

Психоэмоциональный ответ на стресс и экспрессия генов нейропластичности в мозге
На правах рукописи
Берёзова Инна Валерьевна
Психоэмоциональный ответ на стресс
и экспрессия генов нейропластичности в мозге
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
физиология – 03.03.01
Новосибирск – 2012
Работа выполнена в лаборатории функциональной нейрогеномики Учреждения российской академии наук Института цитологии и генетики СО РАН, г. Новосибирск.
| Научный руководитель: | доктор биологических наук Шишкина Г.Т. Институт цитологии и генетики СО РАН, г. Новосибирск |
| Официальные оппоненты: | доктор биологических наук Вольф Н.В. ГУ НИИ физиологии СО РАМН, г. Новосибирск кандидат биологических наук Оськина И.Н. Институт цитологии и генетики СО РАН, г. Новосибирск |
| Ведущее учреждение: | Государственное учреждение Научный центр клинической и экспериментальной медицины СО РАМН, г. Новосибирск |
Защита диссертации состоится «___» ____________ 20___ г. на __________ заседании диссертационного совета Д. 001.14.01 по защите диссертаций на соискание ученой степени доктора наук в ФГБУ НИИ физиологии СО РАМН в конференц-зале Института по адресу:
630117, г. Новосибирск, ул. Академика Тимакова, 4, тел.: (383) 334-89-61, факс: (383) 335-97-54, е-mail: [email protected].
С диссертацией можно ознакомиться в библиотеке ФГБУ НИИ физиологии СО РАМН.
Автореферат разослан «___» _____________ 20___ г.
Ученый секретарь
диссертационного совета,
кандидат биологических наук И.И. Бузуева
Актуальность проблемы. Депрессия – это серьезное психоэмоциональное расстройство, от которого страдает до 21% населения в развитых странах (Gainotti et al., 2001). Это заболевание характеризуется потерей интереса к обычным радостям жизни (ангедония) и в своем крайнем проявлении может сопровождаться попытками суицидального исхода (Nestler et al., 2002). Несмотря на распространенность депрессии и связанные с ней социально-экономические последствия, причины, индуцирующие развитие этого психоэмоционального нарушения, а также пути его формирования остаются неясными.
Среди различных факторов, предрасполагающих к возникновению депрессии, значительная роль отводится в настоящее время стрессорным жизненным обстоятельствам (Kendler et al., 1999; Frodl et al., 2010). Исследование механизмов индуцированной стрессом психопатологии на животных моделях показало, что стресс, провоцирующий у грызунов развитие депрессивноподобного состояния, может сопровождаться различными нейропластическими изменениями, включающими угнетение нейрогенеза в гиппокампе (Fuchs et al., 2004) и усиление гибели нейрональных клеток в коре (Bachis et al., 2008). Было высказано предположение, что эти эффекты стресса на мозг являются одной из причин наблюдаемого у некоторых депрессивных больных уменьшения объемов ряда отделов мозга, таких как гиппокамп, кора и миндалина (Bremner et al., 1995; Bremner et al., 1997; Bremner et al., 2000).
Угнетение нейрогенеза и усиление апоптоза после стресса привлекли внимание к факторам, регулирующим эти процессы. Наибольшее внимание было привлечено вначале к нейротрофическим факторам, прежде всего мозговому нейротрофическому фактору (brain-derived neurotrophic factor, BDNF). Отмечаемое, преимущественно в гиппокампе, снижение экспрессии этого фактора после стрессорных воздействий и восстановление этого снижения длительным введением антидепрессантов привели к созданию нейротрофической гипотезы депрессии (Duman, Monteggia, 2006), согласно которой изменение уровня BDNF является ключевым механизмом как формирования, так и терапии этой психопатологии.
Помимо мозгового нейротрофического фактора, в недавних работах было обнаружено, что хронический стресс может приводить также к изменениям в экспрессии белков, регулирующих процессы апоптоза (Kosten et al., 2008). Однако взаимосвязь этих вызываемых стрессом изменений в экспрессии регуляторов апоптоза с появлением симптомов депрессивных расстройств остается совершенно неясной.
Наряду с выяснением механизмов индуцируемых стрессом психоэмоциональных нарушений, таких как депрессия, чрезвычайно важным является также понимание путей, обеспечивающих устойчивость к возникновению этой патологии. Ведь, несмотря на то, что каждый из нас ежедневно сталкивается со стрессирующими ситуациями, далеко не у всех развивается депрессия, и большинство людей, к счастью, являются нечувствительными к таким негативным эффектам стресса. Выявление эндогенных факторов устойчивости является в настоящее время чрезвычайно быстро развивающимся направлением исследований в проблеме формирования и терапии депрессивных расстройств, и, в дальнейшем, возможно, на основе этих факторов будет создано принципиально новое эффективное средство для борьбы с депрессией.
Целью данной работы явилось, таким образом, исследование на взрослых самцах крыс эффектов стрессорного воздействия, индуцирующего у животных развитие депрессивноподобного состояния, а также введения антидепрессанта флуоксетина на экспрессию регуляторов нейропластичности. В качестве стрессорного воздействия было использовано вынужденное плавания. Неизбегаемые условия этой процедуры являются для грызунов сильным стрессором, значительно активирующим гипоталамо-гипофизарно-адренокортикальную систему и приводящим к развитию характерного синдрома – поведенческой беспомощности, который считается аналогом соответствующего клинического симптома у депрессивных больных. Продолжительность пассивного поведения или замирания (immobility), увеличивающаяся в процессе повторных воздействий вынужденным плаванием, снижается в ответ на введение антидепрессантов (Porsolt et al., 1977; Porsolt et al., 1978), что явилось причиной широкого использования этой простой процедуры для преклинической оценки действия антидепрессантов. В последнее время вынужденное плавание также активно применяется для моделирования депрессивноподобного состояния грызунов с целью исследования механизмов его формирования (Cryan et al., 2005).
В работе были поставлены следующие конкретные задачи:
- Исследовать влияние однократного и повторного стресса вынужденного плавания на экспрессию генов BDNF, а также ключевых регуляторов апоптоза, антиапоптозного белка Bcl-xl и проапоптозного белка Вax, в структурах мозга через 24 часа после воздействия. Данная временная точка выбрана в соответствии с обычно используемой для оценки степени депрессивноподобного состояния.
- Исследовать острые эффекты (через 40 минут и 2 часа) повторного стресса вынужденного плавания на экспрессию регуляторов нейропластичности и сопоставить возможные изменения в экспрессии их генов с выраженностью психоэмоционального ответа на стресс (содержание адренокортикотропного гормона и кортикостерона в плазме крови, продолжительность пассивного поведения).
- Проанализировать возможность модуляции базальной и стрессорной экспрессии генов регуляторов нейропластичности введением антидепрессанта флуоксетина, широко используемого в клинической практике для терапии депрессии.
Научная новизна. В работе получены новые и оригинальные данные, свидетельствующие о значительном влиянии стресса, индуцирующего развитие депрессивноподобного состояния, на экспрессию генов мозгового нейротрофического фактора и белков апоптоза. Выявленные изменения в экспрессии исследованных генов зависели от отдела мозга, продолжительности стрессорного воздействия и времени после его окончания.
В работе было обнаружено, что через сутки после однократного стрессорного воздействия экспрессия мозгового нейротрофического фактора во фронтальной коре была снижена, в то время как через сутки после повторного воздействия экспрессия фактора в этом отделе мозга не отличалась от базального значения. В отличие от фронтальной коры, экспрессия этого нейротрофина в гиппокампе не была изменена через сутки после однократного воздействия, в то время как через сутки после повторного стресса была достоверно повышенной. В среднем мозге через сутки ни после однократного, ни повторного стрессорного воздействия изменений в экспрессии фактора обнаружено не было. Вместе с тем, более подробное исследование экспрессии нейротрофина, проведенное через 40 минут и 2 часа после окончания повторного вынужденного плавания, выявило значительное его снижение в гиппокампе и среднем мозге через 2 часа. Это снижение, очевидно, обусловлено стрессорной активацией секреции кортикостерона, на что указывает достоверная отрицательная корреляция между мРНК фактора в гиппокампе и уровнем гормона в плазме крови.
Согласно нейротрофической гипотезе депрессии, вызванное стрессом снижение экспрессии мозгового нейротрофического фактора в отделах мозга может вовлекаться в формирование депрессивноподобного состояния. Свидетельством такой возможности, по крайней мере, во фронтальной коре, служит достоверная отрицательная корреляция между уровнем мРНК BDNF в этом отделе мозга и продолжительностью пассивного поведения животных. В отличие от снижения через 2 часа после повторного воздействия экспрессии BDNF, его последующее через 24 часа после стресса увеличение в гиппокампе, вероятно, является проявлением механизмов, обеспечивающих защиту или преодоление организмом негативных эффектов стрессорного воздействия на нейрональные клетки. С выполнением этой же функции, очевидно, связано и увеличение экспрессии гена антиапоптозного белка Bcl-xl в этом отделе через 24 часа после однократного стрессорного воздействия.
Наиболее важным и интересным результатом проведенной работы является усиление экспрессии гена и белка Bcl-xl в гиппокампе через 2 часа после повторного стресса вынужденного плавания. Сопоставление этого повышения с выраженностью индуцированного стрессом депрессивноподобного состояния у отдельных животных позволило прийти к заключению, что способность к такому повышению может быть важным фактором, определяющим устойчивость к формированию индуцируемых стрессом поведенческих признаков депрессии. Отсутствие же способности организма отвечать на стрессорное воздействие усилением экспрессии гена Bcl-xl может приводить к развитию негативного психоэмоционального состояния. Подтверждением такого предположения служит достоверная отрицательная корреляция между отношением Bcl-xl/Bax в гиппокампе и продолжительностью поведенческой депрессии. На участие антиапоптозного белка Bcl-xl в развитии индуцируемого стрессом депрессивноподобного состояния указывает также усиление экспрессии гена этого белка после стресса длительным введением антидепрессанта флуоксетина.
Научно-практическая ценность. Результаты работы расширяют теоретические представления о факторах, вовлекаемых в развитие депрессивноподобного поведения, индуцированного стрессорными воздействиями, а также в механизмы устойчивости к формированию этого негативного эмоционального состояния. Кроме того, полученные данные могут иметь и определенное практическое значение, поскольку блокатор обратного захвата серотонина флуоксетин в настоящее время применяется в мировой лечебной практике для терапии депрессии.
Апробация работы. Результаты данной работы были представлены и обсуждены: на отчётной сессии ИЦиГ СО РАН в 2010 г., на XLVI, XLVII и XLVIII международных научных студенческих конференциях «Студент и научно-технический прогресс» (г. Новосибирск, 2008, 2009, 2010), VII Всероссийской конференции с международным участием «Механизмы функционирования висцеральных систем», (г. Санкт-Петербург, 2009), XXI Съезде физиологического общества имени И.П. Павлова (г. Калуга, 2010), V съезде Вавиловского общества генетиков и селекционеров (г. Москва, 2009), Пятой Всероссийской научно-практической конференции с международным участием (г. Новосибирск, 2011), а также международных конференциях «7th World Congress on Stress» (Нидерланды, 2010), «10th Symposium on Catecholamines and Other Neurotransmitters in Stress» (Словакия, 2011), «8th IBRO World Congress of Neuroscience» (Италия, 2011) и «24th ECNP Congress» (Франция, 2011).
Публикации. Материалы диссертации представлены в 16 работах, в том числе 4 статьи в отечественных (2) и зарубежных (2) реферируемых журналах.
Структура и объём работы. Работа включает следующие разделы: введение, обзор литературы, материалы и методы, результаты, обсуждение, заключение, выводы, список цитированной литературы (213 источников). Общий объем составляет 97 листов. Представлено 19 рисунков и 1 таблица.
МАТЕРИАЛЫ И МЕТОДЫ
В работе использовали 75 взрослых самцов крыс линии Вистар массой 220-240 г. Животных содержали в течение эксперимента по одному в клетке при стандартных условиях вивария ИЦиГ СО РАН.
На этих животных в первой части работы исследовали влияние однократного и повторного стресса вынужденного плавания на экспрессию в отделах мозга регуляторов нейропластичности (BDNF, Bcl-xl и Bax) в сопоставлении с выраженностью индуцируемого стрессорным воздействием депрессивноподобного состояния и содержанием гормонов стресса в плазме крови. Продолжительность и порядок стрессорных процедур были аналогичны используемым в тесте Порсолта для оценки антидепрессантного действия препаратов. Согласно классическому варианту этого теста (Porsolt et al., 1978), крыс подвергали с промежутком в 24 часа двум принудительным плаваниям в течение 15 и 5 минут в неизбегаемых условиях стеклянного цилиндра (46*20 см), наполненного водой (24-26°С) до высоты 30 см. Анализ эффектов стресса проводили через 24 часа после однократного, а также через 40 минут, 2 и 24 часа после повторного воздействий. В каждой из этих групп, а также группе контрольных нестрессированных животных содержалось по 10 животных.
Во второй части работы исследовали эффекты на базальный и стрессорный уровни экспрессии генов BDNF, Bcl-xl и Bax антидепрессанта флуоксетина, который вводили с питьевой водой в дозе 7,5 мг/кг/день в течение 2-х недель. Контрольные животные (10 животных) получали воду без антидепрессанта. После последнего приема половина самцов крыс из каждой группы (по 5 животных) были подвергнуты двум описанным выше процедурам вынужденного плавания. Через сутки после повторного плавания эти крысы вместе с оставшимися нестрессированными животными были декапитированы.
После декапитации у животных быстро выделяли на холоде гиппокамп, фронтальную кору и средний мозг. Из образцов ткани мозга одностадийным гуанидин-изотиоционатным методом выделяли суммарную РНК (Chomczynski, Sacchi, 1987). Для получения кДНК использовали Оligo-dT-праймеры и MuLV ревертазу (СибЭнзим, Россия). Уровни мРНК анализируемых генов в первой части работы определяли методом полуколичественной ОТ-ПЦР. Амплификацию специфичных для каждого гена участков кДНК проводили по стандартной методике с использованием прямых и обратных праймеров для BDNF (Bittigau, Sifringer et al. 2002), Bax (Tamatani et al., 1999), Bcl-xl (Shindler et al., 1997), или -актина (Nudel et al., 1983). Уровни мРНК целевых генов оценивали относительно уровня мРНК -актина после сканирования в УФ продуктов ПЦР, разделенных электрофорезом в 1.5% агарозном геле, окрашенном бромистым этидием, с последующей денситометрией. Во второй части работы уровень экспрессии мРНК определяли методом ПЦР в реальном времени (4-5 образцов в каждой исследованной группе) с использованием наборов TaqMan® Gene Expression Assays на амплификаторе AB Prism 7000 (“Applied Biosystems”, США).
В собранной после декапитации плазме крови иммуноферментным методом определяли уровни гормонов с использованием стандартных наборов для кортикостерона (Assay Designs, США) и адренокортикотропного гормона (Biomerica, Германия).
Белок Bcl-xl в гиппокампе выявляли иммуногистохимически (пять животных в каждой группе). Крыс транскардиально перфузировали последовательно фосфатным буфером и 4%-ным параформальдегидом, головной мозг извлекали, насыщали 30%-ной сахарозой и замораживали в изопентане. Срезы маркировали 1 день при 4 °С специфичными антителами к белку Bcl-xl (1:100; Santa Cruz Biotechnology, США). Затем срезы обрабатывали биотинилированными вторичными антителами козы против иммуноглобулина кролика (1:100; Santa Cruz Biotechnology, США) 16 часов при температуре 4 °С, 2 часа при комнатной температуре инкубировали со стрептавидином, конъюгированным с пероксидазой хрена (1:500; “Abcam”, Англия). Пероксидазную активность выявляли 3,3'-диаминобензидином.
Данные обрабатывали статистически с применением t-критерия Стьюдента, однофакторного и двухфакторного дисперсионного анализов (ANOVA) с последующими парными post-hoc сравнениями отдельных групп (Fisher’s LSD), а также корреляционного анализа (по Пирсону).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Влияние однократного и повторного стресса вынужденного плавания на экспрессию генов BDNF, Bcl-xl и Bax в отделах мозга через 24 часа после воздействия
 | Рис. 1. Уровень мРНК BDNF в отделах мозга через 24 часа после однократного и повторного стресса вынужденного плавания. * - p < 0.05 по сравнению с контролем; # - p < 0.05 по сравнению с уровнем через 24 часа после однократного стресса. |
BDNF. Стресс, вызванный неизбегаемыми условиями вынужденного плавания, оказал значительное влияние на экспрессию гена BDNF в гиппокампе (F2,20 = 4.053, p < 0.05) (рис. 1). Однако, как показали последующие парные сравнения, повышающий в целом эффект вынужденного плавания был достоверным лишь после повторного воздействия (p < 0.05).
Однофакторный ANOVA не выявил влияния вынужденного плавания на уровень мРНК BDNF во фронтальной коре (F2,20 = 2.439, p = 0,113). Однако прямые парные сравнения отдельных групп с использованием t-критерия Стьюдента показали достоверное уменьшение экспрессии гена BDNF через сутки после однократного плавания и отсутствие каких-либо отличий от контрольных животных через сутки после повторного воздействия.
Уровень мРНК BDNF в среднем мозге не отличался от контроля ни после однократного, ни повторного стрессорных воздействий.
Bcl-xl. Анализ экспрессии гена Bcl-xl (рис. 2А) выявил достоверное влияние стресса вынужденного плавания на экспрессию этого гена в гиппокампе (F2,22 = 5.483, p < 0.05). Этот эффект обусловлен значительным (p < 0.05) увеличением уровня мРНК Bcl-xl в этом отделе мозга через 24 часа после однократного плавания. В то же время через 24 часа после повторного плавания уровень мРНК Bcl-xl не отличался от контрольного значения. Во фронтальной коре и среднем мозге изменений в экспрессии гена антиапоптозного белка после стрессорных воздействий обнаружено не было.
 |  |
Рис. 2. Уровень мРНК Bcl-xl (А) и Вах (Б) в отделах мозга через 24 часа после однократного и повторного стресса вынужденного плавания. * - p < 0.05 по сравнению с контролем; # p < 0.05 по сравнению с уровнем через 24 часа после однократного стресса.
Вах. В отношение этого проапоптозного белка, небольшое, но достоверное (p < 0.05) снижение его генной экспрессии обнаружено во фронтальной коре через сутки после повторного плавания (F2,21 = 3.891, p < 0.05, рис. 2Б). Небольшие изменения экспрессии гена Bax в гиппокампе и среднем мозге не достигали статистической значимости.
Исследование генной экспрессии BDNF, Bcl-xl и Bax в отделах мозга в связи с вызываемым острым стрессом повторного вынужденного плавания изменением уровней адренокортикотропного гормона и кортикостерона в плазме крови
АКТГ и кортикостерон. Стресс вынужденного плавания является для грызунов сильным стрессором, активирующим их гипоталамо-гипофизарно-адренокортикальную систему и, как представлено на рисунке 3, значительно повышающим содержание в крови как АКТГ (F3,36 = 28.37, p < 0.001), так и кортикостерона (F3,36 = 28.37, p < 0.001).
 | Рис. 3. Уровень АКТГ и кортикостерона в плазме крови через 40 мин, 2 и 24 часа после повторного теста вынужденного плавания. * - p < 0.05 по сравнению с уровнем гормона у контрольных животных. |
BDNF. Вызванное стрессом вынужденного плавания повышение содержания гормонов (АКТГ и кортикостерона) в плазме крови сопровождалось значительным, достигающим достоверности (p < 0.05) через 2 часа после воздействия, снижением уровней мРНК BDNF (рис. 4А) в гиппокампе (F3,24 = 11.199, p < 0.001) и среднем мозге (F3,29 = 3.716). Экспрессия гена нейротрофина через сутки после воздействия намного превышала контрольное значение в гиппокампе (p < 0.05), в то время как в среднем мозге эта экспрессия от контроля не отличалась. Небольшое снижение уровня мРНК BDNF через 2 часа после стресса не было достоверным.
Bcl-xl. В отличие от BDNF, через 2 часа после стрессорного воздействия – вынужденного плавания, экспрессия гена Bcl-xl в гиппокампе достоверно увеличилась (F3,27 = 4.426, p < 0.05), в то время как во фронтальной коре и среднем мозге осталась без изменений (рис. 4Б). Через сутки после стресса уровень мРНК Bcl-xl в гиппокампе не отличался от контроля.
Bax. Уровень экспрессии гена проапоптозного белка Bax не был изменен повторным стрессом вынужденного плавания ни в одном из исследованных отделов мозга (гиппокамп - F3,32 = 0.971, p > 0.05, фронтальная кора - F3,29 = 1.258, p > 0.05, средний мозг - F3,29 = 0.98, p > 0.05) (данные не показаны).
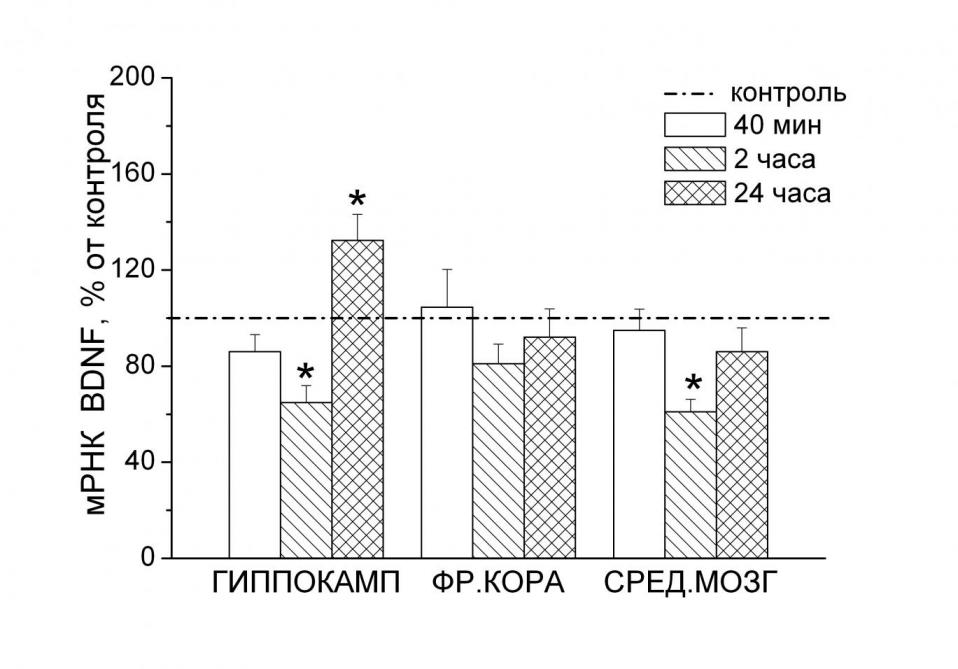 |  |
Рис. 4. Уровень мРНК BDNF (А) и Bcl-xl (Б) в отделах мозга через 40 минут, 2 и 24 часа после повторной процедуры вынужденного плавания. - p < 0.05 по сравнению с контролем.
Сравнение динамик уровней гормонов в плазме крови и мРНК исследованных генов в отделах мозга выявило между экспрессией BDNF в гиппокампе и уровнем кортикостерона в крови наличие достоверной отрицательной корреляции (r = -0.40, p<0.05, n = 28). Повышение уровня кортикостерона сопровождалось угнетением экспрессии фактора в этом отделе мозга, в то время как нормализация уровня гормона – увеличением его экспрессии. Эти результаты свидетельствуют в пользу того, что изменения экспрессии нейротрофина, которые были выявлены в нашей работе, могли быть обусловлены повышением в результате перенесенного стресса уровня глюкокортикоидов. Однако механизм действия глюкокортикоидов в этой связи остается неясным. Классический глюкокортикоид-чувствительный сайт в промоторах гена этого нейротрофина до сих пор не выявлен (Huynh, Heinrich, 2001), хотя угнетающее влияние высоких доз гормонов коры надпочечников на уровень мРНК BDNF в мозге продемонстрировано во многих работах (Barbany, Persson, 1992; Smith et al., 1995; Hansson et al., 2006). Изменение уровня экспрессии BDNF после стресса может быть связано также с модуляцией стрессорными воздействиями, включающими и вынужденное плавание, уровня экспрессии транскрипционного фактора CREB (cAMP response element binding protein) (Bilang-Bleuel et al., 2002), играющего важную роль в контроле генной экспрессии этого нейротрофина (Shieh et al., 1998; Tao et al., 1998).
Снижение экспрессии гена BDNF, обнаруженное во фронтальной коре через сутки после однократного стрессорного воздействия и в гиппокампе через 2 часа после повторного стресса, может быть, согласно нейротрофической гипотезе депрессии, рассматривающей снижение экспрессии фактора в качестве основополагающей причины возникновения депрессивных расстройств, вовлекаться в развитие индуцируемого стрессом вынужденного плавания депрессивноподобного состояния. На такую возможность указывает наличие достоверной отрицательной корреляции между уровнем мРНК BDNF в коре и продолжительностью депрессивноподобного поведения (r = -0,63, p < 0.05). В отличие от такой трактовки функциональной роли сниженного в результате стрессорного воздействия уровня BDNF в гиппокампе и фронтальной коре, последующее, через 24 часа, увеличение экспрессии фактора, учитывая его роль в контроле жизнеспособности нейрональных клеток, может иметь важное нейропротекторное значение. Это повышение может быть одним из предполагаемых процессов, активируемых для защиты мозга от повреждающих эффектов стресса (De Kloet, 2008; Dygalo et al., 2008; McEwen, 2008).
Важное нейропротекторное значение может иметь и повышение в гиппокампе экспрессии гена антиапоптозного белка Bcl-xl, обнаруженное через 24 часа после однократного воздействия и через 2 часа после повторной процедуры. Из литературных данных известно, что повышение экспрессии антиапоптозных белков семейства Bcl-2, включая и Bcl-xl, задерживало или предотвращало гибель нейронов, вызываемую адреналэктомией (Nair et al., 2007) или острым стрессом (Panickar et al., 2005).
Учитывая известные нейропротекторные свойства BDNF и Bcl-xl, повышение их экспрессии в результате стрессорного воздействия может также обусловливать устойчивость к провоцирующему депрессивноподобное состояние действию стрессоров.
Анализ возможного вовлечения генов нейропластичности, а также гормонов стресса в механизмы чувствительности или устойчивости к формированию индуцируемого стрессом вынужденного плавания депрессивноподобного состояния
Анализ поведения животных во время однократной и повторной процедуры вынужденного плавания показал, что не у всех особей развивалось в повторном тесте депрессивноподобное состояние. Так, у 12 из 30 самцов продолжительность пассивного поведения в повторном тесте не отличалась от времени аналогичного поведения в претесте (продолжительность замирания за первые 5 минут составляла при первом плавании 141.1±12.0 секунд, а при повторном - 157.0±5.5 секунд). В противоположность этим животным, названных нами стресс-устойчивыми по отсутствию характерного свидетельства развития индуцируемого вынужденным плаванием депрессивноподобного состояния, остальные 18 крыс при повторном плавании демонстрировали достоверное увеличение продолжительности этого поведения. Эти животные, ведущие себя пассивно при первом вынужденном плавании 167.4±7.7 секунд и при повторном плавании - 201.8±6.6 секунд (p < 0.001) были названы, соответственно, стресс-чувствительными. Животные этих двух групп, стресс-чувствительных, то есть демонстрирующих во втором тесте увеличение продолжительности пассивного поведения, и стресс-устойчивых, этого увеличения не демонстрирующих, были проанализированы на предмет выявления между ними возможных различий по уровням экспрессии в ответ на стресс генов BDNF, Bcl-xl и Bax в отделах мозга, а также АКТГ и кортикостерона в плазме крови. Проведенный анализ не вывил различий между группами по уровням АКТГ и кортикостерона, а также мРНК BDNF, Bax и Bcl-xl в гиппокампе и среднем мозге через 40 мин, 2 и 24 часа после воздействия. В то время в гиппокампе у стресс-устойчивых животных было обнаружено значительное повышение (по сравнению с чувствительными крысами) величины отношения мРНК Bcl-xl/Bax через 2 часа после воздействия (F1,22 = 5.154, p < 0.05, рис.5).
 | Рис. 5. Влияние теста вынужденного плавания на отношение мРНК Bcl-xl/Bax в гиппокампе стресс-чувствительных и стресс-устойчивых крыс. * - p < 0.05 по отношению к значению стресс-устойчивых животных соответствующей временной группы. |
В дополнение к мРНК антиапоптозного белка Bcl-xl, уровень которой менялся через 2 часа после повторного плавания, было исследовано, как ведет себя при примененном стрессорном воздействии сам белок Bcl-xl. В результате проведенного иммуногистохимического анализа было выявлено, что повторный стресс вынужденного плавания индуцировал усиление экспрессии антиапоптозного белка Bcl-xl в гиппокампе (F6, 20 = 5.53, p < 0.001, данные не показаны). Количество иммуноположительных клеток (на 0,1 мм2) в СА1, СА3 областях гиппокампа, а также в зубчатой извилине через 2 часа после стресса было достоверно увеличено по сравнению с нетестированными животными (CA1: контроль – 19.2 ± 8.7, 2 часа – 78.9 ± 4.9, p < 0.001; CA3: контроль – 19.6 ±7.8, 2 часа – 91.5 ±7.6, p < 0.001; зубчатая извилина: контроль – 35.2 ±14.3, 2 часа – 123.5 ±26.2, p < 0.001).
Полученные результаты позволяют предполагать, что Bcl-xl гиппокампа вовлекается в формирование индивидуальных различий в восприимчивости к стрессу, и способность животных отвечать на стресс увеличением экспрессии этого антиапоптозного белка может быть важным фактором, обусловливающим их устойчивость к развитию индуцируемого стрессом депрессивноподобного состояния. Подтверждением этого предположения служит выявленная между параметрами, отношением мРНК Bcl-xl/Bax в гиппокампе и временем пассивного поведения в повторном тесте, достоверная отрицательная корреляция (r =0.49, p < 0.05, n = 28).
Механизмы выявленных в работе индивидуальных различий в поведенческом ответе животных на повторный стресс вынужденного плавания, также как и ответе мРНК Bcl-xl/Bax на примененное стрессорное воздействие, не ясны. Неизбегаемые условия вынужденного плавания являются для грызунов стрессорными, что проявляется, помимо увеличения продолжительности пассивного поведения, значительным повышением уровня кортикостерона как в плазме крови, так и в гиппокампе (Droste, et al., 2008). Этот повышенный уровень гормонов через связывание активируемых ими глюкокортикоидных и минералокортикоидных рецепторов со специфическими глюкокортикоид-чувствительными сайтами в промоторных областях гена Bcl-X способен вызвать усиление транскрипции гена Bcl-xl (Gascoyne et al., 2003; Viegas et al., 2004). Количество же этих рецепторов, оказывающих противоположное влияние на экспрессию Bax и Bcl-xl (Almeida et al., 2000), а также уровень стрессорной гормональной активации могут быть изменены условиями ранней постнатальной жизни (Dygalo et al., 2008), например, качеством материнской заботы (Meaney, 2001; Champagne et al., 2008). Дополнительный вклад в обеспечение индивидуальных различий в чувствительности к стрессу могут вносить комплексы, формирующиеся между глюкокортикоидными рецепторами и антиапоптозным белком Bcl-xl, транслоцирующиеся в митохондрии, что, как полагают, является важным механизмом, обеспечивающим клетки энергией для борьбы с острыми стрессорными воздействиями (Du et al., 2009).
Эффекты антидепрессанта флуоксетина на экспрессию генов белков апоптоза
Для подтверждения возможного вовлечения Bcl-xl в механизмы устойчивости к развитию депрессивноподобного состояния, в следующих экспериментах исследовали эффекты антидепрессанта флуоксетина на экспрессию генов апоптоза в отделах мозга. Введение этого антидепрессанта, уменьшающего продолжительность пассивного поведения в тесте вынужденного плавания, повысило как базальный, так и в ответ на стресс уровень мРНК Bcl-xl (F1,15 = 18.01, p < 0.001, рис. 6А) в среднем мозге. Сходные изменения обнаружены в этом отделе мозга и в отношении мРНК Bcl-xl/Bax (F1,15 = 16.19, p< 0.05, рис. 6Б).
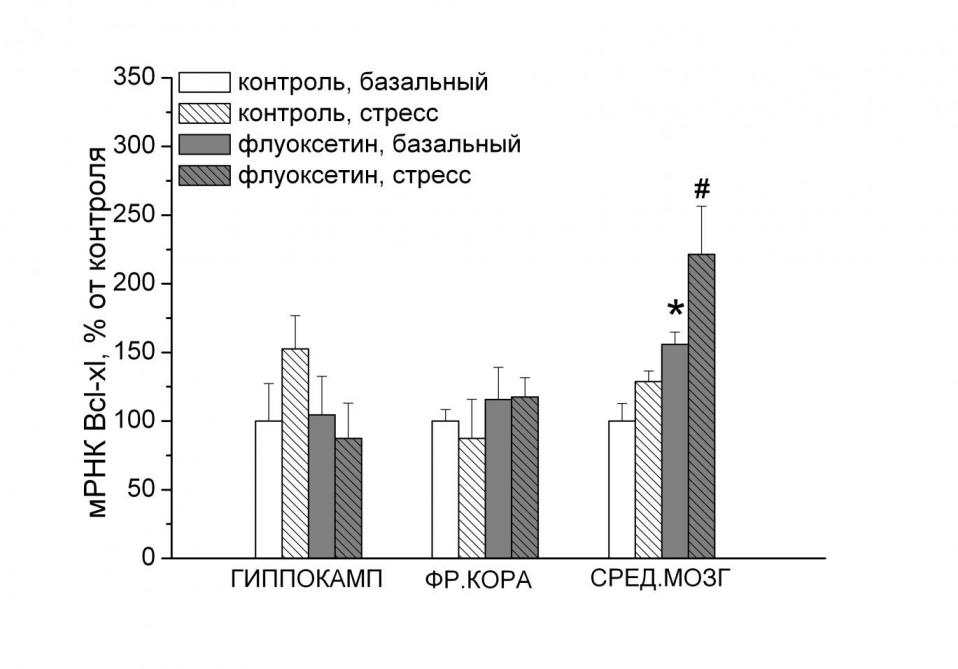 |  |
Рис. 6. Влияние повторного стресса вынужденного плавания и введения антидепрессанта флуоксетина на уровень экспрессии гена Bcl-xl (А) и Bcl-xl/Вах (Б) в отделах мозга. # - p < 0.05 по сравнению с контрольной нестрессированной группой. - p < 0.05 по сравнению со всеми остальными группами.
Выводы
- Стресс вынужденного плавания, индуцирующий у грызунов состояние поведенческой беспомощности, оказал значительное влияние на экспрессию генов мозгового нейротрофического фактора BDNF, а также ключевых регуляторов апоптоза - антиапоптозного белка Bcl-xl и проапоптозного белка Bax - в структурах мозга. Выявленные в работе эффекты зависели от отдела мозга, а также количества примененных стрессорных процедур.
- Уровень мРНК BDNF был достоверно снижен во фронтальной коре через сутки после однократного стрессорного воздействия, а также в гиппокампе и среднем мозге через 2 часа после повторной стрессорной процедуры.
- Между уровнем мРНК BDNF в коре мозга и продолжительностью пассивного поведения в тесте Порсолта обнаружена достоверная отрицательная корреляция, указывающая на возможную взаимосвязь индуцируемых стрессом изменений этих параметров.
- Стресс вынужденного плавания вызывал также активацию нейропротекторных механизмов, о чем свидетельствуют увеличение в гиппокампе мРНК BDNF через сутки после повторного стрессирования, а также Bcl-xl через сутки после однократного и 2 часа после повторного стрессирования.
- Активация антиапоптозного ответа на повторный стресс, оцененная по отношению Bcl-xl/Bax, наблюдалась в гиппокампе у животных, не проявляющих увеличения продолжительности пассивного поведения при повторном стрессировании, и отсутствовала у животных, демонстрирующих отчетливое развитие депрессивноподобного поведения. Полученные результаты являются первым свидетельством способности к увеличению экспрессии антиапоптозного белка как важного фактора, определяющего индивидуальную устойчивость или чувствительность к развитию индуцируемой стрессом депрессии.
- Антиапоптозный белок Bcl-xl может быть также мишенью действия антидепрессантных препаратов. В среднем мозге экспрессия гена этого белка после стресса была достоверно увеличена введением антидепрессанта флуоксетина.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
- Shishkina G.T., Kalinina T.S., Berezova I.V., Dygalo N.N. Stress-induced activation of the brainstem Bcl-xL gene expression in rats treated with fluoxetine: Correlations with serotonin metabolism and depressive-like behavior. // Neuropharmacology, 2012, Vol. 62, № 1, P.177-183.
- Берёзова И.В., Шишкина Г.Т., Калинина Т.С., Дыгало Н.Н. Поведение в тесте вынужденного плавания и экспрессия в мозге крыс генов нейротрофического фактора (BDNF) и антиапоптозного белка Bcl-xl. // Журнал высшей нервной деятельности, 2011, Т. 61, №3, C. 332-339.
- Булыгина В.В., Шишкина Г.Т., Берёзова И.В., Дыгало Н.Н. Экспрессия белка мозгового нейротрофического фактора в гиппокампе крыс после стресса вынужденного плавания. // Доклады Российской академии наук, 2011, Т. 437, C. 82-84.
- Shishkina G.T., Kalinina T.S., Berezova I.V., Bulygina V.V., Dygalo N.N. Resistance to the development of stress-induced behavioral despair in the forced swim test associated with elevated hippocampal Bcl-xl expression. // Behav. Brain Res., 2010, Dec 1, 213(2), P. 218-224.
- Shishkina G.T., Kalinina T.S., Berezova I.V., Dygalo N.N. Stress-induced increase of anti-apoptotic protein, Bcl-xl, gene expression in the brainstem of fluoxetine-treated rats correlates with monoamine metabolism. // Abstracts of 8th IBRO World Congress of Neuroscience, 2011, P. A500 (Italy, 2011).
- Shishkina G.T., Kalinina T.S., Berezova I.V., Dygalo N.N. Effects of the forced swim test and fluoxetine pretreatment on brain expression of genes related to neuronal plasticity. // European Neuropsychopharmacology, 2011, Vol. 21 Suppl. 3, P. S266 (papers of 24th ECNP Congress, France, 2011).
- Dygalo N.N., Kalinina T.S., Menshanov P.N., Berezova I.V., Bulygina V.V., Shishkina G.T. Pro- and anti-apoptotic effects of stress and glucocorticoids in the brain: functional implications. // Endocrine regulations, 2011, Vol. 45, № 2, P. A13 (Abstracts of 10th Symposium on Catecholamines and Other Neurotransmitters in Stress, Slovakia, 2011).
- Берёзова И.В., Дыгало Н.Н., Калинина Т.С., Шишкина Г.Т. Активация антиапоптозных механизмов в среднем мозге крыс в послестрессорный период усиливается на фоне хронического введения антидепрессанта флуоксетина. // Материалы Пятой Всероссийской научно-практической конференции с международным участием «Фундаментальные аспекты компенсаторно-приспособительных процессов», 2011, C. 25 (Новосибирск, 2011).
- Берёзова И.В., Шишкина Г.Т., Калинина Т.С., Дыгало Н.Н. Стресс вынужденного плавания изменяет экспрессию генов нейропластичности. // Тезисы докладов XXI Съезда физиологического общества имени И.П. Павлова, 2010, C. 66 (г. Калуга, 2010).
- Шишкина Г.Т., Калинина Т.С., Берёзова И.В., Булыгина В.В., Дыгало Н.Н. Механизмы устойчивости к развитию индуцируемой стрессом депрессии. // Тезисы докладов XXI Съезда физиологического общества имени И.П. Павлова, 2010, C.700 (г. Калуга, 2010).
- Shishkina G.T., Kalinina T.S., Berezova I.V., Bulygina V.V., Dygalo N.N. Expression of genes associated with cell proliferation and survival in the rat brain after forced swim stress exposure: possible implication in the behavioral immobility response. // Abstracts of 7th World Congress on Stress, 2010, P. 22-23 (The Netherlands, Amsterdam, 2010).
- Тыдыков Л.В., Берёзова И.В. Влияние острого стресса на экспрессию мозгового нейротрофического фактора и антиапоптозного белка Bcl-xl в гиппокампе крыс. // Материалы XLVIII МНСК "Студент и научно-технический прогресс", 2010, C. 53 (Новосибирск, НГУ, 2010).
- Берёзова И.В., Шишкина Г.Т. Изменение гормональной активности коры надпочечников и половых желез при длительном введении антидепрессанта флуоксетина. // Тезисы докладов VII Всероссийской конференции с международным участием "Механизмы функционирования висцеральных систем", 2009, C. 55 (Санкт-Петербург, 2009).
- Шишкина Г.Т., Калинина Т.С., Берёзова И.В. Психонейроэндокринные корреляты фармакологической модуляции экспрессии генов в мозге. // Материалы V съезда Вавиловского общества генетиков и селекционеров, 2009, C. 136 (Москва, 2009).
- Тыдыков Л.В., Берёзова И.В. Влияние стресса и глюкокортикоидов на экспрессию гена мозгового нейротрофического фактора в гиппокампе. // Материалы XLVII МНСК "Студент и научно-технический прогресс" 2009, C. 39 (Новосибирск, НГУ, 2009).
- Берёзова И.В. Участие дофамина мозга в психоэмоциональных эффектах флуоксетина. // Материалы XLVI МНСК "Студент и научно-технический прогресс", 2008, C. 74 (г. Новосибирск, НГУ, 2008).