

Цитоскелетные компоненты мышечных волокон и коллаген в условиях реальной и моделируемой гравитационной разгрузки
На правах рукописи
ЧИСТЯКОВ ИЛЬЯ НИКОЛАЕВИЧ
ЦИТОСКЕЛЕТНЫЕ КОМПОНЕНТЫ МЫШЕЧНЫХ ВОЛОКОН И КОЛЛАГЕН В УСЛОВИЯХ РЕАЛЬНОЙ И МОДЕЛИРУЕМОЙ ГРАВИТАЦИОННОЙ РАЗГРУЗКИ
03.03.01 – физиология
03.03.04 – гистология, цитология, клеточная биология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата медицинских наук
Москва 2010
Работа выполнена в Государственном научном центре Российской Федерации – Институте медико-биологических проблем Российской академии наук
НАУЧНЫЙ РУКОВОДИТЕЛЬ:
доктор биологических наук, профессор Шенкман Борис Стивович
ОФИЦИАЛЬНЫЕ ОППОНЕНТЫ:
доктор медицинских наук, профессор Оганов Виктор Сумбатович
доктор медицинских наук, заместитель директора, профессор Маевский Евгений Ильич
ВЕДУЩАЯ ОРГАНИЗАЦИЯ: Факультет фундаментальной медицины Московского государственного университета им. М.В.Ломоносова
Защита диссертации состоится « » ______ 2010 г. в часов на заседании диссертационного совета Д 002.111.01 при Государственном научном центре Российской Федерации – Институте медико-биологических проблем Российской академии наук по адресу 123007, Москва, Хорошевское шоссе, д.76-А.
С диссертацией можно ознакомиться в библиотеке ГНЦ РФ - ИМБП РАН.
Автореферат разослан «___» ___________ 2010 г.
Ученый секретарь диссертационного совета,
доктор биологических наук Левинских М.А.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы:
Увеличение длительности космических экспедиций и расширение круга решаемых в полете двигательных задач повышает значение бортовой профилактики, прогнозирования эффектов гравитационной разгрузки и борьбы с двигательными нарушениями. Необходимость контроля и сохранения нормальной функции скелетных мышц требует комплексного исследования физиологических механизмов и закономерностей адаптационных процессов, обеспечивающую функциональные изменения, адекватные изменению биомеханической среды. Согласно этическим нормам [Генин А.М. соавт. 2001], человека нельзя подвергать неблагоприятным воздействиям, что затрудняет корректный анализ феноменологии процессов и физиологических механизмов атрофии мышечной ткани человека. Экспериментальные исследования на животных позволяют провести более детальный анализ исследуемых процессов и изучить механизмы структурно-функциональных перестроек, с применением более жестких, чем это возможно для человека, методов воздействия и методик исследования [Booth F.W. 1982; Fitts R.H. 1986; Belozerova I.N. 2002; Adams G.R. et al. 2003].
Известно, что у человека после космического полета наблюдается снижение функциональной активности скелетных мышц [Oganov V.S. et al. 1976; Козловская И.Б. соавт. 1984; Martin T. 1988; Leach C.S. 1992; Shenkman B.S. et. al. 1994, 1996; Edgerton V.R. et al 1998; Adams G.R. et al. 2003]. В литературе описаны такие изменения скелетных мышц, вызванные гравитационной разгрузкой, как снижение объема мышцы [Dapp K. et al., 1989; Kozlovskaya I. B. et al. 1996; Desplanсhes D. et al. 1997], избирательное уменьшение площади поперечного сечения (ППС) мышечного волокна [Оганов В.С. соавт. 1982; Riley D.A. et al. 2002], изменения спектра изоформ тяжелых цепей миозина в пользу миозина быстрого типа [Takacs et al. 1980; Caiozzo V.J. et al. 1994], разнонаправленные изменения в сети десминовых филаментов [Nara K. et al. 2002], нарушения целостности дистрофинового слоя сарколеммальной мембраны [Гасникова Н.М. соавт. 2006; Chopard A. et al. 1998, 2005], увеличение объёма экстрацеллюлярного матрикса [Karpakka J. et al. 1991; Miller T.D. et al. 2001; Kjaer M. 2004]. Данные современных исследований качественных и количественных изменений белков внутриклеточного цитоскелета и экстрацеллюлярного матрикса постуральных мышц в условиях микрогравитации по-прежнему остаются единичными.
Архитектура скелетно-мышечной клетки определяется балансом сил напряжения внутреннего актинового цитоскелета, костамеров и внеклеточного цитоскелета, формирующих систему самоорганизующихся напряженных модулей и обладающих координированной активностью. Действие микрогравитации на мышечную клетку реализуется через ряд механизмов, которые можно условно разделить на внешние системные, (изменение биомеханики движений, отсутствие опоры и снижение сопротивления среды, нейрорегуляторные и гормональные воздействия) и внутренние местные, (накопление ионов Са2+ в мышечном волокне, нарушение процессов белкового синтеза, усиление протеолиза и др.). Гравитационно-зависимые изменения мышечных белков носят комплексный характер, затрагивающий все структурные уровни, и приводят к компенсаторной адаптации мышцы [Oganov V.S. et al. 1980, 1981, 1982, 1990; Edgerton V.R. et. al. 1998; Fitts R.H. et. al. 2000]. Состояние постуральных скелетных мышц во многом зависит от изменений компонентов саркомера (актин, миозин, титин, небулин, белки М-линии), внесаркомерного цитоскелета (десмин), сарколеммальных белков, формирующих костамеры (дистрофин) и экстрацеллюлярного матрикса (интегрин, коллаген).
На сегодняшний день нет ясного понимания физиологической роли промежуточных филаментов (ПФ) десмина в работе скелетных мышц в нормальных условиях. Известно, что экстрамиофибриллярный цитоскелет десмина, опосредованно через синкоилин и десмуслин/сименин, крепится к мембранным адгезивным комплексам, показана прямая связь между десмином и спектрином и анкерином, непосредственное взаимодействие десмина с белком интрафибриллярного цитоскелета небулином [Capetanaki Y. et al. 2002]. В исследованиях на десмин-/- мышах показано, что нарушение синтеза десмина приводит к разуплотнению саркомеров, нарушению латерального выравнивания миофибрилл и их связи с сарколеммой, потере формы и позиционирования ядра, дегенерации, нарушению локализации и изменению формы митохондрий, нарушению клеточной адгезии и расширению межклеточного пространства. Интересно, что в медленных мышцах концентрация десмина вдвое больше, чем в быстрых мышцах, что может определять физиологические особенности постуральных скелетных мышц. Однако, литературные данные, посвященные влиянию гравитационной разгрузки на свойства ПФ десмина в медленной (камбаловидной) мышце в настоящий момент остаются единичными и часто недостоверными [Chopard A. et al. 2005, Capetanaki Y. et al. 2007; Dapp et al. 2004, Nara K. 2002; Enns et al. 2007].
Сложность механизма регуляции коллагеногенеза, включающего перекрёстное влияние различных сигнальных путей ()TGF- и ()TNF-, факторов роста (IGF-1, b-FGF, VEGF, PDFG), простагландинов, цитокинов (Ил-1, Ил-6, Ил-8), NO, активации/подавления ММР-2/TIMP-1, ММР-9/TIMP-2 [Sakuma K. et. al., 2000; Kjaer M., 2004], не позволяет сформировать чёткой гипотезы гравитационно-зависимых изменений.
Целью настоящего исследования являлся экспериментальный гистоморфологический анализ изменений цитоскелетных и экстрацеллюлярных белковых структур мышц человека и животных при различной длительности гравитационной разгрузки, а также в условиях дополнительного механического и фармакологического воздействия.
Задачи исследования:
- Изучить динамику объёма десминовых и дистрофиновых структур мышечного волокна и объема коллагеновых структур I и III типа в скелетных мышцах животных на разных сроках моделируемой гравитационной разгрузки.
- Проверить гипотезу о роли накопления ионов кальция в изменении характеристик цитоплазматических и экстрацеллюлярных белковых структур в постуральных мышцах животных в условиях гравитационной разгрузки и возможность использования блокаторов кальция для предотвращения этих изменений.
- Проверить гипотезу об эффектах пассивного растяжения на коллагеновые и десминовые структуры в условиях моделируемой гравитационной разгрузки.
Научная новизна:
Определена динамика изменений компонентов внутриклеточного цитоскелета (десмина, дистрофина) и экстрацеллюлярного матрикса (изоформ коллагена I и III типа) МВ камбаловидной мышцы животных при разных сроках гравитационной разгрузки.
Выявлены качественные и количественные изменения в структуре изоформ коллагена экстрацеллюлярного матрикса МВ, увеличение доли изоформы коллагена Ш типа к поздним срокам гравитационной разгрузки.
Показана возможность частичного предотвращения увеличения общего объёма, изменения изоформного состава коллагена и сохранения дистрофинового слоя мышечного волокна камбаловидной мышцы крысы при 14-суточном антиортостатическом вывешивании, сочетанном с хроническим введением блокатора кальциевых каналов L-типа, нифедипина.
Показано, что пассивное растяжение на фоне антиортостатического вывешивания камбаловидной мышцы крысы позволяет частично предотвратить увеличение общего объёма и состава изоформ коллагена I и III типа.
Теоретическая и практическая значимость работы:
Настоящее исследование расширяет понимание физиологических и биохимических структурных изменений, происходящих в скелетной мускулатуре животных и человека при гравитационной разгрузке. Полученные результаты могут использоваться при разработке методов профилактики негативного воздействия невесомости. Разработанные иммуногистохимические (ИГХ) и топогистохимические (ТГХ) методы морфологического исследования цитоскелетных и экстрацеллюлярных белков скелетных мышц успешно используются клинической практике, в том числе в гинекологии и неврологии.
Положения, выносимые на защиту:
- Нарастание общего объёма коллагена в камбаловидной мышце при моделируемой гравитационной разгрузке прямо пропорционально её продолжительности и сопровождается увеличением доли изоформы коллагена III типа и уменьшением доли изоформы коллагена I типа.
- Адаптация системы промежуточных филаментов десмина к условиям гипогравитации происходит на ранних сроках гравитационной разгрузки, к 7-м суткам активность процесса адаптации существенно снижается.
- Пассивное растяжение мышцы и введение блокатора кальциевых каналов L-типа позволяет частично предотвратить деструкцию цитоскелетных сократительных и несократительных белковых структур в условиях гравитационной разгрузки.
Апробация работы:
Материалы диссертации доложены на IV, V, VI «Конференциях Молодых Учёных» (Москва ГНЦ РФ ИМБП РАН, 2004, 2005, 2007). На XXXIV European Muscle Conference, Hortobari, Hungary, 2005, P.17. На «XIII международном совещании» и «VI школе по эволюционной физиологии», Санкт-Петербург, 23-28.01.2006. На 27th Annual International «Gravitational Physiology Meeting», Osaka, Japan, 2006. На 16th IAA «Human in Space» Symposium, 21-24.05.2007. На VI Симпозиуме «Химия протеолитических ферментов», 2007, Москва.
Диссертация апробирована на секции Учёного совета 10.11.2009 г. ГНЦ РФ - ИМБП РАН.
Публикации:
По теме диссертации опубликовано 18 печатных работ, в том числе 3 в журналах рекомендованных ВАК.
Структура и объем диссертации:
Диссертация состоит из введения, обзора литературы, описания материалов и методов исследования, результатов, обсуждения, выводов, списка литературы. Текст диссертации изложен на 136 страницах, иллюстрирован 15 рисунками. Полученные данные представлены в 14 таблицах и 34 графиках. Список литературы включает 317 источника, из них 20 – работ отечественных и 297 - иностранных авторов.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Объектом исследования служили биоптаты мышечной ткани человека (4 участника экспедиции на МКС) и животных. В работе были использованы 194 половозрелых самца крыс линии Wistar, 24 самцов монгольской песчанки, 19 самцов мышей линии MDX и 19 самцов мышей линии C57black. Протоколы исследований с участием человека и эксперименты на животных были одобрены Комиссией по биомедицинской этике ГНЦ РФ – ИМБП РАН.
Методы исследования
Исследование экспериментального материала производилось с использованием микроскопа Leica. Анализ микропрепаратов проводился не менее чем на 100 волокнах каждого среза с помощью системы анализа изображений QUANTIMET-500 (Leica, Германия) с цветной цифровой видеокамерой Leica DC-300. Оценка анализируемых параметров проводилась на снимках высокого разрешения (Tiff) с использованием программного обеспечения Leica QWin.
Топогистохимическое выявление коллагена
Для выявления изоформ коллагена I и III типа использовалась гистохимическая реакция Picrosirius Red (Direct Red) с выявлением окраски фосфомолибденовой кислотой [Miller et al., 2001]. Объем общего коллагена на поперечном срезе определяется следующим образом: Доля площади коллагена I типа + доля площади коллагена III типа / ППС среза (рамка измерений). Процентное соотношение изоформ коллагена определяется по следующему уравнению: I (III) тип коллагена % = [ППС I (III) типа / (ППС I типа + ППС Ш типа)] x100.
Иммуногистохимическое выявление десмина
Окраска проводилась с использованием стандартного протокола двойного иммуномечения. При оценке количества десмин-негативных волокон производился прямой подсчёт всех волокон видимого среза. Рассматриваемые волокна считались десмин-негативными, если при окрашенном контуре волокно не имеет чёткого внутреннего окрашивания. Иммуногистохимическое выявление дистрофина
Окраска проводилась с использованием стандартного протокола двойного иммуномечения. Состояние дистрофинового слоя анализировали с использованием стандартизованного метода подсчёта процентного соотношения мышечных волокон с разрывами субсарколеммальной мембраны. В норме дистрофиновый слой выглядит как непрерывная линия, расположенная на внутренней границе волокна. В некоторых волокнах имеются участки, на которых целостность дистрофинового слоя нарушена.
Статистическая обработка
Оценку значимости различия средних значений показателя в независимых выборках проводили по t-критерию Стьюдента, который вычисляли по формуле t = M1 – M2 / (m12 – m22), где М1 и М2 – средние арифметические значения показателя в сравниваемых группах, m1 и m2 – средние квадратичные ошибки среднего арифметического значения показателя. В связанных выборках расчет t-критерия проводили по формуле t = М / mМ, где М – средняя разность показателя в сравниваемых группах, mМ – средняя квадратичная ошибка средней разности показателя. Различие средних значений показателя в сравниваемых выборках считали значимым при уровне значимости p0,05. Результаты представлены в виде М±m [Ю.В.Прохоров, 2003].
Организация экспериментов
Структура исследований, вошедших в диссертацию представлена в табл. 1
Таблица 1. Структура проведенных исследований
| № | Цели исследования | Условия | Объект | Объём исследований | |
| 1 | Исследование влияния реальной гравитационной разгрузки* на изоформный состав и объём белков цитоплазматического цитоскелета и экстрацеллюлярного матрикса скелетных мышц человека и животных. *-условий космического полёта | 1.Бортовой эксперимент «МИОН» в рамках экспедиций МКС № 6,7,8 (2002-2005 гг.) Продолжительность полёта составила 185, 193 и 188 суток. Биоптаты m.soleus человека. | 4 человека |
| |
| 2.Бортовой эксперимент «ФОТОН - М3» в рамках программы ФКА и РОСКОСМОС. Продолжительность полёта составила 12 суток. Биоптаты m.soleus и m. tibialis ant. монгольских песчанок. | 24 монгольских песчанки | ||||
| 2 | Определение динамики изменений изоформного состава и объёма белков цитоплазматического цитоскелета и внеклеточного матрикса скелетных мышц крысы на разных сроках гравитационной разгрузки. | Антиортостатическое вывешивание крыс 3-, 7-, 14-, 30- суток. 10 групп:
| 72 крысы Wistar |
| |
| 3 | Проверка гипотезы о роли накопления Са2+ в развитии изменений цитоплазматических и экстрацеллюлярных белков в постуральных мышцах животных в условиях гравитационной разгрузки и возможность использования хелаторов кальция для предотвращения этих изменений. | 1. Антиортостатическое вывешивание крыс (14 суток) с внутрибрюшинным введением ЭГТА 30 мг/48ч. 3 группы:
| 21крыса Wistar |
| |
2. Антиортостатическое вывешивание крыс (14 суток) с энтеральным введением Нифедипина 7 мг/кг /24ч.
| 36 крыс Wistar | ||||
| 4 | Проверка гипотезы об эффектах пассивного растяжения, участии дистрофина и механизмах рекрутизации компонентов внутриклеточного цитоскелета и ЭЦМ постуральных мышц животных в условиях моделированной гравитационной разгрузки | 1. Антиортостатическое вывешивание (14 суток) на фоне пассивного растяжения мышц голени крыс
| 22 крысы Wistar |
| |
2. Локальное облучение мышц вывешенных крыс (14 суток) на фоне пассивного растяжения голени - облучение 2500 рад /15 мин, кобальт-60
| 43 крысы Wistar | ||||
РЕЗУЛЬТАТЫ ЭКСПЕРИМЕНТОВ И ИХ ОБСУЖДЕНИЕ
1. Эффекты реальной невесомости (условий космического полёта)
1.1 Исследование влияния реальной гравитационной разгрузки на изоформный состав и объём белков цитоплазматического цитоскелета и экстрацеллюлярного матрикса скелетных мышц человека
В ходе изучения биоптатов m. soleus четырёх членов экипажей МКС во всех исследованных случаях было обнаружено, что количество мышечных волокон с нарушениями дистрофинового слоя после космического полёта (~180 суток) было достоверно выше предполётных показателей. При этом было обнаружено высокое содержание таких волокон в предполетном периоде (21,7; 14,1; 22,7%). У космонавтов A и D количество волокон с разрывами дистрофинового слоя увеличилось в 3 раза, у космонавта B в 1,9 раза, у космонавта C в 2,1 раза. Однако, адекватный анализ данных изменений затруднён с одной стороны воздействием усиленных физических тренировок в предполётном периоде, и воздействием реадаптационной нагрузки при отложенном взятии биоптатов после завершения космического полёта.
Доля общего объёма коллагена достоверно увеличилась у всех членов экспедиции. При этом у космонавта А общий объем соединительной ткани увеличился на 112 %, у космонавта В на 69,8 %, у С космонавта до 106%. При этом достоверных изменений соотношения долей изоформ коллагена I и III типа у космонавтов А и В обнаружено не было. Интересно, что у космонавта С наблюдалось примерно равное распределение I и III изоформ коллагена до и после полёта. Таким образом, у всех испытателей наблюдалось достоверное увеличение общего объёма коллагена, без достоверных изменений изоформного состава.
1.2 Влияние реальной гравитационной разгрузки на объём и состав изоформ коллагена мышечных волокон m. soleus и m. tibialis anterior монгольской песчанки
При анализе мышечных волокон животных в течение 12 суток находящихся на борту биоспутника ФОТОН М3 было обнаружено достоверное увеличение доли общего коллагена в m. soleus на 21,4%, в m. tibialis anterior на 23,5%.
Изменения процентного соотношение изоформ коллагена I и III типа после 12 суточного полёта в m. soleus не обнаружено, прирост изоформы коллагена I типа составил 12%, изоформы коллагена III типа – 11%.
В m. tibialis anterior прирост изоформы коллагена I типа составил 8%, изоформы коллагена III типа – 12%.
Известно, что m. tibialis anterior, антагонист m. soleus, активируется в условиях невесомости [Alford E.K. et al., 1987], поэтому можно было ожидать, что атрофические изменения в нем будут слабее выражены. Однако, после 12 суточного космического полёта, увеличение общего объёма коллагена m. tibialis anterior монгольской песчанки и изменение состава изоформ коллагена I и III типа оказалось более выраженным. Известно, что экстрацеллюлярное окружение медленных волокон крыс плотнее быстрых, соответственно адаптивный ответ медленных и быстрых мышц может быть различным. Вероятно, более слабая реакция m. soleus песчанок на действие невесомости может быть связана с физиологическими особенностями (паракринной регуляцией) этого животного, которые до настоящего времени не достаточно изучены.
2. Динамика изменений компонентов внутриклеточного цитоскелета и экстрацеллюлярного матрикса камбаловидной мышцы крысы при моделируемой гравитационной разгрузке
В настоящем исследовании было проведено исследование динамики морфологических изменений различных компонентов внутриклеточного цитоскелета и внеклеточного матрикса в m. soleus крысы на разных отрезках времени в условиях моделируемой гравитационной разгрузки.
После 3-х суточного моделирования условий гравитационной разгрузки в m. soleus крысы наблюдалось 10-ти кратное увеличение количества десмин-негативных волокон. К 7-м суткам количество десмин-негативных волокон по сравнению с контролем было увеличено в 6 раз. Показатели 14-и и 30-и суточной разгрузки достоверно превышали контрольное значение и не отличались друг от друга.
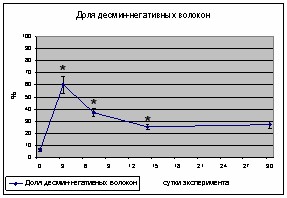

Рис.1 Динамика изменения количества десмин-негативных волокон после 3, 7, 14 и 30 суток гравитационной разгрузки камбаловидной мышцы крысы
Совокупность литературных данных указывает на быстрое восстановление уровня десмина на начальном этапе развития атрофических процессов в скелетной мышце. Тенденция к снижению уровня десмина в m. vastus крысы наблюдается уже в первые часы вывешивания [Enns et al., 2007]. Похожие изменения наблюдали в смешанной пробе m. soleus и m. gastrocnemius крысы после 3 суток антиортостатического вывешивания, при этом, к девятым суткам экспозиции эти изменения были полностью нивелированы [Chopard et al., 2001]. В то же время показано, что после 7 суток вывешивания в m.soleus мыши, уровень экспрессии индивидуальных мРНК десмина значительно снижается [Dpp et al., 2004]. Показано, что после антиортостатического вывешивания крысы определяется морфологическая дезорганизация филаментов десмина [Nara et al., 2002].
Опираясь на литературные данные, можно предположить, что деградация десмина на ранних сроках антигравитационной разгрузки, совпадающая с увеличением концентрации Са2+ в миоплазме, возможно является следствием активации кальпаинов.
При исследовании состояния дистрофина в m.soleus крысы в контрольной группе было обнаружено 10,7%±1,6% (p<0,05) волокон с разрывами дистрофинового слоя. В группе 3-х суточного вывешивания достоверных отличий от контроля не обнаружено. В группе 7-и суточного вывешивания количество повреждённых мышечных волокон увеличилось до 19%±1,5 (p<0,05), также увеличилось количество выпячиваний мембраны мышечных волокон, без видимых разрывов. После 14-суточного вывешивания наблюдалось увеличение количества разрывов дистрофинового слоя в местах выпячивания мембраны до 22%±2,7 (p<0,05). На поздних сроках эксперимента количество волокон с нарушением дистрофинового слоя саркоплазматической мембраны достигло 28%±3,5 (p<0,05), что не отличалось от показателя 14-и суточного вывешивания.
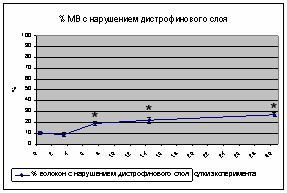

Рис. 2 Динамика изменения количества МВ с разрушением дистрофинового слоя.
Известно, что нарушение дистрофина ведёт к увеличению макромолекулярной проницаемости мембраны, повышению внутриклеточной концентрации ионов кальция и активации кальций-зависимого протеолиза [Гасникова Н.М. соавт., 2006]. Повреждение дистрофина нарушает транссарколеммальную связь актина с интегринами, ламинином-2 и коллагеном [Winder et al., 1997]. Показано, что дистрофин участвует в регуляции дигидропиридиновых каналов [Johnson et al., 2005], нейрональной NO-синтетазы и аквапоринового комплекса [Wakayama et al., 1997, Crosbie et al., 2002]. У животных, дефектных по гену дистрофина, наблюдается лавинообразное усиление протеолитических процессов в мышечной ткани [Acharyya et al., 2005]. Также показано достоверное увеличение доли волокон с повреждениями дистрофинового слоя после 14 суток вывешивания [Гасникова Н. М. соавт., 2006]. Однако, после 14 дней космического полета достоверных изменений в содержании дистрофина в m. soleus крысы не наблюдалось [Chopard et al., 2001, 2005], позднее авторы показали уменьшение относительного содержания дистрофина в m. soleus крысы на 20 и 30 % после 3 и 6 недель вывешивания соответственно.
- Можно предположить, что изменение макромолекулярной проницаемости мембраны вследствие потери дистрофина и накопление в миоплазме ионов кальция покоя вызывает изменения свойств механозависимых интегральных белков и структурные изменения МВ.
При исследовании изменений морфологических характеристик межволоконного коллагенового компонента m.soleus крысы показано, что на ранних сроках воздействия достоверных изменений объёма общего коллагена не наблюдалось. После 7-суточного вывешивания в m. soleus крысы наблюдалось увеличение в 2 раза содержания общего коллагена. После 14-суточного вывешивания наблюдалось дальнейшее увеличение содержания общего коллагена (в 4 раза больше в сравнении с контролем). После 30-суточного вывешивания в m. soleus крысы наблюдалось дальнейшее увеличение (в 7,3 раза в сравнении с контролем) содержания общего коллагена, вероятно, вследствие замещения редуцирующего сократительного компонента.


Рис. 3 Динамика изменения общего объёма соединительной ткани после 3, 7, 14 и 30 суток гравитационной разгрузки
Качественная морфометрия изоформ коллагена I и III типа на начальных сроках воздействия изменений их соотношения не выявила. Начиная с 7 суток эксперимента, доля изоформы коллагена I типа достоверно снижалась, на фоне достоверного увеличения доли изоформы коллагена III типа при значительном увеличении общего объёма соединительной ткани. Разрастание коллагеновых структур, в целом оказалось характерным для более поздних сроков моделируемой гравитационной разгрузки.
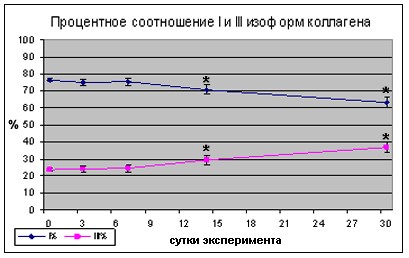

Рис.4 Динамика изменения изоформного состава коллагена после 3, 7, 14 и 30 суток гравитационной разгрузки
Ранее показано, что объем межволоконного компонента m. soleus человека достоверно увеличивается на 60-е сутки длительной антиортостатической гипокинезии, а также при иммобилизации m. Soleus у крысы [Шенкман и др., 2000; Roy et al., 2002]. Разрастание соединительной ткани, также наблюдали и у космонавтов в биоптатах m. soleus после 120-суточного полета на МКС [Belozerova et al., 2002]. Показано [Hirose et al., 2008], что при иммобилизации задних конечностей крыс экспрессия м-РНК проколлагена 1 типа снижается на 3 день и возвращается к контрольному уровню на 7 день. Также показано, что активность пролил 4-гидроксилазы и галактосилгидроксилизин глюкозилтрансферазы, являющихся белками биосинтеза коллагена, и концентрация гидроксипролина в камбаловидной мышце уменьшается после 3 и 42 дней корсетной иммобилизации. Известно, что экспрессия коллагена I типа фибробластами зависит как от непосредственно механо-зависимых путей сигнализации, первичными датчиками в которых являются интегрины, так и от воздействия ростовых факторов, некоторые из которых связаны со структурами внеклеточного матрикса и освобождаются при действии механического напряжения (b-FGF) [Kjr 2004]. Активность м-РНК коллагена 1 типа наблюдается преимущественно в нервных тканях и мышечном веретене [Hirose et al., 2008]. В то же время известно, что концентрация ростовых факторов в мышечной ткани при гравитационной разгрузке значительно снижена [Clarke et al., 1993, 1998, 2000].
Увеличение объема коллагена в условиях вывешивания может зависеть как от снижения проприоцептивной активности мышцы, так и от уровня экспрессии различных факторов роста. Изменение экспрессии изоформ коллагена может быть вызвано как инактивацией интегриновых путей, так и изменением уровня экспрессии b-FGF и аналогичных ростовых факторов.
3. Роль накопления ионов кальция в миоплазме
Антиортостатическое вывешивание крыс (14 суток) на фоне применения блокаторов медленных кальциевых каналов L-типа
В настоящем эксперименте показано, что ограничение проницаемости клеточной мембраны для ионов Са2+ при помощи блокаторов кальциевых каналов L-типа в мышечных волокнах m.soleus крыс привело к частичной компенсации эффектов разгрузки, в частности, предотвращению нарушений целостности дистрофинового слоя сарколеммы и сохранению объёма общего коллагена.
В ходе эксперимента, показано, что количество десмин-негативных волокон, и в группе чистого антиортостатического вывешивания, и в группе антиортостатического вывешивания с введением нифедипином, по сравнению с контролем, увеличился в 4 раза и составил 21,5±1,1% и 25,2±1,9% (p<0,05), без достоверных отличий между вывешенными группами.
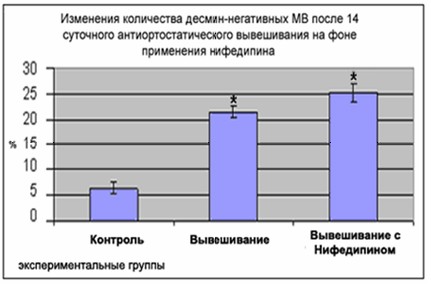
Рис. 5 Изменения количества десмин-негативных МВ m.soleus крыс после 14 суточного антиортостатического вывешивания на фоне применения нифедипина
Таким образом, блокада медленных кальциевых каналов L-типа не оказывает влияния на реализацию адаптационных изменений сети ПФ десмина, что, несмотря на хронологическую корреляцию увеличения количества десмин-негативных волокон с деструкцией саркомерного цитоскелета, свидетельствует об отсутствии зависимости десмина от концентрации Са2+ в миоплазме. Возможно, что в случае с десмином, лишь дальнейшие исследования прольют свет на характер зависимости содержания этих белков от концентрации кальция в миоплазме.
При исследовании состояния дистрофинового слоя оказалось, что процент повреждённых мышечных волокон в группе крыс, получавших нифедипин на фоне вывешивания, оказался достоверно ниже (17,4±1,1 (p<0,05)) этого показателя в группе вывешивания (22% ± 2,7 (p<0,05)), однако превышал контрольное значение (10,7±1,6 (p< 0,05)).
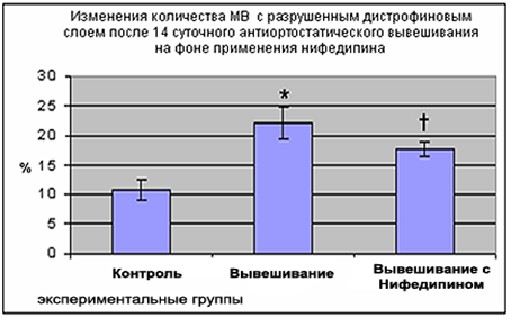
Рис. 6 Изменения количества МВ m.soleus крыс с разрушенным дистрофиновым слоем после 14 суточного антиортостатического вывешивания на фоне применения нифедипина
Менее выраженный уровень повреждения дистрофинового слоя на фоне хронического введения нифедипина может быть обусловлен соседством кальциевых каналов L-типа и дистрофин-саркогликанового комплекса.
При исследовании морфологических характеристик коллагена m. soleus крысы показано, в группе 14-суточного антиортостатического вывешивания увеличение общего объёма коллагена составило 37,5±6,5% (p<0,05). В группе вывешивания с нифедипином увеличение общего объёма коллагена составило 13,3±5,5%, что достоверно отличалось от контрольных показателей.
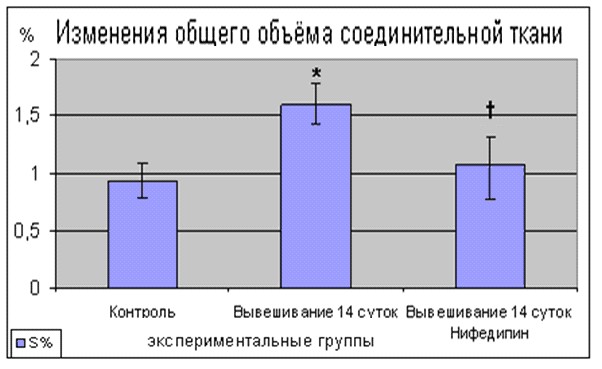

Рис. 7 Изменения содержания общего объёма коллагена МВ m.soleus крыс после 14 суточного антиортостатического вывешивания на фоне применения нифедипина
При 14-суточном антиортостатическом вывешивании крыс в m. soleus наблюдалось изменение соотношения изоформ коллагена с относительным увеличением доли изоформы коллагена III типа (15,4±2,8% (p<0,05)). В группе вывешивания с нифедипином изменение соотношения изоформ коллагена I и III типа составило 12,2±2,5%, таким образом, наблюдалась тенденция к предотвращению увеличения коллагена III типа.
Компенсация фиброза при вывешивании на фоне введения нифедипина, обнаруженный в эксперименте, может быть связан с кальций-зависимым снижением аутокринной экспрессии Ил-6, показанный при вывешивании [Grano et al., 2002], и способного (через цАМФ и IP3) с одной стороны активировать фибробласты и увеличивать экспрессию коллагена, с другой повышать каталитическую активность матриксных металлопротеиназ (ММР). Интересно, что in vivo, блокада рианодиновых кальциевых каналов не влияла уровень экспрессии Ил-6 [Juretic et al., 2005]. Изменения соотношения изоформ коллагена также может регулироваться изменением экспрессии системы MMP/TIMP.
Таким образом, показано, что в ходе развития атрофических и компенсаторных изменений различных компонентов цитоскелета фармакологическая блокада поступления ионов Са2+ покоя частично предотвращает деструктивные эффекты гравитационной разгрузки.
4. Роль внутримышечного механического напряжения.
Антиортостатическое вывешивание (14 суток) на фоне пассивного растяжения мышц голени крыс
В настоящем эксперименте показано, что пассивное растяжение m.soleus крыс на фоне вывешивания компенсирует увеличение общего объёма соединительной ткани в m.soleus.
При исследовании морфологических характеристик коллагена в m.soleus крысы оказалось, что в группе вывешивания с растяжением доля ППС среза, занятого общим коллагеном оказалась достоверно ниже этого показателя в группе вывешивания (5,3%), и составила 3,6% (р<0,05). Композиция изоформ коллагена I и III типа составила 62,4% и 37,6% соответственно, что было достоверно ниже этих показателей в группе 14-суточного вывешивания (58,4% и 42,6% (р<0,05) соотв.), но отличалось от контрольной группы (69,2%, 30,8% (р<0,05)).
Таким образом, экспериментально установлено, растяжение на фоне вывешивания сопровождается несколько меньшим приростом общего коллаген, и, на этом фоне, слабо выраженным изменением композиции изоформ коллагена I и III типа.
Феномен снижения фиброза при пассивном растяжении на фоне вывешивания крысы может быть вызван активацией механо-зависимых путей сигнализации или изменениями уровней локальной экспрессии факторов роста. Показано, что после 7 и 14 суток вывешивания уровни экспрессии м-РНК коллагена I и III, TGF-1 и CTGF во время вывешивания крыс не менялись, а восстановление вызывало увеличение экспрессии м-РНК TGF-1 и мышечного коллагена I и III (более 10 раз) [Hirose et al., 2008]. Было показано, что стимул резистивного упражнения увеличивает объём экспрессии тотального РНК (трансляционная ёмкость), а также уровни экспрессии м-РНК IGF-1, миогенина и протоколлагена III--1, и снижает экспрессию м-РНК миостатина [Wehling et al., 2000]. Известно, что миостатин стимулирует пролиферацию мышечных фибробластов и через активацию Smad, p38, MAPK и Akt запускает продукцию белков экстрацеллюлярного матрикса и in vivo, и in vitro [Zhao et al., 2008]. Показано, что наибольшее иммуногистохимическое выявление TGF-1 и TNF- наблюдается в соединительной и нервной ткани, особенно в области нервных окончания и мышечных веретён, экспрессия м-РНК 1 типа коллагена также локализована преимущественно в нервных тканях и мышечном веретене [Weigert et al., 2007].
Результаты настоящего исследования согласуются с литературными данными и позволяют предположить, что факторы, обеспечивающие антиатрофический эффект, также препятствуют разрастанию соединительной ткани [Таракина соавт., 2008]
Исследование роли пролиферативных процессов в изменениях морфологических характеристик компонентов экстрацеллюлярного цитоскелета m.soleus крыс при гравитационной разгрузке.
При исследовании морфологических характеристик коллагена в m.soleus крысы после 14-суточного антиортостатического вывешивания с использованием локального -облучения голени (2500рад/ 15 минут; изотоп кобальта–60). Воздействие локального облучения на фоне вывешивания способствовало поддержанию общего объёма соединительной ткани (6,9%) и композиции изоформ коллагена (66,8% и 33,1% соответственно) на уровне контрольной группы (5,1%; 64,7% и 32,2%).
Таким образом, в эксперименте было показано, что 14-суточное вывешивание приводит к увеличению объёма соединительной ткани почти вдвое, причём в основном за счёт относительного увеличения III коллагена, характеризующегося большей податливостью к напряжению; а также к снижению коллагена I, обеспечивающего продольную жёсткость и препятствующего механическому повреждению волокна при нагрузке. Локальное облучение m.soleus на фоне растяжения практически полностью предотвращало увеличение соединительно-тканного компонента в условиях антиортостатического вывешивания. Полученные данные могут быть интерпретированы в пользу гипотезы о непосредственном участии пролиферирующих в условиях гравитационной разгрузки синовиальных фибробластов в перестройке соотношения изоформ коллагена.
ВЫВОДЫ:
- Нарастание общего объёма коллагена в камбаловидной мышце крысы при антиортостатическом вывешивании прямо пропорционально его продолжительности и сопровождается увеличением доли изоформы коллагена III типа и уменьшением доли изоформы коллагена I типа.
- Содержания десмина в камбаловидной мышце крысы резко снижается на ранних сроках моделированной гравитационной разгрузки и частично восстанавливается в течение короткого периода времени.
- Состояние дистрофинового слоя волокон камбаловидной мышцы крысы на ранних сроках антиортостатического вывешивания практически не меняется. К 7-м суткам эксперимента количество волокон с нарушенным дистрофиновым слоем удваивается. На более поздних сроках воздействия наблюдается тенденция к постепенному увеличению таких волокон.
- Применение блокаторов медленных кальциевых каналов L-типа предотвращает увеличение общего объёма коллагена, способствует сохранению дистрофинового слоя, но не влияет на изменение изоформного состава коллагена и количество десмин-негативных волокон в камбаловидной мышце крысы при 14-суточном антиортостатическом вывешивании.
- Пассивное растяжение камбаловидной мышцы в условиях 14-суточного антиортостатического вывешивания крыс сопровождается меньшим приростом общего объёма коллагена и менее выраженным изменением его изоформного состава, при этом не оказывает влияния на количество десмин-негативных волокон.
- Локальное облучение камбаловидной мышцы крысы в сочетании с пассивным растяжением на фоне 14-суточного антиортостатического вывешивания в значительной мере компенсирует увеличение объёма общего коллагена и перераспределение изоформ коллагена в сторону изоформы III типа, что свидетельствует о роли пролиферации фибробластов в развитии этих изменений.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
- «Физиологические реакции обезьян в условиях 9-суточного иммерсионного воздействия и длительной антиортостатической гипокинезии»; Новости медико-биологических наук, №4, 2004, С. 46-51
- «Physiological reactions of primates to 9-D immersion and head-down immobilization» J. Gravit. Physiol. 2004, V.11, #2 : P. 29-P30
- «Случай дистальной мышечной дистрофии с поздним дебютом (тип Веландер)» Неврологический журнал, 2006, №6, С. 25-29
- «Креатин как метаболический модулятор структуры и функции скелетных мышц при силовой тренировке у человека. Клеточные механизмы» Российский физиологический журнал им. И.М. Сеченова, 2006, 92 N-1, P.100
- «Синдром ригидного человека с глазодвигательными и мозжечковыми нарушениями» Анналы неврологии, 2007, т.1, №4, С.15-22
- «Патофизиологические аспекты дисфункции тазового дна» Российский медицинский форум, №2, 2008, С. 16-23
Публикации в трудах научных мероприятий
- «Роль опоры в развитии изменений постуральных (m.soleus) и локомоторных (m. vastus lat.) мышц нижних конечностей обезьян в эксперименте 7-суточной сухой иммерсии» Конференция молодых учёных ГНЦ РФ ИМБП РАН, Москва 2004, С. 35
- «Изменения коллагенового матрикса и десминового цитоскелета m.soleus крысы при разных сроках гравитационной разгрузки» Конференция молодых учёных ГНЦ РФ ИМБП РАН, Москва 2005
- «Time course changes of collagen matrix and desmin cytoskeleton alteration in rat soleus at 3, 7, 14 и 30 days of gravitational unloading» XXXIV European Muscle Conference, 2005 P.17
- «Изменения коллагенового матрикса и десминового цитоскелета m.soleus крысы при 3, 7, 14 и 30 сутках гравитационной разгрузки» XIII международное совещание и VI школа по эволюционной физиологии, Институт эволюционной физиологии и биохимии им. И.М.Сеченова, 23-28.01.2006, ВММ, 2006 С.235
- «Resting Intracellular Calcium Level as a Triggering Signal for Soleus Fiber Alterations during Unloading» 27th Annual International Gravitational Physiology Meeting, Osaka, Japan, 2006, С.
- «Time course changes of Desmin immunoreactivity in unloaded rat soleus» Journal of Muscle Research and Cell Motility, 2006
- «Effects of gravitational unloading in soleus fibers of dystrophin-deficient mice» Journal of Muscle Research and Cell Motility, 2006, Vol. 27 № 5-7, P. 510
- «Time course changes of cytoskeletal proteins and collagen isoforms in rat soleus during hindlimb suspension» 16th IAA Human in Space Symposium, 21-24.05.2007, P.207
- «Состояние экстрацеллюлярного матрикса m. soleus крысы при хроническом растяжении на фоне антиортостатического вывешивания» VI Конференция молодых учёных ГНЦ РФ ИМБП РАН, 2007, Москва C. 62
- «Влияние космического полёта на состояние постуральных мышц монгольской песчанки» VII Конференция молодых учёных ГНЦ РФ ИМБП РАН, 2007, Москва, C.74
- «Синдром ригидного человека с глазодвигательными и мозжечковыми нарушениями: сдвиг в сторону синтеза медленных форм миозина, деструкция цитоскелетных белков пораженных мышц» VII Конференция молодых учёных ГНЦ РФ ИМБП РАН, 2007, Москва, C.28
- «Влияние нифедипина на состояние дистрофинового слоя m.soleus крысы при гравитационной разгрузке» VI Симпозиум Химия протеолитических ферментов, 2007, Москва, С. 172