реже (25,8%), а групп в 3-4 птицы увеличивается до 34,8%. Стаи в среднем по 7 ± 2 птиц (n = 21), появляются, как правило, в сентябре. Крупные, свыше 20 птиц, пролетные стаи отмечаются редко и обычно в пиковой (15.09. 01.10.) фазе отлета на зимовку.
Биотопы и кормовые стации. На Архаринской низменности даурский журавль придерживается открытых, слабо увлаженных пространств в окружении лесокустарниковой растительности. В составе индивидуальной территории отмечены естественные биотопы, нарушенные в разной степени человеком, и сельхозугодия. В разные месяцы гнездового сезона пары используют тот или иной тип биотопа, либо их совокупность (табл. 1). Предпочтительными биотопами в гнездовой период являются лугово-болотные, редко с элементами степной растительности, растительные сообщества с доминированием вейника и осок, с примесью разнотравья и вкраплениями незрелых лесных массивов. В послегнездовой период биотопы с нарушенной средой обитания для журавлей предпочтительнее, чем другие. Типичный гнездовой участок даурских журавлей – это небольшого размера переувлажненная мочажина, нередко среди поля или нарушенной среды обитания. Уровень воды около гнезда – 20-40 см, в среднем 29 см (n = 11). Присутствие воды на гнездовом участке обязательно. Гнездовой участок располагается, как правило, в понижениях (западинах) микрорельефа местности. Нередки случаи совместного гнездования японских и даурских журавлей на одном участке.
Таблица 1.
Биотопы, используемые даурскими журавлями в гнездовой сезон
на Архаринской низменности
| Биотоп | Использование биотопов журавлями (число регистраций в биотопе): | |||||||
| Март | Апрель | Май | Июнь | Июль | Август | Сентябрь | Октябрь | |
| Поля (пшеничные, соевые, гречишные, ячменные) | 60% (6) | 33,3% (68) | 27,6% (45) | 29,7% (30) | 36,2% (17) | 19,6% (10) | 79,7% (63) | 93,3% (14) |
| Естественная среда обитания на ООПТ | 20% (2) | 30,9% (63) | 32,5% (53) | 30,7% (31) | 38,3% (18) | 47,0% (24) | 6,3% (5) | 0 |
| Нарушенная среда обитания | 20% (2) | 31,4% (64) | 35,6% (58) | 35,6% (36) | 23,4% (11) | 31,4% (16) | 8,9% (7) | 6,7% (1) |
| Деградирующая среда обитания | 0 | 4,4% (9) | 4,3% (7) | 4,0% (4) | 2,1% (1) | 2,0% (1) | 5,1% (4) | 0 |
| Итого | 100% (10) | 100% (204) | 100% (163) | 100% (101) | 100% (47) | 100% (51) | 100% (79) | 100% (15) |
Журавли используют влажные биотопы только в гнездовой период. В биотопах с высоким уровнем воды птицы не встречены. Не все кормовые стации входят в состав индивидуальной территории. Предпочтительными кормовыми стациями в гнездовой период являются осоковые заболоченные луга и зерновые поля. В послегнездовой период семья покидает гнездовой участок и перебирается ближе к суходольным лугам и сельхозугодиям. Биотопы естественной среды обитания в июне-июле используются только на 30,7-38,3%. Осенью, начиная с августа, птицы кормятся на полях (табл. 1). Места отдыха и ночевки входят в состав индивидуальной территории пары; кормовые стации обычно находятся в стороне от индивидуальной территории. У птиц нет привязанности к крупным водоемам и влажным лугам в летний период.
Территориальное распределение пар на низменности. Территориальные участки даурских журавлей располагаются по периферии охраняемой территории (заповедника, заказника) в зоне повышений микрорельефа. Средняя плотность гнездования на низменности (водно-болотные угодья (ВБУ) составляют 583,1 км2) - 0,024 пары/км2. Территориальные участки в плотных местах гнездования находились на расстоянии 1-8 км друг от друга. В годы благоприятные для размножения индивидуальные участки соседних пар примыкали и даже накладывались друг на друга.
В Антоновском филиале (ВБУ составляют 101 км2) выделены 14 территориальных участков. Плотность заселения филиала журавлями составила 0,01 пары/км2 при минимальной численности, при максимальной – 0,07 пары/км2. Территориальные участки, расположенные на возвышениях микрорельефа, использовались журавлями для гнездования чаще других. Доминирующими растительными сообществами на гнездовых территориях являются осоково-вейниковые заболоченные луга (42,8%) и вейниково-осоковые пушицевые болота (50%). Средний размер выводков на территориальных участках в возвышенной местности выше и составил 1,46 ± 0,14 птенца/пару (n = 25). Заселение территориальных участков журавлями происходило в среднем с частотой 3-4 сезона подряд и перерывами в 1-4, реже 5-7 лет. Самое продолжительное использование участка – 18 лет, однако, нам известен один участок, который журавли, по сообщению Н.С. Панькина, занимали с 1971 г., с некоторыми перерывами. Часто используемые и высокопродуктивные участки в филиале составили 42,8%.
В Лебединском лесничестве (ВБУ составляют 169 км2) выделены 10 территориальных участков. Практически все они располагались по границе лесничества и на сопредельной территории. Плотность гнездования в годы минимальной численности – 0,006 пары/км2, максимальной – 0,035 пары/км2. Территориальные участки относительно постоянные и использовались птицами от 2-х до 17 лет. Предпочтительный гнездовой биотоп (n = 26) – вейниково-осоковое пушицевое болото, в 84,6% случаев гнездования. Средний размер выводков на многолетних участках – 1,77 ± 0,23 птенца/пару (n = 16). Средняя продолжительность заселения территориального участка составила 7 сезонов, максимальная – 17 лет.
В заказнике (ВБУ составляют 314 км2) отмечено 11 территориальных участков. Плотность гнездования – 0,035 пары/км2. Средний размер выводков – 1,5 ± 0,5 птенца/пару (n = 10).
Анализ показывает, что не все потенциальные для размножения территориальные участки заселяются в гнездовом сезоне ежегодно, также неравномерно распределена нагрузка в многолетнем цикле. Имеются территориальные участки, которые пары заселяют в течение многих лет, как правило, они соседствуют с сельхозугодиями. На них зарегистрированы лучшие показатели естественного воспроизводства даурских журавлей. Однако, имеются участки, которые журавли используют с многолетними перерывами. К ним, в первую очередь, относятся территориальные участки, расположенные во влажных биотопах охраняемой территории, в отдалении от нарушенной среды обитания и сельхозугодий. Мы относим их ко второй и третьей категории, в зависимости от частоты заселения в многолетнем цикле наблюдений. Сравнение заселения территориальных участков в годы с обильными осадками и в годы засухи показывает, что 57% занятых гнездовых территорий в засушливые годы – это территории, расположенные в хорошо увлажненных биотопах.
Гнездовой консерватизм у журавлей имеется, однако это не означает, что на территориальном участке не может произойти смена журавлиных пар.
Динамика численности. В исследуемый период численность территориальных пар журавлей в заповеднике не оставалась постоянной. Всего учтено 158 территориальных пар, включая 37 пар без размножения или с отрицательным результатом. Средний показатель заселения модельных участков территориальными парами составил 6,5 ± 4,5 пар в год (n = 21). Число пар, не принимавших участие в размножении, в Антоновском лесничестве составило 28,6%, в Лебединском – 16,4%.
Динамика численности территориальных пар в заповеднике имела положительную тенденцию в период с 1980 по 1983 гг. – численность увеличилась с двух до семи территориальных пар. В следующем периоде, 1984-1991, наступила относительная стабилизация в 6-8 пар. Последний период, 1992-2000, также характеризовался положительной динамикой и число территориальных пар увеличилось до 10-11 в 1992 и 1997 гг. (рис. 1). За период исследований численность возросла почти в 5 раз.
Положительная динамика прослеживается и в изменении численности размножающихся пар. Число размножающихся пар составило 2-6 в 1981-1983 гг. и 5-7 пар в следующем, 1983-1991, периоде. Максимальное число гнездившихся пар зарегистрировали в 1997 г. – 10. В 2000 г. в заповеднике размножались только 4 пары (падение в 2,5 раза), но достоверной информации по размножению в 2000 г. нет (не прослежена судьба гнездившихся пар).
Сравнение динамики численности территориальных пар даурских журавлей в заповеднике со среднегодовым количеством осадков не обнаруживает видимой зависимости между этими показателями, что подтверждает и статистическая обработка данных (рис. 1). В отличие от погодных условий, пожары негативно сказывались на гнездовании журавлей в заповеднике. В годы с пожарами 25-60% пар не имело птенцов.
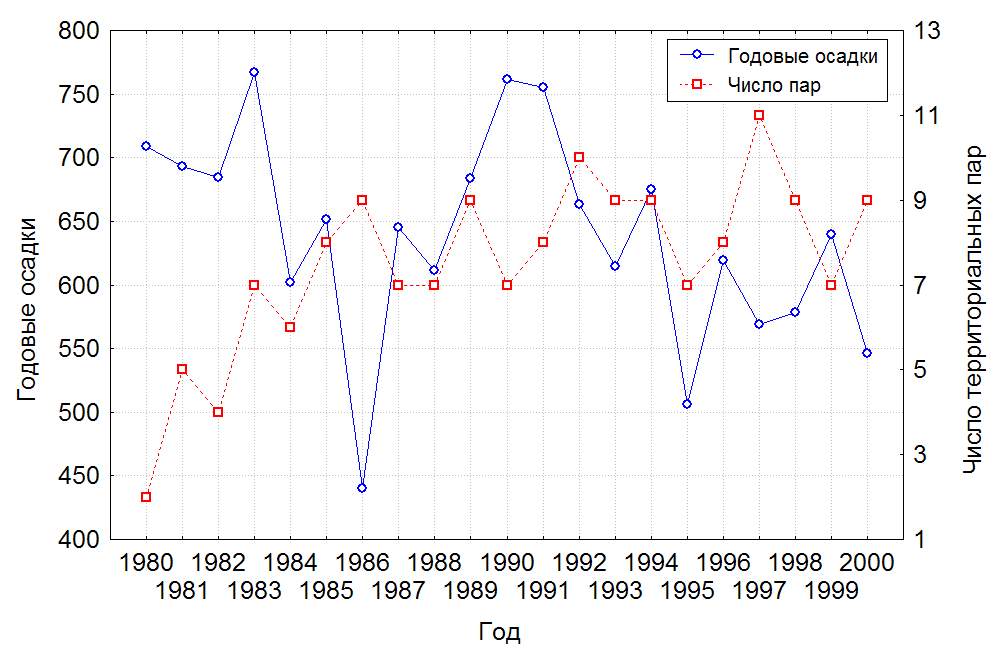
Рис. 1. Динамика численности территориальных пар даурских журавлей в Хинганском заповеднике и ее зависимость от среднегодового количества осадков.
Расчеты показали, что ёмкость биотопов низменности позволяет обитать здесь 33 парам даурских журавлей, именно столько выделено территориальных участков. В годы с минимальной численностью на низменности обитало 6-8 территориальных пар (24% от возможных 33), в обычные годы 11-13 пар (39,4%), в годы максимальной численности – 22 пары (66,7%). По данным авиаучета на низменности в 2003 г. отмечено 33 территориальные пары, т.е.ёмкость низменности была использована на 100%.
Продуктивность низменности, как территории естественного воспроизводства популяции даурского журавля, зависит от состояния среды обитания в сезон гнездования (ее целостность от пожаров, достаточное количество влаги) и состояния кормовой базы, а также от репродуктивного потенциала самой популяции (численности размножающихся пар). Климатический фактор (изменение суммы годовых осадков) на гнездование даурских журавлей оказывает незначительное влияние.
Демографические показатели популяции. В заповеднике ежегодно гнездилось в среднем 5,8 пары в год (n = 21). Средний размер кладок для низменности (учтены все кладки) – 1,98 яйца/пару (n = 49). Доля оплодотворенных яиц (учет выводков по 2 птенца и результаты инкубации природных кладок) – 95,45%. Данных по эмбриональной смертности в природных кладках нет. Средний размер выводков в заповеднике – 1,35 птенца/пару (n = 67).
Смертность птенцов до подъема на крыло в популяции, при известном среднем показателе размера выводка в 1,35 птенца/пару, могла составлять 29,3%. В зависимости от среднегодовой суммы осадков размер выводка изменялся (рис. 2), однако математического подтверждения зависимости нет. Тем не менее, данные таблицы 2 показывают, что если погодные условия мало влияли на гнездование журавлей, то в послегнездовой период выживаемость птенцов в природе зависела от данного природного фактора. Средний размер выводков в засушливые годы был меньше в 1,3 раза, чем в годы с достаточным увлажнением. Соответственно показатель птенцовой смертности до подъема на крыло в годы с недостаточным количеством осадков возрастал более чем в 2 раза.
Основная причина гибели кладок – пожары. Причины птенцовой смертности не установлены. Гибель птенцов отмечается в раннем возрасте, т.к. после 15 июля (возраст птенцов около двух месяцев) и до отлета на зимовку (данные авиаучетов) количество птенцов в семьях журавлей остается неизмененным.
Таблица 2.
Изменение показателей размножения даурских журавлей в Хинганском заповеднике
в зависимости от суммы годовых осадков
| Показатели | Осадки больше 660 мм/год (n = 8) | Осадки 560-660 мм/год (n = 10) | Осадки меньше 560 мм/год (n = 3) |
| Средний размер выводка | 1,6 (n = 28) | 1,18 (n = 32) | 1,3 (n = 7) |
| Потенциальный размер выводка* | 1,9 | 1,91 | 1,91 |
| Смертность в выводках, % | 15,8 | 38,2 | 31,9 |
| Доля размножающихся пар, % | 84,3 | 74,4 | 68** |
Примечание: * - 100% выход птенцов из оплодотворенных яиц (95,45%); ** - в этом климатическом отрезке доля размножающихся пар до 2000 г. составляла 81,2%, в 2000 г. – 44,4%, но по этому году нет точных данных о гнездовании.
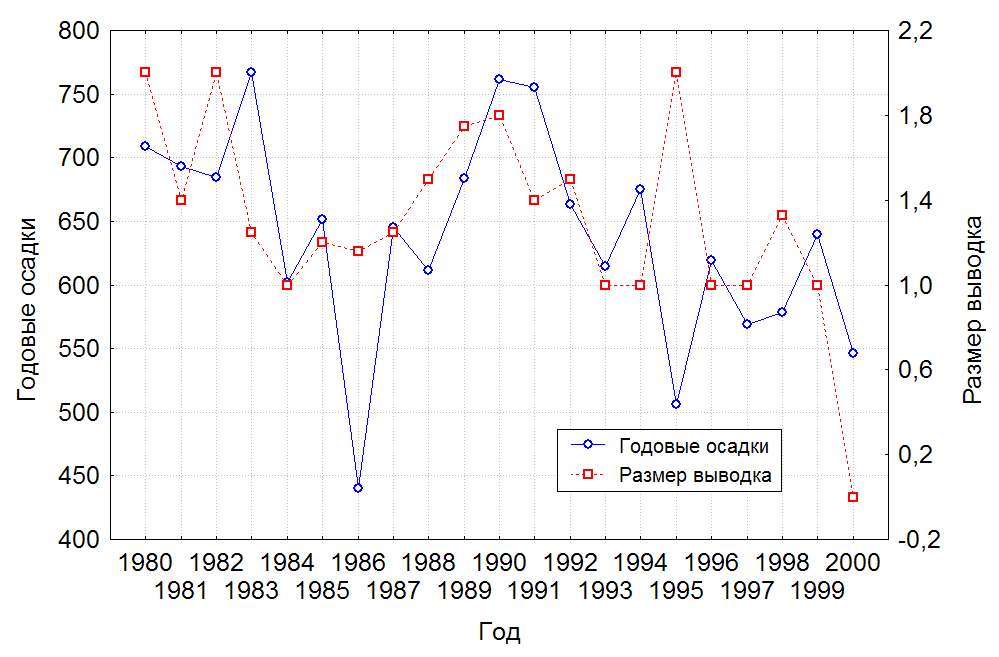
Рис. 2. Влияние осадков в сезон размножения на средний размер выводка у даурских журавлей.
Возрастная структура популяции даурского журавля на Архаринской низменности на конец лета выглядит следующим образом: 52,6% взрослых территориальных птиц, 31,6% неразмножавшихся и неполовозрелых особей и 15,8% летные птенцы.
Анализ демографических показателей свидетельствует о стабильном состоянии даурских журавлей на Архаринской низменности, что отмечено и в других участках ареала (Шибаев, Глущенко, 1988; Горошко, 2002; Chen Bin, Wang Zuoyi, 1991; Williams et al., 1991). Рост численности даурских журавлей на Архаринской низменности нами отмечен и в 2003 г. Средний размер выводков в 2003 г. составил 1,5 птенца/пару.
Гнездовая и послегнездовая жизнь. Приведены сведения о гнездовом и послегнездовом периодах, включая описание гнезд, кладок, суточную активность птиц.
Питание. Сезонные различия в питании связаны с доступностью и обилием корма в природе. Весной и осенью основу питания составляют зерновые культуры и семена диких злаков. Летом птицы больше потребляют почвенных беспозвоночных, насекомых и другой животный корм. Основные способы добывания корма: склевывание с поверхности субстрата и выкапывание из верхних слоев почвы.
Миграция и места зимовок. По информации о встречах окольцованных нами даурских журавлей на Архаринской низменности в ДМЗ (Корея) первых птиц отмечают в октябре: самая ранняя дата встречи – 18.10.2002, средняя дата прилета 25.10. (n = 5). На ДМЗ журавли проводят до недели, иногда больше. Часть птиц остается зимовать на Корейском полуострове, но большинство летит в Идзуми (Izumi)(Япония). В основном месте зимовки, в Идзуми, окольцованных журавлей встречали с 26.12. по 19.02. Подвижка птиц с места зимовки начинается в феврале. В марте журавли снова появляются в ДМЗ Кореи: ранняя встреча отмечена – 02.03.2004, средняя дата прилета – 12.03. (n = 11). Продолжительность остановки в ДМЗ весной составляет 2-9 дней.
Миграционный путь прослежен по даурскому журавлю с радиопередатчиком, которого пометили в Хинганском заповеднике в 1992 г. Период слежения – 08.07.1992-09.01.1993. Наблюдения со спутника вели в течение 186 дней и получили 181 точку с координатами перемещений. Длина перелета составила 2790 км, продолжительность миграции – 68 дней. Конечный пункт остановки – 9 января 1993 г., Идзуми В пути зафиксированы четыре остановки: на р. Бурея, на равнине Трехречье (Sanjianhg Plane) (Китай), оз Ханка, ДМЗ (Корея).
Глава 4. ЯПОНСКИЙ ЖУРАВЛЬ НА АРХАРИНСКОЙ НИЗМЕННОСТИ
Архаринская низменность является основным местом обитания японского журавля в Среднем Приамурье.
Сроки прилета и отлета. Отличительная особенность прилета японских журавлей – прилет парами и семейными группами в сжатые сроки. Средняя дата прилета на низменность – 02.04. (n = 23). Прилет журавлей совпадает с переходом среднесуточных температур приземного воздуха через 0°С к положительным значениям (p = 0,001), для низменности – это 4 апреля. Температурный интервал среднесуточной температуры воздуха для прилета журавлей составляет 8,2…+0,3°С. Снежный покров на марях и в лесу к прилету журавлей еще сохраняется, высота снега может составлять 7 см и более.
Отлет на зимовку также происходит в сжатые сроки. Первыми откочевывают неполовозрелые особи и неразмножавшиеся пары, их перестают регистрировать после 22.10. Отлет первых групп совпадает с переходом положительных среднесуточных температур воздуха через 5С к низким значениям. Обычно в эти же сроки выпадает первый снег. В среднем, после 30.10. (n = 24) японские журавли на Архаринской низменности практически не встречаются. Самые поздние встречи журавлей отмечены 24.11.1996 г. и 27.11.1989 г. Массовый отлет происходит в интервале среднесуточных температур воздуха +7,9…1,3С.
Сезонная ритмика миграционной активности. Массовый прилет в первую декаду апреля – 91,7% встреч. Отдельные особи, не более 8,3% регистраций, отмечены во второй половине марта. Осенью массовый отлет происходит в конце сентября – октябре. В октябре на 1,5% регистраций меньше по сравнению с сентябрем и в 11,5 раза больше, чем в ноябре. Число встреч птиц в ноябре в 8 раз меньше, по сравнению с октябрем.
Число птиц в стаях. Прилетающие группы журавлей насчитывают 1-2, реже 3-4 птицы. В марте прилет пар регистрируют в 53,3% случаев, в апреле – 61,0%. Одинаковый процент встреч групп в 3-4 птицы в марте (13,4%) и апреле (15,3%) объясняется присутствием в гнездовой части ареала неполовозрелых особей. Это могут быть прошлогодние птенцы, которые прилетают в составе своей семьи (n = 11) и молодые особи. Появление крупных стай весной – явление редкое.
Осенью первыми места размножения покидают одиночные птицы и пары, не имеющие потомства. Процент встреч одиночных журавлей в сентябре – ноябре сокращается с 7,2 до 0%. Последними места размножения покидают семьи. Встречи семейных групп (3-4 птицы) в ноябре составляют 60% всех встреч в этом месяце.
Биотоп и кормовые стации. Места обитания японских журавлей на территории заповедника – это широкие, до 6 км, осоковые и вейниково-осоковые болота, расположенные вдоль русел рек Борзя, Ярчиха, Грязная и рядом с крупными ( 50 га) озерами – Долгое, Косое, Лебединые. В биотопах японского журавля основными компонентами растительных сообществ болота и переувлажненных лугов являются осоки и вейники, дополняющие компоненты – вахта трехлистная, сабельник и тростник обыкновенный. Размер индивидуального участка – 6-12 км2 (n = 7). Типичный гнездовой биотоп – это осоково-моховое, часто закочкаренное болото; кочки невысокие, плотные. Гнездовой участок маскируют заросли тростника или невысокого кустарника с разнотравьем. Кормовые биотопы используются в течение всего гнездового сезона, с учетом представленного в них пищевого разнообразия, а также доступности объектов питания. В гнездовой период у партнеров, чаще всего это самцы, кормовые стации могут находиться на расстоянии двух и более километров от гнезда и не входить в индивидуальный участок. Смена кормовых биотопов обуславливается не столько истощением пищевых ресурсов, сколько появлением других объектов питания в большем количестве и доступных в добывании. Предпочтительные места кормежки – это осоково-вейниковое и вейниково-осоковое пушицевое болото, прирусловые участки рек, береговая линия озер. В послегнездовой период к перечисленным биотопам добавляются осоково-вейниковые разнотравные луга и осоково-разнотравные увлаженные луга. Пребывание журавлей в увлажненных биотопах летом составляет 70,3% встреч.
Территориальное распределение. Известные территориальные участки японских журавлей на Архаринской низменности располагались в пределах территорий заповедника и заказника по пойменным болотам и на обширных осоковых болотах в понижениях микрорельефа. Только единичные пары гнездились на сопредельных ВБУ. Плотность гнездования на низменности (ВБУ составляют 583,1 км2) в среднем составила 0,04 пары/км2. Территории распределены довольно плотно. Расстояние между близко расположенными территориальными участками составило менее 0,5 км, удаленными – около 2 км.
В Антоновском лесничестве (в 16 км от р. Амур) выделены 16 территориальных участков. Плотность (ВБУ составляют 101 км2) поселения журавлей в годы минимальной численности – 0,04 пары/км2, максимальной – 0,11 пары/км2. Основное размещение территорий журавлей затрагивало центральную часть филиала: вдоль рек Борзя и Ярчиха и около системы старичных озер в юго-западной части, в стороне Амура. Территориальные участки, расположенные в понижении микрорельефа журавли занимали 2-18 сезонов. Естественное воспроизводство журавлей на участках в понижении микрорельефа имело хорошие показатели – 1,66 ± 0,09 птенца/пару (n = 29). Предпочтительный гнездовой биотоп (n = 72) – вейниково-осоковое пушицевое болото (98,6%) в окружении осоково-вейниковых и осоково-разнотравных лугов. Лесистость на территориальных участках не выражена. Гнездовые территории заселяли по 1-2-3-4 сезона подряд и с интервалами в 2-4-5-6-7 лет. Средний срок заселения – 10 лет. Отмечено занятие территориального участка с 1971 г., однако, не каждый год. Самый высокий процент использования журавлями территории филиала отмечен в 1986 г. – 68,7%, 11 территориальных пар.
В Лебединском лесничестве (ВБУ составляют 169 км2) выделено 14 территориальных участков. Они равномерно распределены по всему лесничеству, не перекрывая друг друга. Высокая плотность гнездовых территорий отмечена в западной части лесничества. По крупным водотокам – рекам Грязная, Урил и озерам Кривое, Лебединые, Хомутиное – сосредоточено 57,1% территорий. Места обитания – широкие открытые пространства болот и лугов. Плотность поселения при минимальной численности – 0,024 пары/км2, минимальной – 0,065 пары/км2. Предпочтительный гнездовой биотоп (n = 26) – вейниково-осоковое пушицевое болото (84,6%), рядом находятся осоково-вейниковые заболоченные и осоково-разнотравные луга. Журавли использовали территориальный участок с периодичностью 1-2-3-4 сезона подряд и интервалами в 2-3-4-5-6-11 лет. Максимальная продолжительность – 17 и 18 лет. Средний размер выводков на продуктивных участках – 1,72 ± 0,28 птенца/пару (n = 42). По среднему показателю гнездовые территории использовали 9 лет (2 участка). В 1986 г. 78,6% территориальных участков были заселены журавлями, обычно – 42,8%.
В заказнике «Ганукан» максимальное количество территориальных пар отмечено в 1986 г. – 12 пар. Средний показатель воспроизводства – 1,0 птенца/пару (n = 20).
Территориальное распределение японских журавлей на Архаринской низменности связано с распространением вейниково-осоковых болот, которые используются журавлями для гнездования в более 80% случаев. Все продуктивные территориальные участки находятся только внутри охраняемой территории, в низинной местности по равнинным рекам и около крупных старичных озер. Выявлено, что погодные условия года не влияют на территориальное распределение в сезон размножения. Большее влияние оказывают пожары. Гнездовые территории используются разными парами, несмотря на имеющийся гнездовой консерватизм в выборе территории и места для гнезда. Это подтверждают наблюдения за окольцованными птицами (n = 4).
Динамика численности территориальных пар. На двух равнинных участках заповедника в 1981-2000 гг. зарегистрировали пребывание 289 территориальных пар, в том числе 81 пара (28%) не гнездилась или гнездование было неуспешным. В годы учетов в заказнике отмечено 58 территориальных пар. Пар без потомства отмечено больше в филиале – 34%, в Лебединском лесничестве этот показатель равен 20,5%. Средний показатель нахождения территориальных пар в заповеднике равен 14,4 пары/год (n = 289).
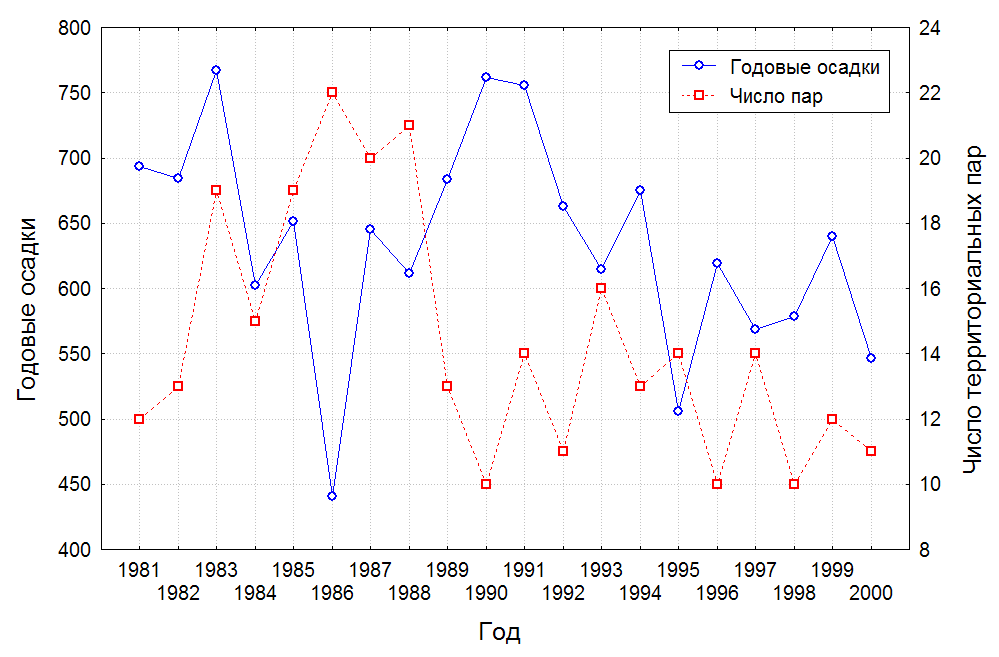
Рис. 3. Динамика численности территориальных пар японских журавлей в Хинганском заповеднике и ее зависимость от количества среднегодовых осадков.
Численность территориальных пар журавлей не оставалась постоянной. В динамике выявлены два больших периода, отличающиеся своими амплитудами (рис. 3). Первый охватывает 1981-1990 гг., когда наблюдался быстрый практически двукратный рост численности с 12 до 22 территориальных пар в 1986 г., и с таким же резким падением до 10 пар в 1990 году. Второй период, с меньшей амплитудой, начался с небольшого роста в 1991 г., в 1993 г. достиг максимума в 16 территориальных пар и к 2000 г. вновь сократился до 11 пар. Оба периода изменения численности территориальных пар укладываются в 9-летний интервал, но возможно их продолжительность немного больше, т.к. наши исследования могли не захватить начало первого и конец второго периодов. Численность территориальных пар японских журавлей в 1956-1971 гг. по оценке Н.С. Панькина и И.А. Нейфельдт (1976) составляла не более 15 пар, на гнездовании в годы полевых исследований 1956-1957, 1960, 1970-1971 гг. они отмечали 2-4 пары. По данным С.В. Винтера (1979) численность японского журавля на Буреинско-Архаринской низменности в 1974-1976 гг. составляла 8 территориальных пар (5 гнездящихся). В Летописи природы Хинганского заповедника указано 5 пар японского журавля в 1980 г., т.е. отмечено ее некоторое падение, а с 1981 г., (рис. 3), численность начала расти. Данные наземных учетов в предыдущие годы свидетельствуют о стабильно низкой численности до 1980 г. с тенденцией к сокращению. Данные авиаучета в 2003 г. тоже показывают на продолжающееся падение численности журавлей на низменности. В 2003 г. на территории заповедника учтено 7 территориальных пар японских журавлей (5 размножающихся) и 6(5) в заказнике. В сравнении с 2000 г. (17 территориальных пар) отмечено небольшое падение численности журавлей, амплитудный характер изменения численности популяции японского журавля на Архаринской низменности подтверждается другими исследованиями. Максимальное число территориальных пар в заповеднике составило 22 пары в 1986 году, минимальное – 10 пар в 1990 и 1998 годах, падение численности в 2,2 раза. В виду низкой численности журавли использовали, имеющиеся на низменности гнездовые территории, только на 33,3-73,3%.
Изменение численности размножающихся пар японских журавлей в заповеднике повторяло динамику численности территориальных пар. Также выделены два больших амплитудных периода подъема и падения численности. В период 1981-1986 гг. число размножающихся пар увеличилось с 11 до 19 пар; после короткого периода отмечено падение до 8 размножающихся пар в 1989 г. Падение численности размножающихся пар продолжилось до 2000 г., численность сократилась в 2 раза. На численность пар, не принимавших участие в размножении или размножавшихся неуспешно, в первую очередь, влияние оказывали пожары, потеря кладок и т.п. В среднем, в 1980-2000 гг. в заповеднике обитало 5,5 ± 4,5 территориальные пары без потомства (6,9%-75,9%).
Следует, что численность японских журавлей на Архаринской низменности нестабильна с тенденцией к общему сокращению. При этом места обитания журавлей не деградируют и не сокращаются, т.к. находятся под охраной. Объяснение нестабильному состоянию популяции этого вида на низменности находится в общем состоянии материковой популяции японского журавля, а также влиянии природных и антропогенных факторов. В годы с хорошим и достаточным увлажнением (1980-1991) численность территориальных пар была высокой, в сухие и засушливые годы (1993, 1995-2000) происходило падение количества территориальных пар в 2,2 раза от максимального количества. Исключение составляет 1986 г., когда при низких показателях среднегодовой суммы осадков (440 мм/год) отмечен максимальный рост числа территориальных пар журавлей. Недобор по осадкам в 1986 г. пришелся на осенний период, а в предыдущие годы количество осадков было выше среднего, поэтому погодные условия года не отразились на размножении журавлей. Хотя корреляция между численностью журавлей и суммой годовых осадков нами не выявлена. Пирогенный фактор серьезно влиял на численность размножающихся пар журавлей. Весенние пожары 1983, 1985, 1987, 1989, 1994 годов, прошедшие по территории заповедника, погубили 18 гнездовых участков, включая гнезда с кладками. В 1985 г., из-за пожаров число пар, не принимавших участие в размножении, изменилось с 0 до 45%. В исследуемом периоде у 6,2% территориальных пар размножение оказалось неуспешным, по причине пирогенного фактора.
Приведенный анализ свидетельствует о стабильно низкой численности японских журавлей на Архаринской низменности. Емкость угодий низменности используется журавлями менее чем на 50%, поэтому теоретически при общем росте материковой популяции можно ожидать рост численности на низменности до 40 и более пар. В годы максимальной численности на Архаринской низменности обитало 34 пары (32 размножались).
Демографические показатели популяции. В заповеднике число успешно размножающихся пар в благоприятные для размножения годы составляло 90-100%, в худшие годы 28,6-36,4%. Средний размер кладок – 1,98 яйца/пару (n = 135). Оплодотворенность яиц (по встречам выводков по 2 птнеца и результатам инкубации природных кладок) – 100%. Кладки с двумя неоплодотворенными яйцами находили только два раза. Такие кладки мы относим к повторному размножению, но не исключаем и другие причины. Данных об эмбриональной смертности в природных кладках японских журавлей у нас нет. Средний размер выводков – 1,5 птенца/пару (n = 162), т.о. смертность птенцов до подъема на крыло в исследуемом нами периоде составила 25%. Нами не обнаружена корреляция между размером выводка и суммой годовых осадков (рис. 4). Однако смертность птенцов в засушливые годы отмечена выше в 1,5-2,2 раза, по сравнению с годами хорошего увлажнения (табл. 3). Гибель кладок регистрировали при пожаре (8), затоплении гнезда (1), разорении воронами (2), прерывании инкубации птицами (1), по неизвестным причинам (1).
Таблица 3.
Влияние количества осадков на размножение японских журавлей
в Хинганском заповеднике
| Показатели | Осадки больше 660 мм/год (n = 7) | Осадки 560-660 мм/год (n = 10) | Осадки меньше 560 мм/год (n = 3) |
| Средний размер выводка | 1,6 (n = 61) | 1,42 (n = 78) | 1,5 (n = 23) |
| Потенциальный размер выводка* | 2,0 | 2,0 | 2,0 |
| Смертность в выводках, % | 20 | 29 | 25 |
| Доля размножающихся пар, % | 79,8 | 70,9 | 59,6 |
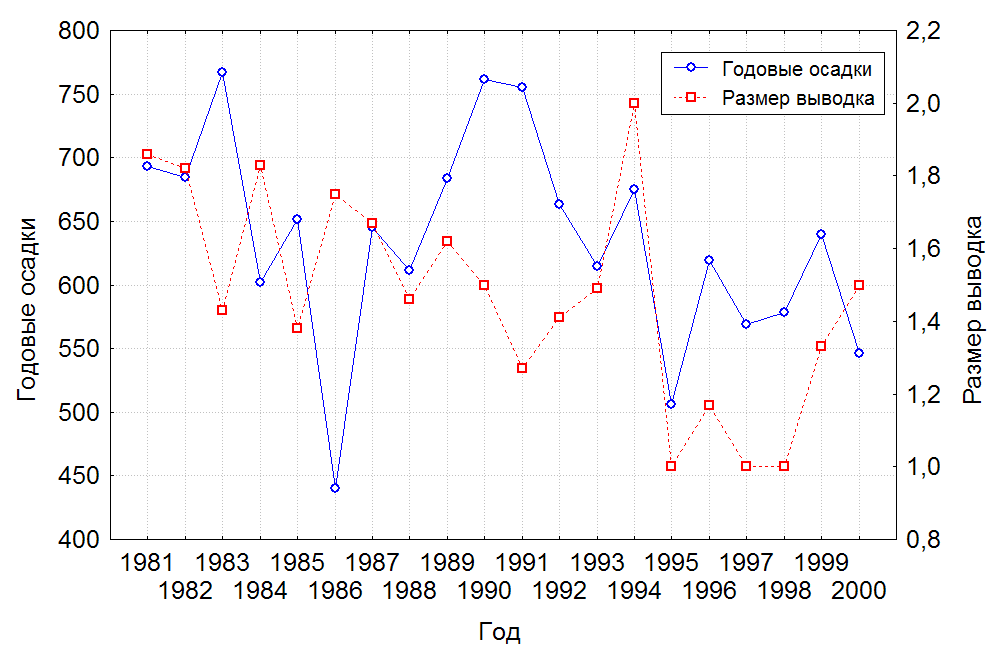
Рис. 4. Влияние годовой суммы осадков на размер выводка у японских журавлей на Архаринской низменности.
Основной отход, по наблюдениям за судьбой выводков, происходит в первый месяц жизни. Анализ судьбы гнезд (n = 95) за 1983-1991 гг. показывает, что смертность птенцов в мае-июле составляет 0-45%.
Возрастная структура популяции японских журавлей по данным авиаучетов осенью выглядит следующим образом: численность птиц старше 1 года относительно птенцов выше в 1,6-3,9 раза. Доля птенцов в популяции осенью не превышает 40%. Исходя из данных о численности журавлей в заповеднике осенью, возрастная структура (отношение половозрелых птиц к неполовозрелым и птенцам) японского журавля выглядит как 1 : 0,67 ± 0,43 : 0,55 ± 0,25, отношение взрослых птиц к птенцам в среднем 1: 0,3.
Продолжительность жизни японских журавлей в природе, по данным кольцевания, составляет 13 лет. Журавль был помечен птенцом в 1991 г. в заповеднике и последняя информация о его встрече на зимовке в Китае датируется 2004 г. Гибель взрослых птиц в природе отмечали 3 раза. Причины гибели – травмы.
Демографические показатели у японских журавлей Архаринской низменности, в целом, совпадают с данными по другим участкам ареала (Шибаев, Глущенко, 1988; Горошко, 2005; Feng Kemin, Zhao Zhongqin, 1991; Willims et al., 1991; Li Xiao-Min, 2000; Lee, 2004).
Гнездовая и послегнездовая жизнь. Приведено описание поведения птиц в гнездовой и послегнездовой периоды, описание гнезд, кладок; приведены данные о суточной активности, взаимоотношениях взрослых и птенцов в семье.
Питание. В питании журавлей соотношение растительной и животной пищи в различные периоды сезона размножения меняется, что вызвано доступностью и обилием питания в кормовых стациях. Преобладает в питании животная пища. Основные способы добывания корма: склевывание и зондирование.
Миграции и места зимовок. Спутниковое слежение за перемещениями журавлей в гнездовой части ареале показало, что в 1998 г. каждая из отловленных нами птиц пребывала на одном участке, диаметром примерно в 10-15 км. В 1999 г. отмечено передвижение журавля в сторону г. Благовещенск (240 км), где птица оставалась до 20.09., после чего с 22.09. по 11.11. вновь находилась в месте отлова. Другой журавль, с места отлова 13.07. на р. Грязная (Хинганский заповедник) к 17.10. переместился на запад, на территорию заказника «Ганукан» (50-60 км). Похожие перемещения и пребывание на площадях большого диаметра в гнездовом ареале мы отмечали у неполовозрелых и неразмножающихся японских журавлей, выпущенных в природу.
Осенний отлет птиц происходит практически в одни сроки: один журавль стал перемещаться в сторону зимовки 10.11.1999 г., другой – 11.11.1999 г. Определены два основных миграционных коридора на северо-востоке Китая, который используют японские журавли от гнездовых территорий в бассейне р. Амур к побережью Ляодунского залива: один проходит по бассейну р. Нэньцзян (западный), и другой, по бассейну р. Сунгари (восточный). Два журавля, помеченные в 1993 г., летели западным путем. Начало их миграции отмечено в первой декаде ноября. Этим же путем летели и журавли в 1999 г. Восточный путь используют журавли с оз. Ханка.
В начале миграции птицы улетели в провинцию Хэйлунцзян (Китай), где отдыхали не более двух суток. Оттуда они полетели в сторону залива Бохай (Bohai Bay), где оставались с 7 по 15 ноября. Журавли отдыхали в заливе Бохай – 6-7 дней. Следующая остановка и отдых отмечены на побережье Ляодунского залива (Liaodong Bay) около г. Тангшан, здесь оставались между 9 и 17 ноября. От побережья Ляодунского залива птицы перемещались вдоль залива Бохай к дельте Желтой реки (Yellow River). Один из меченых журавлей отдыхал здесь 3 дня, до отлета на юг; второй оставался в течение 25 дней. В 1998 г. в этом месте один журавль сменил направление и полетел к эстуариям рек Ханган и Имджинган (Han/Imjin estuary) на Корейском полуострове. От дельты Желтой реки в 1993, 1998 и в 1999 гг. журавли полетели к морскому побережью Яньчэн (Yancheng N.R.), (Higuchi et al., 1998; Tamura et al., 2000). Средняя протяженность миграционного пути составила 2241,8 км. Продолжительность миграции 6-39 дней, в среднем – 24 суток. Западный миграционный путь в 2,5 раза протяженнее, чем восточный, его используют журавли с Приханкайской равнины.
На зимовке птицы чаще находились на сельскохозяйственных землях, чем на водно-болотных угодьях. Только 19,7% времени журавли проводили на водно-болотных угодьях в Китае. В местах гнездования в бассейне Амура они предпочитали водно-болотных угодья.
Весенняя миграция начинается в первых числах марта. На перелет к местам гнездования помеченный спутниковым передатчиком журавль затратил 34 дня и появился на Архаринской низменности 4 апреля.
Журавли, помеченные нами на Архаринской низменности, отмечались также и в Янченге (Китай).
Глава 5. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА БИОЛОГИИ ДВУХ ВИДОВ ЖУРАВЛЕЙ НА АРХАРИНСКОЙ НИЗМЕННОСТИ
Появление двух видов журавлей на низменности определяют погодные условия, сроки прилета отличаются в среднем на 7 дней. Прилет японских журавлей связан с переходом среднесуточных температур приземного воздуха через 0°С к положительным значениям, даурских журавлей – через +5°С к высоким значениям.
Даурские журавли обитают на открытых слабо увлажненные пространства вейниково-осоковых болот и осоково-разнотравных лугов, на возвышениях микрорельефа местности (100-124 м над ур. м.). Индивидуальные территории даурских журавлей обязательно сочетают естественную среду обитания с разной степенью нарушенную человеком и сельхозугодия. 81,8% гнездовых участков располагается по периферии охраняемой территории и не охватывает водно-болотные угодья центральной части ООПТ.
У японских журавлей предпочтительным биотопом являются обширные вейниково-осоковые и осоково-пушицевые болота. Такие биотопы располагаются по руслу равнинных рек и вблизи крупных старичных озер, в понижениях микрорельефа местности (93-107 м над ур. м). Распределение гнездовых территорий японских журавлей на низменности показывает, что 73,3% участков находятся на особо охраняемых природных территориях, в центральной их части. Плотность гнездования японских журавлей на низменности в 2 раза выше, чем у даурских. Показатели размножения у японских журавлей также выше: средний размер выводков составляет 1,5 птенца/пару, у даурских журавлей – 1,37 птенца/пару.
Динамика численности японских журавлей имеет амплитудный характер с тенденцией к постепенному сокращению. Численность даурских журавлей имеет положительный тренд. Изменения численности журавлей на низменности связаны с общим состоянием популяций в мире. Имеются отличия и в биологии гнездового и послегнездового периодов у этих видов журавлей. Отличаются пути миграций и места зимовок. Благополучные условия на зимовках даурских журавлей на о. Кюсю в Идзуми, Япония, и на ДМЗ, Корея, способствуют восстановлению и росту популяции. Зимовки японских журавлей находятся, главным образом, в Китае. Проблемы на зимовках и в гнездовом ареале не способствуют росту и стабильности материковой популяции японского журавля.
Глава 6. УГРОЗЫ И МЕРЫ ПО СОХРАНЕНИЮ ЖУРАВЛЕЙ НА АРХАРИНСКОЙ НИЗМЕННОСТИ
В главе приводятся меры по сохранению журавлей на Архаринской низменности. Важными мерами сохранения журавлей и среды обитания считаем:
1. Активизация международного сотрудничества по сохранению журавлей и среды их обитания.
2. Осуществление постоянного мониторинга за отдельными минипопуляциями и за изменениями среды их обитания в местах гнездования и на зимовках.
3. Для снижения фактора беспокойства и поддержания численности двух видов журавлей могут способствовать работы, которые проводятся на Станции реинтродукци редких видов птиц Хинганского заповедника: реинтродукция потомства, создание популяций «полудиких птиц» в природе.
4. Снижение негативного влияния пирогенного фактора на гнездование журавлей через проведение контролируемых отжигов в сезоны, не связанные с размножением птиц (начало весны, осень).
5. Страхование популяций и среды их обитания в зоне возможного экологического ущерба.
6. Обязательное проведение компенсационных мероприятий при хозяйственном использовании или утрате мест гнездования журавлей.
7. Создание цепи зоопарки-питомники-заповедники, которые позволят осуществлять программы по реинтродукции потомства журавлей в природу.
8. Необходимо продолжать резервировать локальные места обитания журавлей, создавать дополнительные и разнообразные категории ООПТ.
9. Необходимо более глубокое изучение биологии видов, чтобы понять их возможную реакцию на те или иные изменения экологических условий в местах обитания и стремиться к нейтрализации выявленных лимитирующих факторов.
10. Чрезвычайно важно усиление роли целевой пропаганды (День журавля, Музей журавля, Парк журавля и т.п.) в отношении охраны журавлей.
ВЫВОДЫ
- На Архаринской низменности выделено 35 территориальных участка даурских журавлей и 42 японских журавлей. Заселение журавлями всех участков еще не наступило. Плотность гнездования японских журавлей на низменности превышает плотность гнездования даурских журавлей почти в 2 раза. Некоторые участки заняты журавлиными парами 30 лет.
- Прилет и отлет журавлей имеет характерную для вида достоверную зависимость с температурой воздуха или с доступностью корма.
- Распределение двух видов журавлей на низменности имеет существенные отличия: большинство участков даурских журавлей располагаются по границе водно-болотных угодий и сельхозугодий, а японских журавлей – исключительно во влажных биотопах, приуроченных к крупным водотокам.
- Гнездовой консерватизм у журавлей выражен, однако, при определенных причинах могут менять территориальные участки.
- Показатели размножения журавлей изменяются в зависимости от погодных условий года, прохождения пожаров по гнездовой территории и других факторов. Погодные условия больше отражаются на гнездовании японских журавлей, тогда как даурские журавли менее зависимы от этого фактора. Размер выводков у даурских журавлей больше зависит от погодных условий, чем у японских журавлей. Смертность птенцов до подъема на крыло у даурских журавлей – 29,3%, у японских журавлей – 25%.
- Зимовки даурских журавлей с Архаринской низменности находятся на о. Кюсю (Япония) и демилитаризованной зоне, в Корее. Зимовки японских журавлей находятся в Яньчэне, Китай. Состояние миграционного пути и зимовок даурских журавлей не вызывает опасений и способствуют восстановлению и росту популяции, в отличие от зимовок и мест остановок японских журавлей, где отмечается серьезная деградация среды обитания.
- У двух видов журавлей практически отсутствует межвидовая конкуренция по причинам: разные сроки прилета и отлета, использование разных биотопов в послегнездовом периоде, различный спектр питания, отличия в биологии послегнездового периода и др.
- Основная часть гнездовых территорий японских и даурских журавлей находится на особо охраняемых территориях, что обеспечивает защиту и сохранность среды обитания журавлей. Главными лимитирующими факторами журавлей в местах гнездования являются фактор беспокойства, пирогенный и климатический факторы, антропогенная трансформация биотопов (ГЭС, осушение).
- Сохранение среды обитания журавлей на Архаринской низменности требует введения дополнительных природоохранных мер таких, как проведение контролируемых отжигов для защиты гнездовий журавлей, создание устойчивых «полудиких популяций», продолжение работ по реинтродукции, резервирование локальных мест обитания, ведение постоянного мониторинга на ключевых местах и т.п.
СПИСОК ОПУБЛИКОВАННЫХ РАБОТ ПО ТЕМЕ ДИССЕРТАЦИИ
Статья, опубликованная в ведущем рецензируемом научном журнале:
- Hiroyoshi Higuchi, Johanna P. Pierre, Vladimir Krever, Vladimir Andronov, Go Fujita, Kiyoki Osaki, Oleg Goroshko, Mutsuyuki Ueta, Sergei Smirenski, Nagahisa Mita. 2004. Using a Remote technology in conservation: Satellite tracking white-naped Сranes in Russia and Asia // Conservation Biology, Vol. 18, №1: 136-147.
Статьи, опубликованные в других научных изданиях:
- Андронов В.А. 1983. Состояние популяций журавлей и аистов в Хинганском заповеднике в 1982 г. // Охрана живой природы. М.: 8-10.
- Андронов В.А. 1988. Современное состояние японского Grus jаponensis (P.L.S. Muller) и даурского Grus vipio Pallas журавлей в Амурской области // Журавли Палеарктики. Владивосток: 187-190.
- Андронов В.А. 1990. Редкие виды птиц в Хинганском заповеднике // Заповедники Амурской области. Благовещенск, 1990: 94-102.
- Андронов В.А. 2001а. Японский (уссурийский) журавль Grus japonensis (P.L.S. Muller, 1776) // Красная книга Российской Федерации. М.: Астрель: 468-469;
- Андронов В.А. 2001в. Даурский журавль Grus vipio Pallas, 1811 // Красная книга Российской Федерации. М.: 472-473.
- Андронов В.А., Андронова Р.С., Петрова Л.К. 1988. Территориальное распределение японского журавля на Архаринской низменности // Журавли Палеарктики. Владивосток: 59 – 62.
- Андронов В.А., Андронова Р.С. 2005. Новая информация о встрече меченых журавлей на зимовках в Японии, Китае и Корее // Информационный бюлл. РГЖ. Вып. 9. М.: 91.
- Андронов В.А., Дарман Ю.А., Парилов М.П., Нагендран М.,Хигучи Х. 2001. Состояние популяций журавлей и аистов Амурской области в 1998 г. // Орнитология, Вып. 29. М.: 314-319.
- Антонов А.И., Кузнецова Т.А., Андронов В.А. 2005. Архаринская низменность // Водно-болотные угодья России. Том 5. Водно-болотные угодья юга Дальнего Востока России (под общ. ред. В.Н. Бочарникова). М.:Wetlands International: 54-57.
- Дарман Ю.А., Андронов В.А. 1987. Результаты учета японского журавля в Хинганском заповеднике // Сообщения Прибалтийской комиссии по изучению миграции птиц. Вып. 19. Тарту: 107-111.
- Дарман Ю.А., Андронов В.А., Хигучи Х., Нагендран М., Тамура М., Горобейко В.В., Росляков А.Г., Парилов М.П. 2000. Авиаучет редких видов птиц на Средне-Амурской низменности в 1999 г. // Дальневосточный аист в России. Владивосток:: 13-19.
- Смиренский С.М., Андронов В.А. 1988. Первые итоги мечения даурских журавлей // Журавли Палеарктики. Владивосток: 193-194.
- Andronov V. A. 1989. The Strategy of Rare Bird species Preservation in the Middle Part of the Amur Basin // Geology and Ecology of Amur Basin (Vol. 4). Blagoveshchensk: 18-19.
- Smirensky S. M., Andronov V.A., and Roslyakov G.E. 1985. Distribution of Red-crowned Cranes (Grus jponensis) in Priamurye in 1984 Breeding Season // International spring censuses of Grus japonensis (Red-crowned Crane) in 1984. Ed. H. Masatomi. Tokyo, Wild Bird Society of Japan: 3-7.
- Higuchi H., Shibaev Y., Minton J., Osaki K., Surmach, S., Fujita G., Momose K., Momose Y., Ueta M., Andronov V., Mita N. and Kanai Y. 1998. Satellite tracking of the migration of the Red-crowned Crane Grus jаponensis // Ecological Research (1998) 13: 273-282.
- Tamura M., Higuchi H., Shimazaki H., Oguma H., Darman Yu., Andronov V., Nagendran M., Parilov M. 2000. Satellite observation on Movements and habitat conditions of Red-crowned crane and Oriental white stork in East Asia // Global Environmental Research. Tokyo, Vol.4, No.2: 207-217.
- Hiroyki Masatomi, Sergei Smirenski, Кunikazu Momose, Kimiya Koga,Vladimir Andronov, Yuri Darman, and Yulia Momose. 2002. Status of cranes and storks breeding in the middle Amur River basin, Russia, in the summer of 1997 // Biosphere Conservation, 4 (2): 87-102.
Работы, опубликованные в материалах региональных, всероссийских, всесоюзных и международных конференций:
- Андронов В.А. 1986. О создании полудикой популяции японского журавля // Первое Всесоюзное совещание по проблемам зоокультуры. Ч. 2. М.: 7-8.
- Андронов В.А., Андронова Р.С. 2007. Пути сохранения популяций редких птиц в зоне влияния Бурейской ГЭС // Материалы науч. конф. по заповедному делу. Благовещенск, 3-8 октября 2007 г. Благовещенск: 43-47.
- Андронова Р.С., Андронов В.А. 2007. Расселение японских журавлей в гнездовом ареале на примере реинтродуцированных в природу // Тезисы докладов, III Международная конференция по мигрирующим птицам Севера Тихоокеанского региона. Ч. I. Якутск: 8-10.
- Дарман Ю.А., Андронов В.А. 1997. Охрана водно-болотных угодий и редких видов околоводных птиц Амурской области // Тезисы докладов областной научно-практической конференции (Новиковские чтения), посвященной 140-летию образования в Амурской области. Благовещенск: 100-105.
- Andronov V.A., Shibaev Yu.V. 1995. The status of the Red-crowned Crane in former USSR // Crane research and protection in Europe. The proceedings of the international crane workshop. Moscow: 325-326.
- V. Andronov, R. Andronova, E. Ilyashenko, O. Goroshko, M. Parilov. 2008. Status of rare cranes in eastern Siberia // The Current Status and Issues of the Red-crowned Crane; Proceedings of the meeting «Establishment of a Feasible International Project for Protection of the Tancho Grus japonensis in 2007, Hokkaido, 18-23 November 2007:83- 86.
- Higuchi H., Osaki K., Golovushkin K., Goroshko O., Krever V., Minton J., Ueta M., Andronov V., Smirenski S., Ilyashenko V., Kanmuri N. and Archibald G. 1994. The migration routes and important rest-sites of cranes satellite tracked from south-central Russia // The future of cranes and wetlands. Proceeding of the International Symposium Ed. H. Higuchi and J. Milton. Tokyo, Japan: 15-25.
- Darman Yu.A., Andronov V.A. 1998. A commentary on the wetland conservation efforts and status of endangered waterbirds of the Amur Region, Russia // Wetland and waterbird conservation in North East Asia. Beidaihe, China: 265-268.
АНДРОНОВ Владимир Андреевич
Японский (Grus japonensis P.L.S. Mller, 1776) и даурский
(Grus vipio Pallas, 1811) журавли Архаринской низменности
(Среднее Приамурье)
АВТОРЕФЕРАТ
Лицензия №….. от ……..
Подписано к печати 01.11.2008 г. Формат 6084 1/16. Уч.-изд. л. 1.0.
Печать RISOGRAPH TR 1510. Тир. 100 экз. Заказ №….
Отпечатано с оригинала заказчика …….