

Применение стволовых клеток при возрастной нейродегенеративной патологии ( экспериментальное исследование )
На правах рукописи
АНИСИМОВ
Сергей Владимирович
Применение стволовых клеток
ПРИ возрастной
нейродегенеративной патологии
(экспериментальное исследование)
14.01.30 - геронтология и гериатрия
03.03.04 - клеточная биология, цитология, гистология
А в т о р е ф е р а т
диссертации на соискание ученой степени
доктора медицинских наук
Санкт-Петербург - 2011
Работа выполнена в Федеральном Центре сердца, крови и эндокринологии им. В.А.Алмазова», Геронтологическом исследовательском центре Национального института старения США (г. Балтимор, США) и Валленберговском Нейрологическом Центре Люндского Университета (г. Люнд, Швеция).
Научные консультанты:
академик РАН,
доктор биологических наук, профессор
Никольский Николай Николаевич
профессор, M.D., Ph.D.
Брюндин Патрик (Швеция)
Официальные оппоненты:
заслуженный деятель науки РФ,
доктор медицинских наук, профессор
Кветной Игорь Моисеевич
заслуженный деятель науки РФ,
академик РАМН,
доктор медицинских наук, профессор
Скоромец Александр Анисимович
доктор медицинских наук, профессор
Имянитов Евгений Наумович
Ведущее научное учреждение:
Учреждение Российской академии наук «Институт мозга человека им. Н. П. Бехтеревой» РАН
Защита диссертации состоится "_____"____________2011 г. в ____ часов
на заседании диссертационного совета Д601.001.01 при Санкт-Петербургском институте биорегуляции и геронтологии СЗО РАМН по адресу: 197110, Санкт-Петербург, пр. Динамо, д. 3
С диссертацией можно ознакомиться в библиотеке Санкт-Петербургского института биорегуляции и геронтологии СЗО РАМН (197110, Санкт-Петербург, пр. Динамо,
д. 3).
Автореферат разослан "_____" ______________2011 г.
Ученый секретарь
диссертационного совета
доктор биологических наук, профессор Козина Л.С.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы
Проблема постарения населения, то есть, увеличение в структуре населения доли лиц пожилого возраста в последние десятилетия приобретает глобальный характер.
Об актуальности проблемы старения для всего человечества свидетельствует разработанная по инициативе ООН «Программа по исследованиям старения в ХХI веке». Отдельный раздел этой программы посвящен изучению механизмов старения и связанных с ним заболеваний [Сидоренко А. и Эндрюс Г., 2000]. Среди этих заболеваний одно из важнейших мест занимают нейродегенеративные. Хотя нейродегенеративный процесс может быть обусловлен генетической патологией (наследственные метаболопатии), являться осложнением инфекционных заболеваний, травм или воздействия токсических факторов, важнейшее значение имеет широкая распространенность именно возрастной идиопатической нейродегенеративной патологии [Литвиненко И. В., 2006]. Тремя наиболее распространёнными нейродегенеративными заболеваниями в мире являются болезнь Альцгеймера (БА), болезнь Паркинсона (БП), и болезнь [диффузных] телец Леви (БДТЛ), соответственно. Важно отметить, что в то время как для БА и БДТЛ характерна диссеминация нейродегенеративного процесса (с поражением нейронов коры головного мозга в целом), при БП нейродегенеративный процесс преимущественно локализован. Как хроническое неуклонно прогрессирующее заболевание ЦНС, БП сопровождается дегенерацией нигростриарных нейронов и нарушением функции базальных ганглиев.
За исключением наследственных (ювенильных) форм, БП поражает лиц пожилого возраста, при этом заболеваемость и распространенность болезни Паркинсона неуклонно увеличиваются с возрастом: от ~1% среди людей старше 60 лет до ~2% среди людей старше 70 лет [Langston J. W., 1998]. Продолжительная (5-10 лет) фармакотерапия связана
с появлением значимых побочных реакций и общим снижением эффективности действия.
В связи с этим, в течение последних 18-20 лет производятся попытки создать принципиально новые методы лечения БП, - такие, как глубокая стимуляция головного мозга и заместительная клеточная терапия дофаминергическими нейронами донорского происхождения [Бехтерева Н. П. и др., 1990; Hitchcock E. R., et al., 1988; Lindvall O., et al., 1988; Madrazo I., et al., 1988]. Стволовые клетки (СК) разных типов способны служить ценным источником нейронов как субстрата клеточной терапии нейродегенеративных
и иных заболеваний [Дыбан А. П. и Дыбан П. А., 2002; Никольский Н. Н. и др., 2007; Зинькова Н. Н. и др., 2007]. Наиболее перспективным субстратом заместительной клеточной терапии локализованной возрастной нейродегенеративной патологии считаются эмбриональные стволовые клетки (ЭСК), однако существующие протоколы их дифференцировки в функциональные дофаминергические нейроны являются крайне длительными и чрезвычайно сложными технически [Carpenter M. K., et al., 2001; Kim J. H., et al., 2002]. Кроме того, сам механизм дифференцировки, то есть перестройки плюрипотентной ЭСК в высокоспециализированный нейрон является весьма сложным и в значительной степени необъяснённым [Iacovitti L., et al., 2007]. Для фактического применения данного подхода в клинической практике необходима разработка эффективного протокола, основанного на экспансии недифференцированных ЭСК in vitro, дифференцировки их в функциональные дофаминергические нейроны, а также обеспечение безопасности субъектов трансплантации (в первую очередь, с точки зрения развития злокачественной тератомы), поскольку пожилой и старческий возраст является фактором, способствующим промоции и прогрессии канцерогенеза. Важнейшими направлениями данного исследования являлись раскрытие механизмов пролиферации и дифференцировки ЭСК человека, а также обеспечение ксенологической и онкологической безопасности трансплантации и повышение её эффективности за счёт модификации протоколов дифференцировки и смены формата селекции пересаживаемого клеточного материала.
Цель исследования
Изучить механизмы пролиферации и дифференцировки стволовых клеток,
и разработать способ заместительной клеточной терапии локализованной, ассоциированной с возрастом нейродегенеративной патологии в условиях эксперимента.
Задачи исследования
1. Провести анализ возможности проведения клеточной терапии ассоциированной с возрастом нейродегенеративной патологии и идентифицировать действенный субстрат клеточной терапии;
2. Выявить факторы, определяющие возможность эффективного размножения стволовых клеток в культуре in vitro;
3. Изучить возможность направленной дифференцировки эмбриональных стволовых клеток человека в функциональные дофаминергические нейроны в культуре in vitro;
4. Разработать стандартизованный протокол, позволяющий получить из эмбриональных стволовых клеток человека оптимальное количество функциональных дофаминергических нейронов, пригодных для терапии, ассоциированной с возрастом нейродегенеративной патологии;
5. Оценить влияние фактора роста фибробластов 20 (FGF-20) на клетки-предшественники дофаминергических нейронов;
6. Разработать технологию подготовки клеточного материала к трансплантации, обеспечивающую максимальную выживаемость пересаженных клеток и эффективность лечения ассоциированной с возрастом нейродегенеративной патологии;
7. Разработать способ предотвращения развития злокачественной тератомы в сайте трансплантации клеточного материала при терапии, ассоциированной с возрастом нейродегенеративной патологии;
8. Изучить эффекты трансплантации производных эмбриональных стволовых клеток человека лабораторным животным, модельным в отношении локализованной, ассоциированной с возрастом нейродегенеративной патологии.
Научная новизна исследования
Выявлена биологическая роль фактора роста фибробластов 20 (FGF-20), который был идентифицирован ранее, но биологическая роль которого до сих пор оставалась неизвестной. В ходе проведения экспериментов по направленной дифференцировке чЭСК в нейроны in vitro было установлено, что фактор роста фибробластов 20 стимулирует дифференцировку и поддерживает выживаемость дофаминергических прогениторных клеток, что может способствовать повышению эффективности клеточной терапии ассоциированных с возрастом нейродегенеративных заболеваний.
Впервые установлена существование обратной зависимости между временем пребывания чЭСК в культуре in vitro (пропорциональным доле постмитотических клеток в общем объеме гетерогенной популяции производных чЭСК) и риском развития злокачественной тератомы (тератокарциномы) в сайте трансплантации клеточного материала.
Сформулирована концепция «окна возможности» проведения заместительной клеточной терапии локализованной возрастной нейродегенеративной патологии, в пределах которого в культуре in vitro можно получить достаточное количество дофаминергических нейронов-производных чЭСК, и при этом выживаемость донорских дофаминергических нейронов в ходе трансплантационной процедуры является приемлемой, а риск развития злокачественной тератомы (тератокарциномы) в сайте/сайтах трансплантации клеточного материала отсутствует, что особенно важно при последующем применении заместительной клеточной терапии у пациентов пожилого и старческого возраста.
Разработан и использован в экспериментальной работе двухэтапный протокол направленной дифференцировки чЭСК в функциональные дофаминергические нейроны in vitro, который позволяет получить однородную популяцию содержащих такие клетки клеточных агрегатов, пригодных к трансплантации непосредственно, то есть без диссоциации. Разработана также техника трансплантации таких клеточных агрегатов. Установлено, что использование этой технологии многократно повышает выживаемость донорских дофаминергических нейронов в сайте трансплантации, одновременно полностью ликвидируя риск развития злокачественной тератомы (тератокарциномы) у реципиентов трансплантации.
Разработана технология, позволяющая проводить эффективное размножение эмбриональных стволовых клеток человека (чЭСК) в культуре in vitro (экспансию чЭСК) в сложных клеточных системах, основанных на применении фидерных клеток человеческого происхождения.
Проведена комплексная оценка параметров транскриптомов стволовых клеток разных типов в сравнении с транскриптомами иных типов клеток (не стволовых). В результате проведенного анализа выявлены важные закономерности и идентифицирована панель молекулярных мишеней, играющих важную роль в контроле пролиферации и дифференцировки стволовых клеток.
Разработана, запущена в опытное производство и начала применяться в экспериментальной биологии специализированная микрочиповая (микроматричная) платформа «NeuroStem» предназначенная для изучения транскриптома стволовых клеток человека в ходе дифференцировки в клетки нейрального/нейронального ряда. Данная микрочиповая платформа обладает рекордными характеристиками в отношении представленности на ней молекулярных мишеней, имеющих отношение к биологии дофаминергической системы, в значительной степени способствуя прогрессу в разработке надежных, безопасных и эффективных методов клеточной терапии ассоциированных с возрастом нейродегенеративных заболеваний.
Научно-практическое значение
Применение технологии размножения эмбриональных стволовых клеток человека (чЭСК) в культуре in vitro (экспансии чЭСК) в сложных клеточных системах, основанных на применении фидерных клеток человеческого происхождения снижает влияние ксеногенных факторов на клетки человека, что снижает ксеногенные риски в случае практического применения производных этих клеток в формате клеточной терапии, понижает влияние ксеногенных факторов на параметры транскриптома и протеома клеток человека, что важно для верной интерпретации полученных научных результатов. Кроме того, применение для экспансии чЭСК клеток человеческого происхождения является значительно более удобным по сравнению с клетками мыши.
Проведенный широкомасштабный анализ транскриптомов стволовых клеток разных типов в сравнении с транскриптомами иных типов клеток позволил идентифицировать панель молекулярных мишеней, играющих важную роль в контроле пролиферации и дифференцировки стволовых клеток. Отдельные молекулярные мишени из состава данной группы могут быть использованы в качестве мишени генной модификации клеток
(с использованием технологии генной инженерии) для воздействия на процессы пролиферации и дифференцировки стволовых клеток разных типов, - в том числе
в интересах генной и генно-клеточной терапии ассоциированных с возрастом заболеваний.
Разработанная, апробированная и запущенная в производство специализированная микрочиповая (микроматричная) платформа «NeuroStem» позволяет с высокой производительностью и высоким качеством исследовать транскриптом стволовых клеток человека, нейронов и глиальных клеток разных типов, а также динамику транскриптома стволовых клеток человека в ходе дифференцировки в клетки нейрального/нейронального ряда. Это позволяет получать ценную информацию о фундаментальных механизмах, лежащих в основе жизнедеятельности стволовых клеток, нейронов, и их коммитированных предшественников. В свою очередь, это может иметь большое значение для идентификации новых диагностических, прогностических и терапевтических мишеней терапии ассоциированных с возрастом нейродегенеративных заболеваний.
Применение фактора роста фибробластов 20 (FGF-20) в качестве добавки в составе питательной среды используемой для культивирования производных стволовых клеток человека in vitro в ходе их направленной дифференцировки в дофаминергические нейроны позволяет повысить эффективность исходного протокола, что определяет возможность получения из чЭСК большое количество жизнеспособных и функциональных дофаминергических нейронов для использования в исследовательской работе и в качестве субстрата заместительной клеточной терапии локализованной возрастной нейродегенеративной патологии.
Установленная обратная зависимость между временем пребывания чЭСК в культуре in vitro и риском развития злокачественной тератомы (тератокарциномы) в сайте трансплантации клеточного материала, и сформулированная концепция «окна возможности» проведения заместительной клеточной терапии локализованной возрастной нейродегенеративной патологии позволили адаптировать существующие подходы к основанной на применении стволовых клеток терапии, минимизировав риск развития злокачественной тератомы (тератокарциномы) в сайте/сайтах трансплантации клеточного материала донорского происхождения.
Разработанный и апробированный в эксперименте двухэтапный протокол направленной дифференцировки чЭСК в функциональные дофаминергические нейроны in vitro позволяет получить однородную популяцию содержащих такие клетки клеточных агрегатов, что даёт возможность избежать проведения этапа диссоциации клеток при проведении трансплантации. Использование такой технологии многократно повышает выживаемость донорских дофаминергических нейронов в сайте трансплантации, что открывает новые перспективы перед подходами к заместительной клеточной терапии локализованной возрастной нейродегенеративной патологии. Разработанная технология будет способствовать дальнейшему прогрессу в создании безопасных и удовлетворяющих клиническим требованиям патогенетически обоснованных методов лечения ассоциированных с возрастом заболеваний, сопровождающихся локализованным нейродегенеративным процессом.
Положения, выносимые на защиту
1. Сравнительный анализ ключевых биологических свойств стволовых клеток разных типов показывает, что эмбриональные стволовые клетки человека (чЭСК) являются наиболее адекватным субстратом для проведения заместительной клеточной терапии локализованной возрастной нейродегенеративной патологии.
2. Анализ транскриптомов стволовых клеток разных типов (в том числе недифференцированных и находящихся на различных этапах дифференцировки)
с использованием технологии серийного анализа генетической экспрессии (SAGE) и микрочиповой технологии демонстрирует практическую невозможность действенной стимуляции дифференцировки стволовых клеток в функциональные дофаминергические нейроны методами генетической инженерии.
3. Разработана технология эффективной направленной дифференцировки эмбриональных стволовых клеток человека в функциональные дофаминергические нейроны in vitro; впервые установлен вклад фактора роста фибробластов 20 (FGF-20)
в дифференцировку и выживаемость дофаминергических прогениторных клеток. Применение FGF-20 позволило пятикратно повысить эффективность исходного протокола в приложении к выходу дофаминергических нейронов.
4. В экспериментальных моделях локализованной ассоциированной с возрастом нейродегенеративной патологии установлено существование обратной зависимости между риском развития злокачественной тератомы и временем пребывания клеток в культуре
in vitro.
5. Предложена концепция «окна возможности» проведения заместительной клеточной терапии локализованной возрастной нейродегенеративной патологии, в пределах которого выживаемость донорских дофаминергических нейронов является приемлемой, а риск развития злокачественной тератомы в сайте/сайтах трансплантации клеточного материала отсутствует.
6. Разработанный и использованный двухэтапный протокол направленной дифференцировки эмбриональных стволовых клеток человека в функциональные дофаминергические нейроны in vitro позволяет получить однородную популяцию содержащих такие клетки клеточных агрегатов, пригодных к трансплантации непосредственно, без диссоциации. Использование этой технологии более чем в 5 раз повышает выживаемость донорских дофаминергических нейронов в сайте трансплантации, одновременно полностью устраняя риск развития злокачественной тератомы у реципиентов.
Апробация работы
Основные результаты исследований и положения диссертационной работы, выносимые на защиту, были представлены и доложены на следующих научных конференциях, съездах и практикумах: III, V, VI Annual National Institute on Aging Intramural Scientific Retreat (Линтицум Хэйтс, США, 1998, 2000, 2001); Stem Cells: Origins, Fates and Functions, Keystone Symposia (Кейстоун, США, 2002); From Stem Cells to Therapy, Keystone Symposia (Стимбоат Спрингс, США); INYS Workshop on Stem Cell Research. Nobel Forum (Стокгольм, Швеция, 2004); 4th Forum of European Neuroscience (Лиссабон, Португалия, 2004); Symposium of Lund Strategic Center for Stem Cell Biology and Cell Therapy (Люнд, Швеция, 2005); Meeting of SRC-SDARF-JDRF Joint Programme in Stem Cell Research (Сигтуна, Швеция, 2005); International Society for Stem Cell Research 3rd Annual Meeting
(Сан-Франциско, США, 2005); From Neuron to Neurology and Psychiatry: The Baltic Summer School (Копенгаген, Дания, 2005); 2nd European Science Foundation Functional Genomics Conference “Functional Genomics and Disease” (Осло, Норвегия, 2005); NSR (Nervous System and Repair) and NCoE (Nordic Center of Excellence) Joint Symposium: Disease models in vivo and in vitro (Люнд, Швеция, 2005); Swedish Research Council NeuroFortis (Strong Research Environment in Brain Damage and Repair) Annual Meeting (Изаберг, Швеция, 2009); General Biology of stem cell systems: EuroSTELLS Meeting (Венеция, Италия, 2006); Pre-clinical Evaluation of Stem Cell Therapy in Stroke (STEMS) kick-off meeting (Париж, Франция, 2006), II Съезд Общества клеточной биологии и Юбилейная конференция, посвященная 50-летию Института цитологии РАН (Санкт-Петербург, 2007); IV и V Международная конференция «Медицина долголетия и качества жизни» (Москва, 2008, 2009); Всероссийская научно-практическая конференция c международным участием «Высокотехнологичные методы диагностики и лечения заболеваний сердца, крови и эндокринных органов» (Санкт-Петербург, 2008, 2009); Российско-шведский практикум «Cell Technologies for Medicine» (Санкт-Петербург, 2009); Всероссийская научная школа для молодёжи «Аутологичные стволовые клетки: экспериментальные и клинические исследования» (Москва, 2009); Итоговая конференция по результатам выполнения мероприятий за 2009 г. в рамках приоритетного направления «Живые системы» ФЦП «Исследования и разработки по приоритетным направлениям развития научно-технологического комплекса России на 2007-2012 годы» (Москва, 2009); LXXI конференция молодых ученых СПбГМУ им. акад. И.П.Павлова (Санкт-Петербург, 2010), Всероссийской научно-практической конференции «Актуальные нейродегенеративные заболевания XXI века» (Санкт-Петербург, 2010).
Публикации
По материалам диссертации опубликовано 18 статей в журналах, рекомендуемых ВАК Российской Федерации для публикации результатов докторских исследований, 4 статьи в иных журналах, 5 глав в коллективных монографиях, 2 главы в методических пособиях, 12 тезисов докладов.
Внедрение
Полученные в ходе диссертационной работы данные используются в научно-исследовательской работе и образовательном процессе в ФГУ «Федеральный Центр сердца, крови и эндокринологии им. В.А.Алмазова» Министерства здравоохранения и социального развития Российской Федерации, Учреждении РАН Институте цитологии РАН, Институте эволюционной физиологии и биохимии им. И.М.Сеченова РАН, и Санкт-Петербургском Институте биорегуляции и геронтологии СЗО РАМН, Национальном институте старения США, и Люндском Университете (Швеция). Разработанная в ходе выполнения данной работы и производимая компанией «SweGene» (Швеция) специализированная микрочиповая (микроматричная) платформа «NeuroStem» в настоящее время используется в работе лабораторий клеточной биологии и экспериментальной неврологии Люндского Университета (Швеция) и Университета Тель-Авива (Израиль) для экспериментальной работы, направленной на разработку методов терапии ассоциированной с возрастом нейродегенеративной патологии, основанной на применении эмбриональных и соматических стволовых клеток человека.
Структура и объем работы
Диссертация состоит из введения, обзора литературы, описания материалов и методов работы, собственных результатов и обсуждения, и выводов. Объем работы составляет 429 страниц. Список литературы содержит 29 отечественных и 560 зарубежных источников. Диссертация иллюстрирована 16 таблицами и 93 рисунками.
СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы исследования
Для изучения механизмов пролиферации и дифференцировки стволовых клеток, и разработки способа заместительной клеточной терапии локализованной, ассоциированной с возрастом нейродегенеративной патологии использовались стволовые и фидерные клетки животных и человека, и экспериментальная модель локализованной возрастной патологии, а именно:
Эмбриональные стволовые клетки мыши (мЭСК) линии R1 культивировали на фидерных (питающих) клетках - митотически инактивированных первичных культурах эмбриональных фибробластов мыши, в желатинизированных чашках Петри. Экспансию мЭСК линии R1 проводили в питательной среде на основе среды Игла в модификации Дюльбекко с высоким содержанием глюкозы (DMEM high glucose, Gibco BRL, США), с 15% сыворотки. Пассажи осуществляли энзиматически. Бесфидерные культуры мЭСК линии R1 вели с использованием среды BRL-CM «приспособленной» (кондиционированной) на культуре клеток мышиной печени (линия BRL) полученной из коммерческого источника (American Type Culture Collection, ATCC, США), с добавлением 1000 Ед/мл лейкемического ингибиторного фактора (LIF, Sigma, США или Chemicon, США), 0,01 мг/мл трансферрина (Sigma, США), и 0,45 мМ монотиоглицерина (Sigma, США).
Эмбриональные стволовые клетки мыши (мЭСК) линии D3 (American Type Culture Collection, ATCC, США) культивировали на фидерных клетках - митотически инактивированных первичных культурах эмбриональных фибробластов мыши,
в желатинизированных чашках Петри в присутствии лейкемического ингибиторного фактора (LIF, Sigma, США или Chemicon, США). Экспансию мЭСК линии R1 проводили
в питательной среде на основе среды Игла в модификации Дюльбекко с высоким содержанием глюкозы (DMEM high glucose, Gibco BRL, США), с 10% сыворотки. Пассажи осуществляли энзиматически.
Клетки эмбриональной карциномы (ЭК) мыши линии P19 культивировали
в бесфидерной системе. Экспансию клеток ЭК линии Р19 проводили в питательной среде на основе среды Игла в модификации Дюльбекко с высоким содержанием глюкозы (DMEM high glucose, Gibco BRL, США), содержащей 10% сыворотки. Пассажи клеток ЭК линии Р19 осуществляли энзиматически.
Полученные из первичных половых клеток (ППК) стволовые клетки мыши линии EG-1 культивировали на фидерных клетках - митотически инактивированных первичных культурах эмбриональных фибробластов мыши, в желатинизированных чашках Петри
в присутствии LIF (Sigma, США или Chemicon, США). Экспансию клеток линии EG-1 проводили в питательной среде на основе среды Игла в модификации Дюльбекко с высоким содержанием глюкозы (DMEM high glucose, Gibco BRL, США), содержащей 15% сыворотки, 20 нг/мл LIF (Sigma, США или Chemicon, США) и 10 нг/мл лиганда с-Kit (BMA Biomedicals, Швейцария) также известного как soluble steel factor (SF), фактор стволовых клеток (SCF, также ростовой фактор тучных клеток (MGF)), KIT лиганд (KITLG, KL-1, Kitl)
и DKFZp686F2250. Пассажи клеток линии EG-1 осуществляли энзиматически.
Эмбриональные стволовые клетки человека (чЭСК) линии SA002 (также известна как SA02) были предоставлены компанией Cell Therepeutics Scandinvia / Cellartis AB (Швеция). Клетки культивировали на фидерных клетках - митотически инактивированных культурах эмбриональных фибробластов мыши или митотически инактивированных культурах неонатальных фибробластов крайней плоти человека, в желатинизированных плашках для искусственного оплодотворения (IVF-плашках). В качестве питательной среды для культивирования чЭСК линии SA002 использовали готовую питательную среду VitroHES (VitroLife, Швеция). Дополнительно в питательную среду добавлялся основной фактор роста фибробластов (bFGF, FGF-2) (R&D Systems, США) в концентрации 4 нг/мл. Пассажи осуществляли каждые 6 дней механически, с использованием стерильных клеточных ножей (диаметром 300 мкм, реже 200 мкм) и клеточных пипеток (диаметром
200 мкм, реже 300 мкм) (SweMed, Швеция) под 40-кратным увеличением инвертированного микроскопа CKX41 (Olympus, США), на установленной в манипулятор нагревательной площадке MATS-CKTS (вариант с утоньшением центральной части площадки) (Tokai Hit, Япония) с установкой температуры 42оС. Все манипуляции проводили исключительно под контролем электронного таймера, длительность пребывания IVF-плашек с чЭСК вне инкубатора во всех случаях не превышала 6 минут ровно. В ходе механических пассажей чЭСК линии SA002, использовали колонии чЭСК без признаков спонтанной дифференцировки и массовой клеточной, что указывалось следующими основными показателями: 1) полигональная или каплевидная форма колоний чЭСК, 2) отсутствие
в пределах колоний фокусов темного (темно-серого, желтого, бурого) или светлого (очень светло-серого) цветов, 3) четкое контрастирование краев колоний на фоне фидерных клеток. В ходе экспансии использовали заморозку клеток чЭСК линии SA002 методом витрификации (остекленения) в соломинках, в целом согласно протоколу [Reubinoff B. E.
et al., 2001]. При этом использовали крио-среды на основе человеческого сывороточного альбумина, трехалозы, этиленгликоля и диметилсульоксида (ДМСО).
Эмбриональные стволовые клетки человека (чЭСК) линии HUES-3 были предоставлены кафедрой молекулярной и клеточной биологии Гарвардского университета (Кембридж, Массачусетс, США). Клетки культивировали на фидерных клетках - митотически инактивированных культурах эмбриональных фибробластов мыши или митотически инактивированных культурах неонатальных фибробластов крайней плоти человека, в желатинизированных чашках Петри. В качестве питательной среды для культивирования чЭСК линии HUES-3 использовали стерилизованную фильтрованием питательную среду Игла в модификации Дюльбекко (DMEM, Invitrogen, США), содержащую 10% плазманата (Plasmanate, Green Cross Corp., Япония). Дополнительно в питательную среду добавляли bFGF в концентрации 4 нг/мл. Пассажи чЭСК линии HUES-3 осуществляли каждые 4 дня (для культивирования на культурах эмбриональных фибробластов мыши) или 6 дней (для культивирования на культурах неонатальных фибробластов крайней плоти человека), энзиматически.
Эмбриональные стволовые клетки человека (чЭСК) линии #237 были предоставлены отделением акушерства и гинекологии госпиталя Хаддинге (Karolinska University Hospital Huddinge) (Стокгольм, Швеция). чЭСК линии #237 культивировали на фидерных клетках - митотически инактивированных культурах эмбриональных фибробластов мыши или митотически инактивированных культурах неонатальных фибробластов крайней плоти человека, в желатинизированных IVF-плашках, как описано выше. В качестве питательной среды для культивирования чЭСК линии #237 использовали стерилизованную фильтрованием питательную среду Игла в модификации Дюльбекко в варианте «Нокаут» (КО-DMEM, также DMEM КО, Invitrogen, США), содержащую 20% заменителя сыворотки (Invitrogen, США). Дополнительно в питательную среду добавляли bFGF в концентрации 4 нг/мл. Пассажи чЭСК линии #237 осуществляли механически как описано выше для чЭСК линии SA002, или энзиматически при помощи обработки диспазой. В ходе экспансии использовали заморозку клеток методом витрификации в соломинках,
в целом согласно описанному выше методу, но с использованием крио-среды на основе
4-(2-оксиэтил)1-пиперазинэтансульфоновой кислоты (HEPES, Invitrogen, США), заменителя сыворотки, этиленгликоля, сахарозы и ДМСО.
В качестве фидерных (питающих) клеток использовали эмбриональные фибробласты мыши (мЭФ) или фибробласты крайней плоти новорожденных (чНФ). Первичную культуру мЭФ получали из диссоциированных тканей мышиных эмбрионов
(3 недели) линии С57BL/6; экспансию проводили в питательной среде на основе среды Игла в модификации Дюльбекко с высоким содержанием глюкозы (DMEM high glucose, Gibco BRL, США), с 10% сыворотки. Культуру неонатальных фибробластов крайней плоти человека (линия CCD-1112Sk) получали из коммерческого источника (American Type Culture Collection, ATCC, США; региональный центр в Голландии). Для пролиферации клеток использовали питательную среду на основе среды Искове (Iscove’s medium, IMDM; Invitrogen, США), с 10% сыворотки. Антимитотическую обработку мЭФ проводили митомицином С или гамма-радиацией (мЭФ, 30 Гр; чНФ, 40 Гр) с использованием экранированного облучателя Gammacell 1000 (MDS Nordion, Канада). Стромальные клетки мыши линии РА6 (также известны как MC3T3G2; RICKEN, Япония) культивировали
в стандартных условиях в питательной среде на основе минимальной среды Игла
в модификации альфа (-МЕМ); митотическую инактивацию не проводили.
Дифференцировку стволовых клеток мыши разных линий запускали методом «висячей капли». Метод висячей капли основан на приготовлении серийных разведений суспензии стволовых клеток в специфической концентрации, 20-мкл капли которых инкубируются в течение 2 суток в присутствии ДМСО. Сформировавшиеся «эмбриональные тельца» доращиваются в течение 1 суток в большом объеме суспензионной культуры,
и высаживаются на желатиновую подложку.
Дифференцировку чЭСК в функциональные дофаминергические нейроны проводили с использованием протокола SDIA, основанном на совместном культивировании (ко-культивировании) чЭСК и стромальных клеток мыши в сложной клеточной системе
в течение 14-23 дней. Фактор роста фибробластов 20 (FGF-20) использовался в протоколах направленной дифференцировки чЭСК в концентрации 100 пг/мл - 200 нг/мл питательной среды.
Формирование агрегатов производных чЭСК проводили отбором спонтанно формирующихся эмбриональных телец, с использованием метода «висячей капли» описанного выше, на основе фрагментов колоний чЭСК, и с применением самостоятельно разработанного двухэтапного метода, позволяющего получить гомогенную популяцию «трансплантационных единиц» (Transplant Units, TU) с заданными свойствами. Разработанная техника создания «трансплантационных единиц» основана на последовательном применении двух технологий: 1) нейрональной дифференцировки чЭСК по протоколу SDIA и 2) культивировании коммитированных клеток в суспензионной культуре. На первом этапе работы, культивирование линий чЭСК и стромальных клеток мыши линии РА6 в сложной клеточной системе производилось в течение 14-18 дней. После завершения этапа, производилось отделение колоний производных чЭСК от подложки (стромальных клеток) при помощи обработки папаином (Worthington Biochemical Corporation, США). Инкубацию клеток с ферментом сочетали с механической обработкой,
с соблюдением технологии известной как «метод звездного одеяла», и последующей диссоциацией клеток при помощи аккутазы (Innovative Cell Technologies, США). Клетки концентрировали центрифугированием и ресуспинзировали в питательной среде
NS (Neurosphere Medium), состоящей из питательной среды N2 (Neurobasal Medium, Invitrogen, США), содержащей 2,0% заменителя сыворотки (реагент В27),
0,5 мМ L-глутамина и 4 нг/мл основного фактора роста фибробластов (FGF-2). Варинатами протокола являлись использование приспособленной на клетках линии PA6 среды NS
(среды CNS), а также фактора роста фибробластов 20 (FGF-20).
Диссоциированные клетки (производные этапа 1 направленной дифференцировки чЭСК в дофаминергические нейроны) исследовали в отношении жизнеспособности при помощи окраски трипановым синим (TrBlue, Sigma, США), после чего разводили питательной средой до концентрации 50 000-500 000 жизнеспособных кл/мл. При этом испытывались следующие варианты концентрации: 50 000, 100 000, 150 000, 200 000, 250 000, 500 000 кл/мл. 2 мл клеточной суспензии (то есть 100 000-1 000 000 жизнеспособных клеток) переносили в чашки Петри диаметром 35 мм с высокой адгезивностью (Nalgene, Дания), и инспектировали под микроскопом. Используя стерильные носики для механических пипеток, из чашек Петри (то есть из клеточной суспензии) удаляли все замеченные аггрегаты клеток и артефакты (фрагменты пластика, волокна, и т.п.). После этого маркированные чашки Петри помещали на рабочую площадку шейкера типа
ES-X (Khner AG, Швейцария), помещенную на дно СО2-инкубатора типа BBD6620 (Heraeus, Германия), с выносом управляющего (контрольного) блока шейкера на крышку инкубатора через пуповину, проведенную через технологическое отверстие на задней панели инкубатора. Значение скорости работы шейкера устанавливали 60 об/мин, с постепенным разгоном и торможением. После 2 суток инкубации, сформировавшиеся аггрегаты клеток (трансплантационные единицы) переносили в чашки Петри диаметром 20 мм с низкой адгезивностью типа NUNC (Дания) или Ultra Low Attachment (Corning, США).
Для анализа функциональности дофаминергических нейронов-производных чЭСК использовали технологию высокопроизводительной жидкостной хроматографии (HPLC): при этом применяли прибор ESA Coulochem III (ESA Biosciences Inc., США). Клетки растворяли в хлорной кислоте с использованием ультразвуковой обработки (соникации). Супернатант собирали, фильтровали, и использовали для электрохимической детекции дофамина и метаболитов дофамина. Подвижная фаза (смесь ацетата натрия
в концентрации 5 г/л, октансульфоновой кислоты 100 мг/л, ЭДТА 30 мг/л и 12% метанола; рН = 4,2) добавлялась с относительной скоростью 500 л/мин в колонку для обращенной хроматографии типа С18 (Chrompack/Varian, США). Анализ интенсивности пиков, соответствующих дофамину и 3,4-дигидрофенилуксусной кислоте (DOPAC) проводили
с использованием программного пакета Azur Chromatographic (DATALYS, Theix, Франция).
Для проведения иммуноцитохимического (ИЦХ) анализа, клетки фиксировали холодным 4% раствором параформальдегида (ПФА). Для ИЦХ анализа образцов «трансплантационных единиц» (ТЕ), их предварительно насыщали 30% раствором сахарозы и после фиксации замораживали в лунках пластиковой формы типа CryoMold (Sakura Finetek; Япония) с реактивом Tissue-Tek О.С.Т. (Sakura Finetek; Япония). Для проведения иммуногистоохимического (ИГХ) анализа тканей экспериментальных животных, крысы умерщвлялись введением 0,4 мл фенобарбитала интраперитонеально; используя перистальтический насос модели SciQ 323 (Watson Marlow Bredel, США) осуществляли перфузию большого круга кровообращения холодным 4% раствором ПФА (использовали минимум 300 мл ПФА). Головной мозг насыщали 30% водным раствором сахарозы с 0,5% азида натрия (NaN3). «Трансплантационные единицы» или образцы головного мозга экспериментальных животных помещали на площадку микротома поверх капли реагента O.C.T. Tissue-Tek (Sakura, Япония), и позиционировались/ориентировались по трём осям. Площадку микротома обкладывали сухим льдом, и образец примораживали; в ходе работы добавляли сухой лёд по мере необходимости. ТЕ нарезали на слои толщиной 6-8 мкм, образцы головного мозга - на слои толщиной 40 мкм. Слои последовательно раскладывали
в 10 маркированных виалов.
Гистологические образцы окрашивали по DAB с использованием системы АВС
(ABC Staining System; Santa Cruz Biotechnology, США), либо стандартным методом. Фиксированные образцы пермиабилизировали 0,1% раствором Тритон Х-100, инкубировали с первичными антителами при комнатной температуре в течение 2 часов, в темноте, затем инкубировали с вторичными антителами в течение 2 часов при комнатной температуре,
в темноте. При необходимости, после промывок образцы докрашивали 4',6-диамидино-
2-фенилиндолом (разведение до 300 нг/мл в 1% сыворотке).
Для создания экспериментальной модели локализованной, ассоциированной
с возрастном нейродегенеративной патологии в интересах отработки хирургической техники и изучения возможностей клеточной терапии, была использована технология, основанная на токсическом воздействии 6-гидроксидофамина (6-OHDA)
на дофаминергические нейроны. В работе была использованы 121 половозрелая самка крыс линии Sprague-Dawley, полученных из питомника B&K Universal Ltd. (г. Соллентуна, Швеция) весом 180-220 г. За 10 мин до начала хирургической операции, крыс анастезировали интраперитонеальным введением фентанила и домитора (0,3 мг/кг живого веса и 6,3 мл/кг живого веса, соответственно). Анестезированных и маркированных крыс укладывали на рабочую площадку системы (рамки) стререотаксической хирургии
(David Kopf Instruments, США). Крыс фиксировали в системе последовательным введением
в оба ушных отверстия тупоконечных стержней (с контролем возможности движения
в единственной оси) и установкой ограничителя (ТВ) с зацепом за передние резцы. Увеличение обеспечивалось применением систем хирургической стереомикроскопии
SMZ-168 и SMZ-645 (Nikon, Германия). Координаты брегмы регистрировали в карте операции. Расчеты координат будущих инъекций токсина по продольной и сагиттальной осям проводили из расчета: Инъекция 1: сагиттальная ось: -4,4 мм от брегмы; продольная ось: -1,2 мм от брегмы (при установке ограничителя (ТВ, от “tooth bar”) -2,3 мм от нуля). Инъекция 2: сагиттальная ось: -4,0 мм от брегмы; продольная ось: -0,8 мм от брегмы
(при установке ограничителя (ТВ) +3,4 мм от нуля). При этом перемещение расчетной точки производили вперёд по сагиттальной оси и вправо по продольной оси. Гамильтоновский шприц объемом 10 мкл устанавливали в рамку стререотаксической хирургии в строго вертикальное положение (контроль осуществляли по стеклянной призме). Иглу трижды промывали ледяным 0,02% раствором аскорбиновой кислоты (Sigma, США) в физрастворе. Раствор токсина готовили непосредственно пред началом серии инъекций, с соблюдением мер предосторожности. Инъекции токсина осуществляли по следующим суммарным координатам: Инъекция 1: 2,5 мкл токсина; сагиттальная ось: -4,4 мм от брегмы; продольная ось: -1,2 мм от брегмы; фронтальная ось: -7,8 мм от твердой мозговой оболочки
(при установке ограничителя (ТВ) -2,3 мм от нуля). Инъекция 2: 2,0 мкл токсина; сагиттальная ось: -4,0 мм от брегмы; продольная ось: -0,8 мм от брегмы; фронтальная ось:
-8,0 мм от твердой мозговой оболочки (при установке ограничителя (ТВ) +3,4 мм от нуля). При этом перемещение расчетной точки производили вперёд по сагиттальной оси, вправо
по продольной оси, и вентрально по фронтальной оси. После проникновения под твёрдую мозговую оболочку, иглу погружали на заданную глубину. Инъекцию 1 токсина проводили со скоростью 0,5 мкл за 30 сек (не постоянно, а с интервалами длительностью 30 сек). После окончания инъекций, продолжали оказывать давление на поршень шприца в течение 5 мин, затем (не отпуская поршень) иглу медленно извлекали, используя соответствующий винт собственно стереотакса. После завершения Инъекции 2, крысам делали подкожные инъекции темгезика (разведение физраствором 1:10) в дозировке 1,7 мл разведения на кг веса животного и подожную инъекцию 2 мл физраствора. После 5 минут вне рамки, крысам делали подкожные инъекции антидота (разведение антиседана в дистиллированной воде 1:5) в дозировке 1 мл разведения на кг веса животного, затем вновь подожную инъекцию 2 мл физраствора. Частичное пробуждение наступало приблизительно через 6 мин после введения антидота.
В опытах по трансплантации клеточной суспензии (производные чЭСК) животным модельным в отношении локализованной ассоциированной с возрастом нейродегенеративной патологии, после завершения дифференцировки чЭСК по протоколу SDIA (дни 16, 20 и 23 протокола дифференцировки) производили отделение колоний производных чЭСК от подложки (стромальных клеток) при помощи обработки папаином (Worthington Biochemical Corporation, США), с последующей диссоциацией клеток при помощи аккутазы (Innovative Cell Technologies, США), как описано выше. Диссоциированные клетки исследовали в отношении жизнеспособности при помощи окраски трипановым синим (TrBlue, Sigma, США), после чего разводили в сбалансированном солевом растворе (солевом буфере) Хэнкса (HBSS) с добавлением 0,05% деоксирибонуклеазы I (ДНКазы I; Sigma, США) до концентрации 50 000 жизнеспособных кл/мкл. Используя систему для стереотаксической хирургии, модельным животным (крысы, получившие интрастриатальные инъекции 6-гидроксидофамина (6-OHDA)) вводили 100 000 жизнеспособных клеток (2 мкл клеточной суспензии). Инъекции проводили
с использованием техники, полностью совпадающей с описанной выше. Инъекцию клеточной суспензии осуществляли по следующим суммарным координатам: сагиттальная ось: +1,0 мм от брегмы; продольная ось: -3,0 мм от брегмы; фронтальная ось: -5,0 мм
от твердой мозговой оболочки. Клеточный материал хранили на льду, клетки использовали для трансплантаций в течение максимум 4 часов после отделения от подложки. Скорость инъекции составляла 0,5 мкл клеточной суспензии за 30 сек (не постоянно, а с интервалами длительностью 30 сек). После окончания инъекций, продолжали оказывать давление на поршень шприца в течение 5 мин, затем (не отпуская поршень) иглу медленно извлекали, используя соответствующий винт собственно стереотакса. Клеточный материал,
не использованный для трансплантации модельным животным, применяли для засева индивидуальных лунок готовых 8-луночных «рамочных» плашек, покрытых смесью полиорнитина и ламинина (Becton, Dickinson, США), - по 14 000 живых клеток на лунку. Cпустя 24 часа плашки фиксировали, и изучали для уточнения клеточного состава пула использованных для трансплантации клеток.
Трансплантация «трансплантационных единиц» животным модельным
в отношении локализованной ассоциированной с возрастом нейродегенеративной патологии проводилась с использованием техники стереотаксической хирургии, аналогичной описанной выше, однако с рядом важных изменений и дополнений. Перед началом трансплантации, «трансплантационные единицы» под контролем микроскопа пипеткой переносили из суспенизонной культуры в стерильную пробирку с небольшим объемом питательной среды. При этом проводилась селекция «трансплантационных единиц», не содержащих посторонних включений (обычно ворсинок или фрагментов пластика), имеющих правильную форму (шаровидную или округлую, вытянутую по одной из осей). Несколько (2-3) репрезентативных «трансплантационных единиц» диссоциировали аккутазой для измерения числа содержащихся в них клеток. Концентрацию суспензии диссоциированных клеток измеряли в счетной камере и нормализовали по числу диссоциированных «трансплантационных единиц». Все остальные, предназначенные для трансплантации «трансплантационные единицы» переносили в солевой буфер (сбалансированный солевой раствор) Хэнкса (HBSS) с добавлением
0,05% деоксирибонуклеазы I (ДНКазы I; Sigma, США) и пробирку помещали на лёд. Собственно трансплантацию производили в течение 40 минут - 2 часов после извлечения
из СО2-инкубатора.
Для трансплантации «трансплантационных единиц» в структуры головного мозга животных модельных в отношении локализованной, ассоциированной с возрастом нейродегенеративной патологии использовали специальные иглы для Гамильтоновских шприцев объемом 10 мкл. Использовали остроконечные иглы размера 22G (Gauge) и 23G (Gauge). При этом наружные диаметры игл составляли 0,72 и 0,64 мм, соответственно; внутренние диаметры игл составляли 0,41 и 0,34 мм, соответственно; толщина стенок игл
в обоих случаях составляла 0,15 мм. Загрузка «трансплантационных единиц»
в Гамильтоновский шприц производилась под контролем световой микроскопии, на малом увеличении (х 3,5) микроскопа модели CK30 (Olympus, Япония). После подготовки животного к трансплантации и выверки координат стереотаксической хирургии, как описано выше, шприц вынимали из зажимов. На крышку стерильной чашки Петри наносили
2 отдельно лежащих капли солевого буфера Хэнкса (HBSS). Необходимое количество «трансплантационных единиц» (соответствующее 100 000 живых клеток) переносили из пробирки микропипеткой в одну из капель, затем, под контролем микроскопа - в другую. Для предупреждения выпадения «трансплантационных единиц» из шприца в ходе манипуляций, иглы Гамильтоновских шприцев после промывок загружали следующими слоями (считая от поршня к острию иглы): физраствор (целиком заполняет мертвое пространство иглы), «воздушный замок» (объем 0,3 мкл), чистый солевой буфер Хэнкса (HBSS; объем 1,5 мкл), солевой буфер Хэнкса (HBSS) с «трансплантационными единицами» (объем 2,5 мкл), «воздушный замок» (объем 0,3 мкл). Скорость инъекции составляла 0,5 мкл общего объема содержимого шприца за 30 сек (не постоянно, а с интервалами длительностью 30 сек). После окончания инъекций, продолжали оказывать давление на поршень шприца в течение 5 мин, затем (не отпуская поршень) иглу медленно извлекали, используя соответствующий винт собственно стереотакса. Эффективность трансплантации «трансплантационных единиц» контролировалась промывкой использованного для инъекций Гамильтоновского шприца физраствором, с изучением лаважа под световым микроскопом.
Для предупреждения отторжения ксеногенного для модельных животных клеточного материала, проводилась агрессивная иммуносупрессорная терапия. За 1 сутки
до проведения трансплантационной процедуры, крысам делали интраперитонеальную инъекцию циклоспорина А (Сандимунн, Novartis, Швейцария; разведение в физрастворе до 100 мг/мл) в дозировке 15 мг/кг живого веса. Также за 1 сутки до проведения трансплантационной процедуры, крысам делали интраперитонеальную инъекцию бетапреда (Бетаметазон, Glaxo-Wellcome, Швеция) в дозировке 0,12 мг/живого веса.
В послеоперационном периоде животные содержались в полипропиленовых клетках, помещенных в стерильный кондиционируемый шкаф типа Scantainer (Scanbur Ltd., Дания). Инъекции циклоспорина А производили ежедневно в течение всего эксперимента, однако начиная с 15 дня после трансплантационной процедуры дозировку уменьшали до 10 мг/кг живого веса. Инъекции бетапреда производили ежедневно в течение 10 дней после трансплантационной процедуры, после чего прекращали.
Анализ моторных параметров (моторных функций) животных модельных
в отношении локализованной ассоциированной с возрастом нейродегенеративной патологии до и после трансплантации клеточного материала был основан на исследовании ротационного поведения со стимуляцией амфетамином. При использовании этого метода, значимые изменения ротационного поведения наблюдаются при потере 40-50% секреции дофамина в полосатом ядре (что соответствует потере >50% дофаминергических нейронов
в substantia nigra pars compacta), при этом использование модели, основанной на «полном» токсическом поражении дофаминергических нейронов 6-гидроксидофамином («complete lesions») не сопровождается спонтанным и быстрым восстановлением моторных функций [128]. 3-4 недели спустя после введения модельным животным 6-гидроксидофамина, либо после трансплантации клеточного материала, с промежутками по 2-3 недели, крысам интроперитонеально вводили амфетамин из расчета 2,5 мг/кг веса, разведенный
в стерильном физрастворе. Немедленно после этого крыс помещали в чашки системы ротометрии, и фиксировали защащенной пласиковым покрытием петлёй поперек тела. Мониторинг поворотов (ротометрия) производили с использованием автоматизированной системы Rotomax Analyzer (AccuScan Instruments Inc., США), при этом запись показателей ротометра производился с использованием программного обеспечения RotoMax v.1.40 (AccuScan Instruments Inc., США), а форматирование показателей и их импорт в формат MS Excell - с использованием программного обеспечения Rotolyze v.1.20 (также AccuScan Instruments Inc., США). В настройках программного обеспечения задавали:
1) идентификацию поворотов животного на каждые оборота (т.е. на 90о; регистрировалось как 0,25 оборота) и 2) проведение анализа промежутками по 5 минут, то есть минуты 0-5,
5-10, 10-15 анализа, и т.д. Общая длительность исследования составляла 90 минут. Одновременно могло исследоваться до 8 крыс. Исследование проводили в отдельном помещении с постоянным освещением; в ходе исследования минимизировали перемещения людей поблизости от испытуемых животных. Значение моторной функции животных (ротационный показатель) рассчитывали как отношение общего числа полных правых минус полных левых поворотов к длительности исследования в минутах.
В опытах по изучению транскриптома стволовых клеток разных линий, тотальную РНК выделяли из клеток с использование систем RNase AWAY (QIAGEN, США). мРНК выделяли из тотальной РНК олиго(dT)-целлюлозой системой Messagemaker (Gibco BRL, США). кДНК синтезировали из 5 мкг мРНК с использованием системы cDNA Synthesis kit (Gibco BRL, США), с мечением продукта радиоактивным изотопом [32P] (1 мккюри) (Amersham, США).
Создание SAGE-библиотек осуществлялось на основе протокола 1.0с серийного анализа генетической экспрессии (SAGE) с многочисленными модификациями, в том числе собственными. При этом последовательно проводили создание ярлыков кДНК; создание, амплификацию, и очистку спаренных ярлыков кДНК; конкатемеризацию и клонирование спаренных ярлыков кДНК; контроль качества созданных SAGE-библиотек и их секвенирование; создание и пре-процессинг SAGE-каталогов; и процессинг SAGE-каталогов. В ходе работы, ярлыки кДНК создавали с использованием рестрикционных ферментов NlaIII (якорный фермент) и BsmFI (ярлыковый фермент) (New England Biolabs, США). Конкатемеры ярлыков кДНК клонировали в клетки E.coli ElectroMAX DH10B (Gibco BRL, США) при помощи плазмидного вектора pZeRO-1 (Invitrogen, США) и электропорационного аппарата Gene-Pulser II (Bio-Rad, США). Селекцию бактериальных клонов производили на низкосолевом LB-агаре с зеоцином (50 мкг/мл). Качество клонирования контролировали при помощи полимеразной цепной реакции (ПЦР) и структурирование конкатемеров проверяли определением их нуклеотидной последовательности (секвенированием) на полуавтоматическом секвенаторе MBI310 Prizm (PE Applied Byosistems, США). Бактериальные клоны выращивали в 96-луночных плашках с питательным бульоном ТВ/зеоцин, крупномасштабное секвенирование производилось фирмой Celera Genomics/PE Applied Byosistems (США). Ярлыки кДНК идентифицировали при помощи программного пакета SAGE 2000 Version 4.12, идентификация ярлыков кДНК производилась с использованием баз данных Национального Центра биотехнологической информации США.
Для работы по анализу характеристик транскриптома стволовых клеток и их производных, в том числе в динамике, была создана собственная высокоспециализированная микрочиповая платформа «NeuroStem». Основными группами генов мишеней платформы являются: I. Гены «стволовости» (установленные/общепринятые маркеры, кандидатные маркеры, вторичные гены-мишени, биологически связанные с генами «стволовости»); II. Гены нейральных и нейрональных клеток разных типов (в том числе пан-нейральные маркеры, имеющие разный уровень специфичности маркеры клеток нейрального/нейронального рядов, маркеры роста таких клеток, их окончательной дифференцировки, созревания (матурации), миграции, миелинизации, разветвления (браншинга) аксонов нейронов, образования синапсов, и т.п., гены вовлечённые в патогенез различных нейродегенеративных заболеваний); III. Гены коммитирования/дифференцировки стволовых клеток в клетки нейрального/нейронального рядов (маркеры роста и деления клеток, дифференцировки, регенерации, нейральной/нейрональной дифференцировки, и т.п.); дополнителельно (без выделения в собственные группы) были использованы гены, соответствующие многочисленным типам контролей: а) положительных и б) отрицательных), в том числе гены, обладающие относительно высоким уровнем экспрессии в клетках любых типов (так называемые «гены домашнего хозяйства»), гены, соответствующие маркерам отдельных типов клеток,
не являющихся нейронами и глиальными клетками, и т.д., в) гены, относящиеся к системе апоптоза (программируемой клеточной гибели), г) так называемые «часовые гены», имеющие отношение к синхронизации биологических ритмов, д) гены, относящиеся
к антиоксидантной системе, е) импринтинговые гены, ж) гены (члены семейств генов), вовлеченные в сигнальные системы FZD, WNT, BMP, STAT, FGF, з) гены систем каспаз и Kruppel-гены, и) гены, относящиеся к теломерной/теломеразной системе, й) маркеры раковых клеток.
Длинные олигонуклеотидные зонды (длина = 69-71 нуклеотиду) соответствующие избранным генным мишеням были избраны из баз зондов человека AROS версий V2
(22272 вводных) и V3 (5520 вводных) (Operon Biotechnologies Inc., США). Печать микрочиповых платформ осуществлялась компанией SweGene (Швеция), являющейся автономным подразделением/дочерним предприятием Люндского университета (Швеция) (SweGene DNA Microarray Resource Centre, Department of Oncology at Lund University). Печать производилась роботизированой системой (споттером) типа MicroGrid II 600R arrayer,
с наконечниками (печатающими головками) типа MicroSpot 10 K (Harvard BioRobotics, США). Платформы печатали на стеклянных пластинках (слайдах) формата предметных стёкол, типа UltraGAPS aminosilane slides (Corning Inc., США). Плотность печати составляла 140 мкм между центрами точек нанесения генетического материала (зондов) и 90-110 мкм диаметр точек нанесения генетического материала. Непосредственно после печати готовые платформы маркировались наклейками с уникальными штрих-кодами, сушились
и хранились до момента использования в закрытых непрозрачных пластиковых контейнерах в вакууме. Срок хранения готовых микрочиповых платформ не превышал 4 месяцев после печати.
Тестирование готовых микрочиповых платформ (тестирование качества печати) осуществлялось визуальным обследованием (микроскопией) и пробной (тестовой) гибридизацией с образцом РНК гарантированно имеющим высокий уровень экспрессии широкого спектра генов разных семейств, - представляющим собой готовую смесь РНК человека, Human Universal Reference RNA (Stratagene, США). Оценивалась точность печати как в отношении правильности размеров и однородности размеров точек нанесения генетического материала (зондов), так и точности нанесения точек отосительно друг друга в пределах одного блока, и отдельных блоков отосительно друг друга в пределах одной платформы (слайда), а также положение всей зоны печати в пределах слайда и ориентировка блоков (по экспрессии -актина, уникальная точка нанесения в левом верхнем углу каждого из блоков) и зоны печати целиком, отсутствие артефактов (ворсин, царапин, и т.п.).
Для применения микрочиповых платформ «NeuroStem» в эксперименте по изучению транскриптома стволовых клеток и их производных, последовательно проводили синтез кРНК-зондов, меченных флуоресцентными красителями; прегибридизацию; гибридизацию; постгибридизационные отмывки; сканирование и трансформацию данных; анализ данных. Синтез кРНК-зондов с интегрированным этапом мечения флуоресцентными красителями проводили с использованием системы Low RNA Input Fluorescent Linear Amplification Kit (Agilent Technologies, США). Предварительно тестированные образцы высококачественной тотальной РНК (50-500 нг) разводили или концентрировали (с использованием вакуумного концентратора SpeedVac; Thermo Scientific, США)). В качестве контроля использовали готовую смесь РНК человека (400 нг) Human Universal Reference RNA (Stratagene, США).
В каждую пробирку с образцом РНК добавляли смесь, содержащую обратную транскриптазу MMLV (RT MMLV), а на следующем этапе - флуоресцентный краситель цианин-3-ЦТФ (Су3) либо цианин-5-ЦТФ (Су5) (Perkin-Elmer, США). Для синтеза кРНК, в пробирки
с продуктами первой реакции добавляли смесь, содержащую неорганическую пирофосфатазу и Т7 РНК-полимеразу. Продукты второй реакции экстрагировали с применением системы RNeasy mini (QIAGEN, США). Качество образца оценивали по флуоресценции Су3 или Су5
с помощью спектрофотометрии на спектрофотометре ND-1000 (Nano-Drop, США). Стеклянные слайды системы “NeuroStem” облучали УФО в ДНК-кросслинкере Biolink (BioMetra, США) с установкой уровня энергии 800 мДж/см2. Гибридизацию проводили
с использованием системы MAUI в рабочем режиме гибридизации «А», при этом применяли сборные камеры типа 1 MAUI Mixer AO plate (BioMicro Systems, США). Гибридизацию флуоресцентных проб с микрочиповыми платформами (слайдами) проводили при 42оС
в течение 17 часов. После постгибридизационных отмывок, слайды помещали
(с использованием адаптеров) в гнёзда карусели роботизированного сканера высокого разрешения типа DNA Microarray Scanner G2565CA (Agilent Technologies, США). Ориентация штрих-кодов соответствовала требованиям сканера штрих-кодов. Запись данных (с инкорпорацией штрих-кода, считанного с соответствующей позицией на карусели сканера) осуществлялась с использованием программного пакета DNA Analytics Software (Agilent Technologies, США), немедленный контроль качества сканирования (в частности, полноты сканирования) - с использованием программного пакета GenePix Pro (Axon Instruments, США). Изображения форматировали (переворачивали и разводили на «зелёное» и «красное») с использованием программного пакета Tiff Image Channel Splitter Utility (Agilent Technologies, США). После форматирования изображения производили приложение координатной сетки, с адаптацией её положения вручную для слайда в целом, и для каждого блока в отдельности.
Для каждого слайда затем проводился контроль отсутствия артефактов для каждой точки нанесения материала в пределах каждого блока. Под артефактами понимались следующие варианты: 1) кристаллы флуоресцентных красителей, совпадающие по координатам с индивидуальными точками нанесения материала, 2) ворсинки или 3) царапины, совпадающие по координатам с индивидуальными точками нанесения материала (или несколькими точками сразу). В каждом таком случае точка нанесения материала маркировалась «флагом», отрицающим её использование для анализа. Одновременно проводилась уточнение размеров автоматически наложенной на координатную сетку разметки сбора сигнала в соответствие с интенсивностью сигналов для индивидуальных точек. В отдельных случаях, особыми «флагами» маркировались точки без сигнала, что отрицало их использование для анализа, но не для расчета интенсивности фонового сигнала. Данные трансформировали из изображений в цифровую форму (интенсивность сигнала, ассоциированная с конкретной точкой нанесения генетического материала (зонда), обладающей уникальным ярлыком (именем) с использованием программного пакета GenePix Pro (Axon Instruments, США); использовалась функция сокращения («вычитания») интенсивности фонового сигнала, рассчитываемая индивидуально для каждого гибридизованного слайда.
Нормализацию данных проводили индивидуально для каждого блока микрочиповой платформы с использованием алгоритма LOWESS со значением фактора сглаживания принятым равным 0,33. Сведение данных от технических репликатов и анализ данных проводили с использованием базы данных и комплекса аналитического программного обеспечения BioArray Software Environment database (BASE; http://base.thep.lu.se/, Люндский университет, Швеция). Визуализацию графиков вариации интенсивности сигнала осуществляли с использованием программы TIGR MultiExperiment Viewer (MEV; http://www.tm4.org/mev/; http://www.tigr.org) из состава программного пакета TM4 Microarray Software Suite (Dana-Farber Cancer Institute, США).
Полимеразная цепная реакция с обратной транскрипцией (ПЦР-ОТ) производилась на программируемом амплификаторе типов GeneAmp PCR System 9600 (Perkin-Elmer, США) или MiniOpticon (Bio-Rad, США). Праймеры для ПЦР-ОТ были избраны из опубликованных ранее работ или разработаны с использованием программного обеспечения Oligo 4.0 (Molecular Biology Insight, США), Primer3 или Clone Manager Suite 7.1 (Sci Ed Software, США). Использовали Tth полимераза (Promega, США), либо полимеразу REDTaq (Sigma-Aldrich, США). Переменные (число циклов амплификации, температура отжига) отрабатывались на этапе отработки оптимальной технологии ПЦР-ОТ, в том числе
с использованием функции «градиента температуры» программируемого амплификатора MiniOpticon (Bio-Rad, США). Продукты ПЦР-ОТ анализировали электрофорезом
в 1% агарозном геле, с визуализацией окрашиванием бромидом этидия. Интенсивность сигналов продуктов ПЦР-ОТ анализировалась при помощи программного пакета Multi-Analyst 1.1 (Bio-Rad, США).
Для статистической обработки результатов исследования использовали современные математические методы анализа полученных данных, включая однофакторный дисперсионный анализ и критерий Шеффе (расчет влияния времени пребывания в культуре in vitro на выход дофаминергических нейронов), дисперсионный анализ Краскала-Уоллеса (анализ результатов трансплантации: выживаемость, трансплантация, дифференцировка), критерий U Манна-Уитни (попарное сравнение между группами в один момент времени, либо между двумя временными точками в пределах одной группы), t-критерий Стьюдента (анализ воздействия биологических факторов на клеточные культуры in vitro), локально взвешенную регрессию (LOWESS curve fitting; нормализация данных гибридизации образцов с микрочиповыми платформами). Сравнительный анализ распределения частот встречаемости SAGE-ярлыков в индивидуальных SAGE-библиотеках производили
с использованием метода Madden. В работе использовали программы статистической обработки данных MS EXCEL, StatView 5.0 и EPCLUST.
Результаты исследований и их обсуждение
В соответствии с задачами исследования, направленного на изучение механизмов пролиферации и дифференцировки стволовых клеток, и разработки способа заместительной клеточной терапии локализованной, ассоциированной с возрастом нейродегенеративной патологии в условиях эксперимента, в рамках проведенной работы последовательно выполнялись эксперименты по изучению транскриптома стволовых клеток разных типов, в том числе в ходе дифференцировки; отработки протоколов размножения стволовых клеток in vitro, их направленной дифференцировки в функциональные дофаминергические нейроны; клеточной терапии производными стволовых клеток с использованием лабораторной модели локализованного возрастного нейродегенеративного процесса, и анализ результатов трансплантации клеточного материала.
Экспансия стволовых клеток разных типов in vitro
С целью получения значительного объема клеточного материала для применения в протоколах экспериментальных исследований, была успешно отработана технология размножения (экспансии) эмбриональных стволовых клеток человека in vitro, после чего проведен сравнительный анализ протоколов. Методика культивирования экспансии фидерных (питающих) клеток не вызвала больших затруднений: в ходе работы, в течение нескольких пассажей (мЭФ: пассажи 0-2; неонатальные фибробласты крайней плоти человека: пассажи 4-7) удавалось наработать значительный объем клеточного материала. Клетки данных типов хорошо переносили процедуры заморозки и размораживания, и продолжали активно пролиферировать на дочерних плашках. Отработанные протоколы антимитотической обработки фидерных клеток (при помощи митомицина С или гамма-радиации) обеспечили весьма эффективное подавление пролиферативной активности этих клеток, что позволило с успехом применять их в сложных клеточных системах (в формате ко-культивирования с ЭСК). В ходе работы была выявлена способность чЭСК линии SA002 к росту на фидерных (питающих) клетках человеческого происхождения без каких-либо иных адаптаций протокола. В каждом случае, размораживание чЭСК из соломинок проходило с использованием мЭФ, и после 2 пассажей на мЭФ, чЭСК в ходе очередного пассажа переносили в «дочерние» плашки с неонатальными фибробластами крайней плоти человека (чФ), после чего культивировали в стандартных условиях. Такая возможность выгодно отличала чЭСК линии SA002, что позволило использовать ее в экспериментах по разработке способа заместительной клеточной терапии локализованной, ассоциированной с возрастом нейродегенеративной патологии.
Успех культивирования чЭСК линий #237 и SA002 определялся следующими факторами: 1) использованием для хранения чЭСК протокола витрификации (остекленения); 2) применением фидерных клеток на низких пассажах; 3) культивированием чЭСК в желатинизированных плашках для искусственного оплодотворения (IVF-плашках); 4) высокой лабораторной культурой, в частности крайне жестким температурным и газовым регламентом. Использование весьма сложной технологии механических пассажей и строгое соблюдение перечисленных выше правил позволило добиться эффективной экспансии чЭСК in vitro, с наработкой значительного объема клеточного материала для проведения последующих экспериментов. Совокупность данных по экспансии эмбриональных стволовых клеток человека разных линий in vitro позволила заключить, что с точки зрения удобства и эффективности протоколов заморозки и размораживания, предпочтительной является работа с чЭСК линий SA002 и HUES-3; с точки зрения возможности получения значительного объема клеточного материала (для применения в протоколах экспериментальных исследований) - линий #237 и SA002; с точки зрения удобства пассажей - работа с чЭСК линии HUES-3; и с точки зрения получения на выходе пассажа популяции клеток в малой степени контаминированной фидерными клетками (в том числе ксеногенного происхождения) - линии SA002. Таким образом, чЭСК линии SA002 были признаны в наиболее полной степени отвечающими выдвинутым требованиям по возможности использования в эксперименте, и именно эта линия чЭСК стала объектом экспериментальной работы по дифференцировке в дофаминергические нейроны и субстратом клеточной терапии локализованной, ассоциированной с возрастом нейродегенеративной патологии в экспериментальной модели.
Идентификация ключевых факторов контроля пролиферации и дифференцировки стволовых клеток
С целью идентификации генов, отвечающих за поддержание стволовых клеток разных типов в недифференцированном состоянии в ходе культивирования in vitro, а также генов вовлеченных в процессы дифференцировки стволовых клеток, с использованием метода серийного анализа генетической экспрессии (SAGE; Velculescu V., et al., 1995) были созданы следующие SAGE-библиотеки, то есть библиотеки так называемых «ярлыков» транскриптов:
1) SAGE-библиотека недифференцированных эмбриональных стволовых клеток мыши линии R1;
2) SAGE-библиотека недифференцированных эмбриональных стволовых клеток мыши линии D3;
3) SAGE-библиотека недифференцированных стволовых клеток мыши линии EG-1, производных первичных половых клеток;
4) SAGE-библиотека недифференцированных клеток эмбриональной карциномы мыши линии Р19;
5) SAGE-библиотека клеток эмбриональной карциномы мыши линии Р19, дифференцируемых в кардиомиоциты, на раннем этапе (день 3 + 0,5 протокола in vitro дифференцировки);
6) SAGE-библиотека клеток эмбриональной карциномы мыши линии Р19, дифференцируемых в кардиомиоциты, на среднем этапе (день 3 + 3,0 протокола in vitro дифференцировки);
7) Эмбриональные фибробласты мыши, пассаж 3 (P3).
SAGE-библиотеки создавалась с использованием оригинального протокола серийного анализа генетической экспрессии (SAGE), с широким рядом модификаций, в том числе собственных. В результате определения нуклеотидных последовательностей клонов индивидуальных SAGE-библиотек были созданы SAGE-каталоги общим объёмом свыше 460 тысяч ярлыков кДНК. В ходе препроцессинга они были последовательно очищены от дупликативных спаренных ярлыков, артефактов клонирования и сиквенирования (для чего была разработана и использована собственная математическая модель), неинформативных ярлыков-копий используемых линкеров и поли(А) ярлыков. Таким образом, в ходе работы были созданы имеющие высокую степень надежности SAGE-каталоги, анализ которых позволил получить важные данные о ключевых генах, экспрессирующихся в активно пролиферирующих недифференцированных стволовых клетках разных типов (гены «стволовости») и в ходе ранних этапов дифференцировки, - а также транскриптоме фидерных клеток. Все SAGE-каталоги были размещены в общедоступной базе данных Gene Expression Omnibus (GEO) Национального центра биотехнологической информации США (NCBI) для дальнейшего использования научным сообществом под следующими индексами: R1 ES cells, GSM580; D3 ES Cells, GSM11349; EG-1 Cells, GSM11348; P19 EC cells, GSM1682; P19 EC cells, differentiation Day 3+0.5, GSM1683; P19 EC cells, differentiation Day 3+3.0, GSM1684; Pimary mouse embryonic fibroblasts, GSM7759. Сравнительный анализ групп SAGE-каталогов позволил выявить следующие закономерности:
а) Большинство транскриптов клеток разных типов имеет низкий уровень экспрессии. Состав транскриптов с высоким уровнем экспрессии (копийности SAGE-ярлыков) определяется функциональными свойствами конкретного типа клеток в данных условиях.
б) В клетках разных типов, наиболее высоко экспрессированные транскрипты (относящиеся как к ядерному, так и к митохондриальному геномам) соответствуют продуктам, вовлеченным в синтез белков, энергетический метаболизм, цитоплазматическим и мембранным белкам, белкам цитоскелета, ферментам, и секреторным белкам.
в) Подавляющее большинство транскриптов имеют равный уровень экспрессии вне зависимости от уровня дифференцировки и её направления.
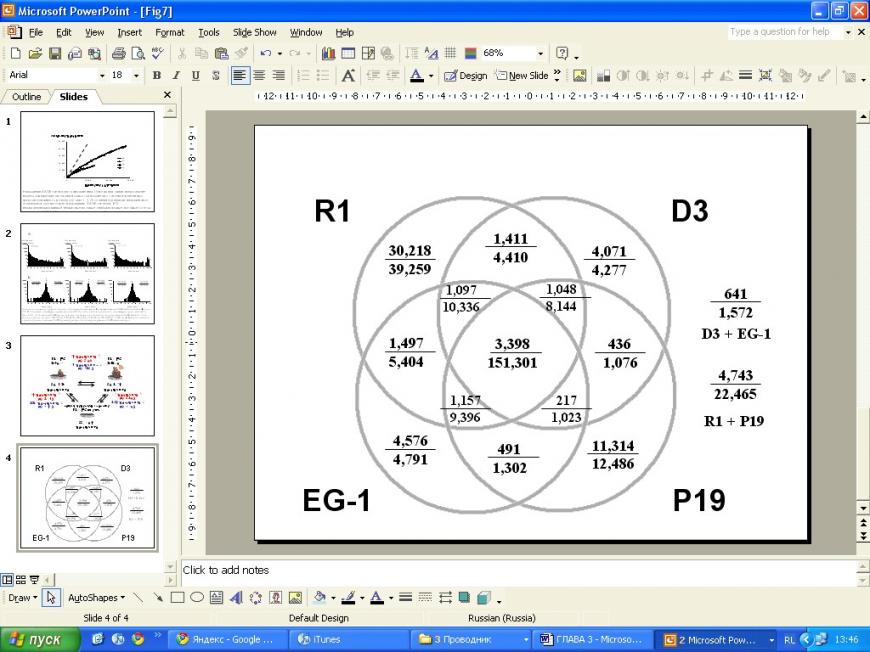
В ходе сравнительного анализа транскриптома недифференцированных стволовых клеток разных типов (линии R1, D3, EG-1, и Р19) было проанализировано распределение 277 242 надежных (т.е. очищенных от неинформативных ярлыков и артефактов) SAGE-ярлыков, соответствующих 66 315 уникальным транскриптам (Рис. 1). Из них, 3 398 уникальных транскриптов (5,12% спектра всех уникальных транскриптов, идентифицированных в 4 SAGE-каталогах вместе взятых) были представлены во всех 4 SAGE-каталогах («фокус» диаграммы Венна). При этом число таких уникальных транскриптов представленных более чем 1 копией в по крайней мере 3 SAGE-каталогах составило 2 540 (3,83%), а представленных более чем 1 копией во всех 4 SAGE-каталогах - 1 752 (2,64%). На следующем этапе анализа, нормализованные значения уровней экспрессии этих транскриптов проверяли для доступных SAGE-каталогов, представляющих клетки иной природы (не стволовые). Проведенный анализ позволил идентифицировать группу из 238 уникальных транскриптов, представленных более чем 1 копией в по крайней мере 3 из 4 собственных SAGE-каталогов недифференцированных стволовых клеток и имеющих средний уровень экспрессии в стволовых клетках по крайней мере в 4 раза выше, чем в среднем для иных типов клеток.
Рис. 1. Качественный сравнительный анализа транскриптомов стволовых клеток мыши разных линий, представленный в виде единой Венновской диаграммы. В каждом сравнении, в числителе представлено число идентифицированных уникальных транскриптов, в знаменателе - общее число SAGE-ярлыков.
В ходе дальнейшей работы, «кандидатные» молекулярные мишени из состава групп, функция которых потенциально связана со «стволовостью» (то есть способностью стволовых клеток пролиферировать, сохраняя недифференцированное состояние) и ранними этапами дифференцировки были использованы в дизайне микрочиповой платформы “NeuroStem”, специализированной для исследования транскриптома стволовых клеток в динамике. В ходе создания микрочиповой платформы “NeuroStem” в интересах изучения профилей генной экспресии (в том числе динамических) стволовых, нейральных/нейрональных и переходных типов клеток была предпринята попытка создать универсальную платформу, пригодную для исследования не только чЭСК, но и других типов СК. В ходе дизайна микрочиповой платформы “NeuroStem” первого поколения (“NeuroStem” 1.0) основными группами генов-мишеней платформы стали: Установленные (например, Oct3/4, Tdgf1, Nanog) и кандидатные (например, Cpxm1, Pdcd11) генетические маркеры «стволовости» - 101; Гены, имеющие отношение к процессам дифференцировки, развития, пролиферации клеток в целом, в том числе установленные (например, Ssea3, Gata4, Wig1) и кандидатные (например, Nedd4, Lefty) маркеры этих процессов - 57; Гены, имеющие отношение к процессам нейральной/нейрональной дифференцировки (например, NeuroD1, Ash1, Pax2) - 81; Генетические маркеры нейронов и глиальных клеток разных типов (например, Tubb3, Gap43, Gfap) - 110; Маркеры иных типов клеток, в том числе клеток нормальных тканей (печени, поджелудочной железы, скелетной и гладкой сердечной мышцы, эпителия, эндотелия), субпопуляций клеток крови (например, CD4, CD14, CD19, CD33), раковых клеток разных типов (лимфома Ходжкина, клетки рака поджелудочной железы, молочной железы, мочевого пузыря, яичников) - 54; Особые группы генов (гены, относящиеся к системе апоптоза, антиоксидантной системе, теломерной/теломеразной системе, импринтинговые гены, члены семейств генов вовлеченных в сигнальные системы FZD, WNT, BMP, STAT, FGF, систем каспаз, и др.) - 308; Контроли (положительные и отрицательные). В общей сложности,
в состав платформы “NeuroStem” 1.0 вошло 1000 молекулярных мишеней из состава групп 1-7) и 18 500 иных генов (сигнал которых используется, в частности, для нормализации фоновых значений флуоресценции в пределах платформы) наносились на микрочиповую платформу в формате дупликатов (всего 39 000 точек нанесения генетического материала).
Микрочиповая платформы “NeuroStem” первого поколения (“NeuroStem” 1.0) была
с успехом использована для серии пилотных экспериментов, в ходе которых было получено подтверждение возможности её широкого использования в работе по изучению транскриптома клеток человека (в том числе нейронов, предшественников нейронов,
и стволовых клеток разных типов). На основании полученных данных было принято решение о модификации микрочиповой платформы. В ходе данного этапа работы
к исходному списку мишеней были добавлены дополнительные молекулярные мишени, ставшие известными за это время, либо признанные ценными в интересах выполнения последующей работы. Главным же изменением дизайна стало применения квадрипликатов для каждой молекулярной мишени или контроля (за исключением -актина, используемого как контроль ориентировки при печати платформ и представленного на каждом слайде
в 48 копиях). Микрочиповая платформа “NeuroStem” второго поколения (“NeuroStem” 2.0) содержала, в общей сложности, 1 312 молекулярных мишеней и 10 220 иных генов, которые наносились на микрочиповую платформу в формате квадрипликатов (всего 46 128 точек нанесения генетического материала). Платформа содержала 48 блоков, в пределах каждого из которых генетический материал наносился печатающей головкой на стеклянный слайд в виде 961 индивидуальных точек (31 х 31 точка); расстояние между центрами точек нанесения генетического материала составляло 140 мкм; средний размер точек составлял
90-110 мкм (Рис. 2). Одним из наиболее ценных параметров дизайна микрочиповой платформы “NeuroStem” стало наличие в её пределах 88 молекулярных мишеней, имеющих отношение к дофаминергической системе (маркеры дофаминергических нейронов, дофаминергических прогениторных клеток (клеток-предшественников), созревания дофаминергических нейронов, зрелых дофаминергических нейронов, маркеры незрелых (ранних) дофаминергических нейронов, и др.), в том числе идентифицированные в ходе собственных пилотных экспериментов (табл. 1).
Таблица 1
Представленные на микрочиповой платформе “NeuroStem” 2.0 молекулярные мишени, имеющие отнашение к дофаминергической системе
| Гены | Функция |
| Aadc, Ant2 (Slc25a5), Calb1, Dat (Slc6a3), Girk2 (Kcnj6), Igf1, Ptx3, Th | Маркеры дофаминергических нейронов |
| Dll1 1, En1, En2, Lmx1b 2, Pax5, Shh, Wnt1 | Маркеры дофаминергических прогениторных клеток (клеток-предшественников) |
| Adh2, Lhx1 (Lim1), Lhx5 (Lim2) | Маркеры созревания дофаминергических нейронов |
| Drd2 2, Vmat2 | Маркеры зрелых дофаминергических нейронов |
| Gfra1, Gfra2, Gfra3, Gfra4, Otx2 | Маркеры незрелых (ранних) дофаминергических нейронов |
| Dlx1, Dlx2, Flj38973, Lmx1a, Metrn 1, Mgc33365 1, Tafa4 (Fam19a4) 1 | Гены, имеющие отношение к процессам дифференцировки дофаминергических нейронов |
| Adcy7, Alcam, Art (Artn), Bcl11a, Calca, Cart, Cbln1, Cdh2, Col11a1, Cx43, D1lip (Drd1ip), Darpp32 (Ppp1r1b), Dbh, Doc2a, Drd1, Drd3, Drd4, Drd5, Drip78 (Dnajc14), Egln3 (Phd3), Egr1, Fos, Fxyd6, Galm, Ghr, Grin2c, Grp, Gsbs (C7orf16), Hs6st2, Igfbp4, Kcna5, Kcnab1, Math2, Mlp (Marcksl1), Moxd1, Mpp3, Nrip3, Nrp2, Nt (Nts), Ntn (Nrtn), Nurr1 (Nr4a2), Pac1 (Adcyap1r1), Pacap, Plagl1 (Zac), Pvrl3 (Nectin 3), Rab3c, Rcn1, Slc17a6 (Vglut2), Slc6a1, Sox6, Spp1, Tacr3, Trhr, Zfp161, Vav3, Zdhhc2 | Гены, имеющие отношение к дофаминергической системе |
1 Кандидатные молекулярные мишени, идентифицированные в ходе собственных пилотных экспериментов.
2 Могут также выполнять иные функции, имеющие отношение к дофаминергической системе.
Большинство перечисленных генов может также выполнять иные функции, не имеющие отношение к дофаминергической системе.
После производства и тестирования, микрочиповая платформа “NeuroStem” была активно использована в экспериментах по изучению транскриптома клеток разных типа. В полном соответствии с назначеним микрочиповой платформы, воплощенном в её дизайне, она была, в частности применена в сериях экспериментов по изучению транскриптома эмбриональных стволовых клеток человека (чЭСК), в том числе дифференцированных in vitro в дофаминергические нейроны, клеток-предшественников дофаминергических нейронов, «питающих» (фидерных) клеток используемых для экспансии чЭСК in vitro, стромальных клеток, и др. Используя комплекс аналитического программного обеспечения BioArray Software Environment database (BASE; Saal L. H., et al., 2002), производили селекцию данных, с вычитанием фонового сигнала и сигнала с точек нанесения генетического материала, помеченных «флагами». Полученные данные обладали высоким уровнем достоверности: так, например, в экспериментах с недифференцированными чЭСК коэффициент корелляции составлял для технических репликатов > 0,96, а в экспериментах с противопоставлением транскриптома недифференцированных чЭСК контролю со «сменой цвета» он был равен 0,78. В ходе экспериментов было получено подтверждение экспрессии в недифференцированных чЭСК линии SA002 многих признанных маркеров стволовых клеток (Hesx1, Grem1, Gja1, Zic3 и др.). С другой стороны, при дифференцировке чЭСК в функциональные дофаминергические нейроны было отмечено снижение уровня экспрессии многих маркерных генов стволовых клеток (Hesx1, Grem1, Dnmt3b, Utf1 и Nanog) и динамическое повышение (Log2 >1,0) уровня экспрессии генов имеющих отношение к дофаминергической системе (а именно, Igf1, Ptx3, Th, Wnt1, Lhx1 (Lim1), Drd2, Gfra1, Adcy7, Bcl11a, Col11a1, Doc2a, Drip78 (Dnajc14), Ghr, Igfbp4, Kcnab1, Moxd1, Mpp3, Nrp2, Nt (Nts), Ntn (Nrtn), Plagl1 (Zac), Slc17a6 (Vglut2), Trhr, Zdhhc2). Разнообразие молекулярных мишеней представленных на микрочиповой платформе “NeuroStem” и отвечающих изменением уровня экспрессии на дифференцировку чЭСК в функциональные дофаминергические нейроны полностью отвечает сложности биологических механизмов, вовлеченных в это направление дифференцировки. После 16 дней протокола in vitro дифференцировки, клеточная популяция является чрезвычайно гетерогенной. Лишь около 0,2% содержащихся в ней клеток экспрессируют тирозингидроксилазу (TH+) и способны синтезировать дофамин. Одновременно, в культуре находятся резидуальные (остаточные) недифференцированные чЭСК; остальные же клетки являются промежуточными типами разного уровня дифференцировки.

Рис. 2. Фрагмент микрочиповой платформы “NeuroStem”. ДНК-материал визуализирован (фактический эксперимент по анализу транскриптома недифференцированых чЭСК линии SA002). Репрезентативный блок (31 х 31 точка). Белые квадраты указывают на повторяющиеся как квадрипликаты группы молекулярных мишеней, разнесенные в пространстве. Параметры печати: расстояние между центрами точек нанесения генетического материала = 140 мкм; средний размер точек = 90-110 мкм.
С целью подтверждения результатов экспериментов с микрочиповой платформой была проведена серия экспериментов с использованием технологии ОТ-ПЦР, в которой были использованы те же образцы РНК. Было проведено исследование экспрессии в образцах следующих групп репрезентативных генов: снижение уровня экспрессии, Sox2, En1 и Nanog; без изменений, Gadph, Aldh1a1, Sdha, Tubb и Nestin; повышение уровня экспрессии, Actb, Th, Msx1 и Pitx2. Среднее нормализованное отношение интенсивности сигнала микрочиповой платформы для дифференцированных и недифференцированных чЭСК соответствовало результатам анализа продукта ОТ-ПЦР для всех групп мишеней. Принимая во внимание разные принципы, лежащие в основе этих двух технологий (гибридизация и амплификация, соответственно) данные этого эксперимента подтверждают высокую надёжность проводимого с использованием микрочиповой платформы “NeuroStem” качественного и количественного анализа динамики экспрессии генов на ранних этапах дифференцировки чЭСК в дофаминергические нейроны.
Дифференцировка стволовых клеток в функциональные дофаминергические нейроны in vitro
Использованная технология направленной дифференцировки эмбриональных стволовых клеток человека (чЭСК) в дофаминергические нейроны была основана на принципе, «индуцирующей активности, ассоциированной со стромальными клетками» (stromal cell-derived inducing activity, SDIA) [Kawasaki H., et al., 2000]. чЭСК культивировали в сложной клеточной системе, совместно со стромальными клетками мыши линии РА6. В ходе ко-культивирования, чЭСК продолжали активно пролиферировать (с активным ростом исходных колоний) и одновременно дифференцироваться. Последний процесс сопровождался значимыми морфологическими изменениями. В частности, для чЭСК линии SA002 было отмечено появление «кратерообразных» структур, а после продолжительного ко-культивирования наблюдалась массивная компартментализация производных чЭСК, с формированием в пределах почти каждой колонии множественных отсеков большей или меньшей величины и сложности, иногда с выделением настоящих цист. В отдельных случаях наблюдалось также отрастание от колоний нейритов типичной морфологии, иногда сливающихся в тяжи (Рис. 3). Встречались и морфологически смешанные типы колоний производных чЭСК.

Рис. 3. Примеры «кратерообразных» структур, компартментализации колоний и формирования тяжей нейритов, формируемых дифференцирующимися чЭСК линии SA002 при ко-культивировании со стромальными клетками мыши линии РА6. Шкала = 100 мкм.
Эффективность принятого протокола направленной дифференцировки чЭСК в функциональные дофаминергические нейроны подтверждалась следующими путями: 1) Экспрессия производными чЭСК биохимических маркеров нейронов и дофаминергических нейронов: тирозингидроксилазы (ТН) и -III-тубулина (TUBB3) клетками человеческого происхождения (окраска на ядерный белок человека), - иммуноцитохимический анализ. 2) Экспрессия производными чЭСК генетических маркеров нейронов и дофаминергических нейронов: Aldh1, Msx1, Th, и других, - применение специализированной олигонуклеотидной микрочиповой платформы “NeuroStem” и ОТ-ПЦР. 3) Синтез производными чЭСК дофамина, - хроматографический анализ (HPLC).
После ко-культивирования с клетками линии РА6 в течение 2,5 недель, 42+20% (среднее значение + стандартное отклонение) колоний чЭСК экспрессировали маркер незрелых нейронов -III-тубулин (TUBB3), и 38+22% - маркер функциональных дофаминергических нейронов тирозингидроксилазу (TH); данные о составе клеточной популяции (производных чЭСК) в целом представлены в таблице 2. После ко-культивирования с клетками линии РА6 в течение 6 недель лишь 1,0+2,0% TH+ клеток экспрессировали периферин, маркер нейронов периферической нервной системы (ПНС). Это указывает на то, что подавляющее большинство TH+ нейронов полученных с использованием данного протокола имеют фенотип, характерный для нейронов центральной нервной системы (ЦНС). Иммуноцитохимический анализ позволил также установить, что экспрессирующие тирозингидроксилазу нейроны-производные чЭСК ко-экспрессируют везикулярный транспортер моноаминов 2 (VMAT-2), вовлеченный в везикулярный транспорт дофамина и транскрипционный фактор PITX3, необходимый для развития дофаминергических нейронов substantia nigra. В совокупности, эти данные подтверждают успех используемого протокола направленной дифференцировки чЭСК в функциональные дофаминергические нейроны.
Таблица 2
Процентная доля -III-тубулин+ и ТН+ клеток в клеточной суспензии,
подготовленной для трансплантации
| Длительность ко-культивирования | |||
| 16 дней | 20 дней | 23 дня | |
| Процентная доля -III-тубулин+ клеток от общего числа клеток | 8,8 | 31,9 | 20,0 |
| Процентная доля ТН+ клеток от числа -III-тубулин+ клеток | 1,9 | 13,7 | 37,1 |
| Процентная доля ТН+ клеток от общего числа клеток | 0,2 | 4,4 | 7,4 |
Исследование биологических свойств фактора роста фибробластов 20 (FGF-20)
Фактор роста фибробластов 20 (FGF-20) был открыт в 2000 г. [Kirikoshi H., et al., 2000]. Было показано, что этот фактор преимущественно экспрессируется в substantia nigra pars compacta головного мозга [Ohmachi S., et al., 2000]; в связи с этим, нами была выдвинута гипотеза о том, что FGF-20 играет важную роль в дифференцировке нейрональных клеток-предшественников в дофаминергические нейроны и установлено, что этот рецептор экспрессируется недифференцируемыми чЭСК, стромальными клетками мыши линии РА6,
и производными чЭСК, дифференцированными в дофаминовые нейроны, что может указывать на способность FGF-20 непосредственно влиять на чЭСК и на дофаминергические нейроны. Мы также предположили, что FGF-20 может стимулировать синтез и секрецию стромальными клетками линии РА6 факторов, способствующих дофаминергической дифференцировке чЭСК и выживанию дофаминергических нейронов-производных чЭСК. Чтобы выявить эффект FGF-20 на чЭСК, FGF-20 добавляли в питательную среду при ко-культивировании чЭСК линии SA002 со стромальными клетками мыши линии РА6.
В пилотной серии экспериментов исследовали эффект, оказываемый FGF-20 в разных концентрациях (100 пг/мл, 1 нг/мл и 10 нг/мл), при этом выявили, что наилучшее соотношение эффекта/дозы достигается при добавлении FGF-20 в культуру в концентрации
1 нг/мл. После окончания этапа культивирования анализировали итоговый клеточный состав. В результате анализа морфологии колоний производных чЭСК, количества клеток-производных чЭСК после окончания протокола культивирования, профиля экспрессии биохимических и генетических маркеров, были сделаны следующие основные заключения: 1). bFGF (FGF2) является необходимым фактором пролиферации чЭСК в культуре.
В вариантах протокола, в которых bFGF не добавляли в культуральную среду, дифференцировка чЭСК в нейроны проводилась в соотношении близком к 1:1 (то есть, большинство исходных клеток дифференцировали в нейроны, но не пролиферировали).
2). FGF-20 является важным фактором стимуляции дифференцировки чЭСК
в дофаминергические нейроны. По сравнению с базовым протоколом (без добавления
FGF-20), эффективность протокола, основанного на добавлении в культуральную среду
FGF-20, возросла пятикратно. 3) В высоких дозах (10 нг/мл), FGF-20 способен также
в ограниченной степени стимулировать пролиферацию клеток. В меньших дозах этот эффект не выражен: так, при добавлении в культуральную среду FGF-20 в концентрации 1 нг/мл, число клеток экспрессирующих человеческий ядерный белок (HNuc), то есть производных чЭСК, ко-экспрессирующих маркер пролиферирующих клеток белок Ki67 возрастало незначительно, с 7,0+2,0 до 8,0+1,0.

Рис. 4. Пример иммуноцитохимического окрашивания колоний производных чЭСК линии SA002 ко-культивированных со стромальными клетками мыши линии РА6 в течение 21 дня; (А) без добавления FGF2 и FGF-20 и (Б) с добавлением bFGF и FGF-20 в течение всего времени культивирования. Светлые клетки экспрессируют тирозингидроксилазу (ТН)
и человеческий ядерный белок (HNuc). (В) Отношение числа клеток, экспрессирующих ТН
к числу клеток, экспрессирующих HNuc. FGF-20, 1 нг/мл, (**), p<0,002.
При добавлении в культуральную среду FGF-20, эффективность протокола направленной дифференцировки чЭСК в дофаминергические нейроны возрастает по сравнению с базовым протоколом (SDIA + bFGF), что было оценено по расчету отношения числа клеток экспрессирующих тирозингидроксилазу (ТН) к общему числу клеток, экспрессирующих человеческий ядерный белок (HNuc) (Рис. 4). При добавлении FGF-20
в концентрации 200 нг/мл, этот показатель возрос с 2,2+1,3 (базовый протокол) до 8,6+2,3; при добавлении FGF-20 в концентрации 1 нг/мл - с 3,0+1,0 до 15,0+2,0. Также отмечали возрастание экспрессии иных нейрональных маркеров, включая нестин и -III-тубулин: при добавлении FGF-20 в концентрации 200 нг/мл, эти показатели возросли с 43,3+3,7 (базовый протокол) до 83,4+2,1 для нестина, и с 6,2+1,7 до 26,5+5,5 для -III-тубулина, соответственно. В более низкой концентрации (1 нг/мл) FGF-20 повышал экспрессию клетками -III-тубулина не значимо: с 16,0+2,0 до 18,0+1,0. Эти данные позволили заключить, что FGF-20 является специфичным промотором дифференцировки чЭСК в дофаминергические нейроны, - но не промотором нейрональной дифференцировки в целом. В ходе работы мы также идентифицировали апоптотические клетки в культуре по экспрессии активной каспазы (cleaved caspase) 3, 8 и олигомеризованного белка BCL2-associated X protein (ВАХ). В культурах чЭСК/РА6 обнаруживаются многочисленные клетки, экспрессирующие активную каспазу 8 и ВАХ, однако при добавлении в культуральную среду FGF-20, такие клетки практически не обнаруживаются, а клетки экспрессирующие активную каспазу 3 обнаруживаются гораздо реже: снижение с 2,5+0,2 до 1,2+0,5. Это указывает на то, что снижение частоты апоптотических событий в культуре под влиянием нейротрофической активности FGF-20 может повысить выживаемость дофаминергических нейронов.
Трансплантация производных чЭСК модельным животным и анализ результатов трансплантации
На следующем этапе работы объектом исследования являлась экспериментальная модель возрастной нейродегенерации, основанная на использовании 6-гидроксидофамина
(6-OHDA) (Ungerstedt U., 1968). Крысам проводили стереотаксические инъекции токсина
в область полосатого тела. Поскольку поражение одного полосатого тела (проекции дофаминергических нейронов substantia nigra) вызывает «гемипаркинсонизм», крысы не инвалидизируются полностью. Оценку эффективности поражения дофаминергической системы (полноты токсического поражения дофаминергических нейронов) проводили
3-4 недели спустя, методом исследовании ротационного поведения со стимуляцией амфетамином, с использованием автоматизированной системы Rotomax Analyzer. В ходе отработки хирургической техники, эффективность модели дополнительно подтверждали окраской препаратов головного мозга модельных животных на тирозингидроксилазу (ТН).
В течение 6 недель после подготовки модельных животных, им производили инъекции клеточной суспензии производных чЭСК линии SA002, содержащие дофаминергические нейроны. Используя технику стереотаксической хирургии, животным модельным
в отношении локализованной, ассоциированной с возрастом нейродегенеративной патологии вводили в область полосатого тела 100 000 жизнеспособных клеток (2 мкл клеточной суспензии). При этом были сформированы следующие экспериментальные группы общей численностью 38 крыс: 1) Трансплантация производных чЭСК, длительность протокола
in vitro дифференцировки в дофаминергические нейроны = 16 дней; 2) Трансплантация производных чЭСК, длительность протокола in vitro дифференцировки в дофаминергические нейроны = 20 дней; 3) Трансплантация производных чЭСК, длительность протокола in vitro дифференцировки в дофаминергические нейроны = 23 дня (табл. 2). Клеточный материал, не использованный для трансплантации модельным животным, применяли для засева плашек, и изучали для уточнения клеточного состава пула использованных для трансплантации клеток. Через 2 недели после трансплантации, по 4 крысы из каждой группы умертвляли для проведения гистологического исследования, направленного на изучение выживаемости и пролиферации донорских клеток. Остальных крыс использовали для исследования моторных функций в динамике и умертвляли через 13 недель после трансплантации; крысы, погибшие от онкологических осложнений трансплантации, также использовались для изготовления гистологических препаратов.
Окраска гистологических образцов антителами к маркерам ядер человеческих клеток по DAB подтверждала наличие донорских клеток в анатомических структурах, являющихся мишенью трансплантации с миграцией в латеральном направлении. Иммуногистохимическое исследование подтвердило выживаемость части донорских клеток, являющихся производными чЭСК: в том числе глиальных клеток (астроцитов, маркер S100), нейронов (нейрон-специфический ядерный антиген, NeuN), и дофаминергических нейронов (ТН). При этом отмечали, что выживаемость собственно дофаминергических нейронов была лучшей (~15,0%) в группе 1 (16 дней); хуже (~0,5%) в группе 2 (20 дней) и худшей (~0,3%) в группе 3 (23 дня) (р<0,05). В то же время, донорские клетки
не экспрессировали маркеров иных типов нейронов, в том числе холинергических (холинацетилтрансфераза, CHAT), ГАМК-эргических (глутаматдекарбо-ксилаза, GAD) и серотонинергических (5-гидрокситриптамин, 5-HT), и глиальных клеток, включая олигодендроциты (гомолог хондроитин-сульфат протеогликана NG2); идентифицированы клетки таких типов не коэкспрессировали человеческий ядерный антиген (HNuc) и принадлежали животным-реципиентам. Чрезвычайную важность имела информация о том, что в отдельных трансплантатах число клеток экспрессирующих человеческий ядерный белок в 2,5-8,0 раз превосходило число клеток, введенных в ходе трансплантации, свидетельствуя о пролиферации донорских клеток после трансплантации. Для того чтобы оценить пролиферативную активность производных чЭСК, в течение последних суток перед умерщвлением, модельным животных вводили 5-бром-2-дезоксиуридин (БДУ; 3 инъекции через 8 часов), визуализируемый в препаратах головного мозга иммуногистохимическими методами. Спустя 2 недели после трансплантации, все трансплантаты содержали значительное количество меченных БДУ клеток (средние значения: группа 1, 30%; группа
2, 6%; группа 3, 12%). Однако спустя 13 недель после трансплантации, доля меченных БДУ клеток снизилась (группы 2 и 3, 2%), что свидетельствует об истощении пролиферативного потенциала клеток донорского происхождения в условиях микроокружения трансплантата. Дополнительное подтверждение пролиферативного потенциала донорских клеток в составе трансплантата получили анализом экспрессии в клетках транскрипционных факторов POU domain,>

Рис. 5. Развитие злокачественной тератомы (тератокарциномы) в сайтах трансплантации. (А-Б) Макроскопическая картина, (А) аксиальная, (Б) сагиттальная. Видимый объем опухоли очерчен белым пунктиром. (В) Микроскопическая картина.
Необходимо отметить, что данное исследование стало первым, в котором было доказано наличие обратной корреляции между длительностью пребывания чЭСК в культуре in vitro (как функции степени их дифференцировки) и риском развития злокачественной тератомы (солидной тератокарциномы) в сайте трансплантации производных чЭСК. Исследование же динамики моторных функций модельных животных не выявило даже тенденции к улучшению ни в одной из групп. Это, несомненно, обусловлено неадекватным количеством функциональных дофаминергических нейронов, переживших трансплантационную процедуру по сравнению с объемом токсического поражения. В этом отношении, полученный результат совпал с результатами более ранних экспериментов по трансплантации производных чЭСК структуры головного мозга животных, модельных в отношении возрастной нейродегенерации, в которых число выживших дофаминергических нейронов донорского происхождения было минимальным [Zeng X., et al., 2004] или равно нулю [Park C. H., et al., 2005]. Таким образом, исход трансплантации (в аспектах выживания клеток и развития опухолей) прямо зависит от уровня дифференциации используемых клеток-производных ЭСК, что представляет собой трудноразрешимый технологический парадокс. С одной стороны, по сравнению с незрелыми клетками, относительно зрелые дофаминергические нейроны (то есть клетки на высоком уровне дифференциации) являются в значительно большей степени чувствительными к стрессам, неизбежно сопровождающим работу с клетками в ходе трансплантационной процедуры, а также на непосредственно предшествующем ей этапе (подготовка клеточной суспензии) и после трансплантации [Brundin P., et al., 2000]. Можно предположить, что чистая популяция дофаминергических нейронов является значительно менее устойчивой к подобным стрессам, по сравнению
с гетерогенной популяцией, содержащей в том числе нейральные клетки других типов, способные оказывать дофаминергическим нейронам некоторую трофическую поддержку.
С другой стороны, в случае, если в используемой для трансплантации клеточной популяции содержатся резидуальные клетки с низким уровнем дифференциации, они могут стать причиной возникновения в сайте трансплантации злокачественной тератомы или нетератомных опухолей, с фатальными последствиями для реципиентов. Следовательно, поиск «окна возможности» проведения успешной трансплантации в уровне дифференциации дофаминергических нейронов-производных чЭСК (Рис. 6) является необходимым этапом отработки основанных на применении этих клеток подходов к заместительной клеточной терапии БП.
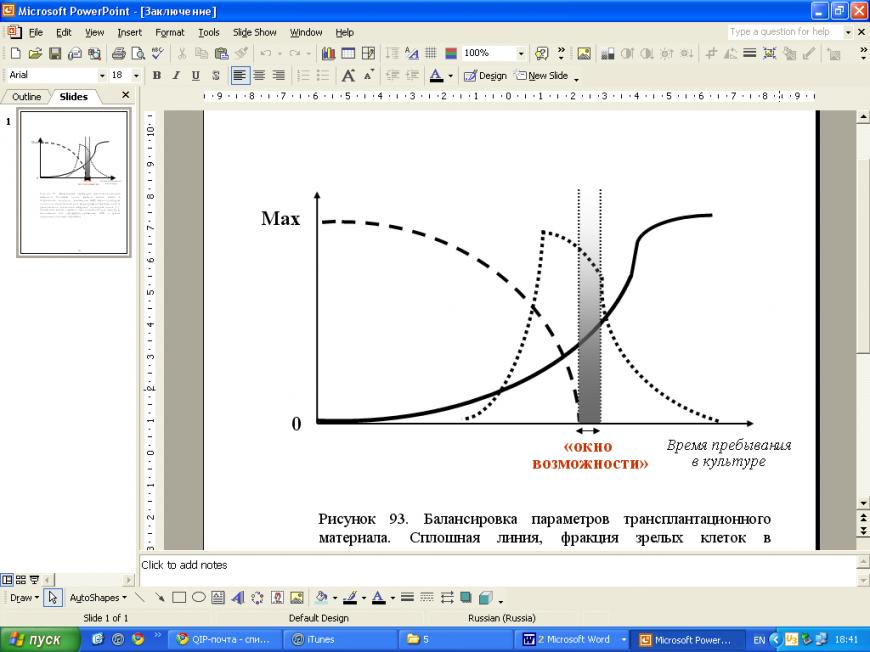
Рис. 6. Балансировка параметров трансплантационного материала. Сплошная линия, фракция зрелых клеток в гетерогенной популяции производных ЭСК; штрих-пунктирная линия (---), относительный риск формирования злокачественной тератомы в сайте трансплантации клеточного материала; пунктирная линия (…), способность клеток пережить трансплантационную процедуру (минимальна для недифференцированных ЭСК и зрелых нейронов с длинными отростками).
Получение «трансплантационных единиц» и их трансплантация модельным животным
Концепция применения «трансплантационных единиц» (Transplant Units, TU) была предложена автором как мера радикального повышения выживаемости нейронов-производных чЭСК при трансплантации. Выше отмечалось, обсуждалось, что массовая гибель донорских нейронов в сайте трансплантации (острая или отсроченная) является одним из критичных факторов, препятствующих успеху заместительной клеточной терапии возрастной нейродегенерации. Многочисленные вариации используемых протоколов направленной дифференцировки чЭСК в дофаминергические нейроны основаны
на культивировании клеток in vitro в течение продолжительного времени, и после накопления в клеточной популяции значительного количества функциональных дофаминергических нейронов, - диссоциация клеток и трансплантация клеточной суспензии. При этом жизнеспособность клеток резко снижается (до <2%) в ходе диссоциации исходной клеточной культуры с получением высококачественной суспензии (содержащей единичные клетки и минимальные (по 2-3 клетки) агрегаты клеток). Этот эффект может в значительной степени быть опосредован аноикисом: использующей механизмы апоптоза клеточной гибелью в результате утери межклеточных контактов и контактов с внеклеточным матриксом [Raff M. C., 1992; Meredith J. A., et al., 1993]. Выживаемость же зрелых дофаминергических нейронов (выделенных из эмбрионального/фетального мезенцефалона) является значительно меньшей, чем выживаемость незрелых нейронов, что обусловлено аксотомией. Между тем, длительность пребывания клеток в культуре обратно пропорциональна риску развития онкологических осложнений трансплантации. Отсюда, выбор популяции клеток-производных чЭСК не способных вызвать развитие онкологических осложнений в сайте трансплантации (безопасных), но при этом хорошо переносящих трансплантацию (эффективных) представляет собой серьезную проблему. Теоретически, применение агрегатов клеток-производных чЭСК могло бы позволить повысить выживаемость дофаминергических нейронов. В случае если бы конечной точкой протокола дифференцировки чЭСК in vitro в функциональные дофаминергические нейроны, могли стать не индивидуальные клетки (адгезивная культура), а агрегаты клеток (суспензионная культура), можно было бы избежать диссоциации клеток. При этом не проводилось бы энзиматическое и механическое воздействие на клетки; сохранялись бы установившиеся межклеточные контакты в пределах микроокружения дофаминергических нейронов, в том числе между нейронами и иными типами клеток.
Подобные агрегаты клеток-производных чЭСК должны удовлетворять следующим ключевым требованиям: 1) содержать максимально возможное число функциональных дофаминергических нейронов: т.е. эффективность протокола должна быть сравнимой
со стандартным протоколом; 2) не содержать резидуальных недифференцированных чЭСК; 3) быть достаточно малого размера для проведения трансплантации с использованием отработанной практики (т.е. проходимыми через носик микропипетки и иглу Гамильтоновского шприца); 4) быть достаточно большого размера, чтобы содержать значительно число клеток; 5) являться в значительной степени однородными
по величине/числу содержащихся клеток.
Поскольку эффективное формирование агрегатов недифференцированных чЭСК
(с последующей дифференцировкой в суспензионной культуре) с использованием стандартных методов (спонтанное образование ЭТ, «висячая капля», и т.д.) оказалось невозможным, было принято решение разделить протокол на 2 этапа:
Этап I: ко-культивирование чЭСК в течение 14-17 дней со стромальными клетками мыши линии РА6, с дифференцировкой в нейрональные/ дофаминернические прогениторные клетки, как описано выше; Этап II: перевод адгезивной культуры в суспензионную, с формированием в течение 6-7 дней «трансплантационных единиц» с заданными свойствами, содержащих дофаминергические нейроны и постмитотические дофаминергические прогениторные клетки. В основу протокола были положены несколько ключевых факторов:
1) Нейрональные/ дофаминернические прогениторные клетки значительно более устойчивы к диссоциации по сравнению с недифференцированными чЭСК и зрелыми дофаминергическими нейронами;
2) Получение большого количества таких клеток отработано (базовый протокол, основанный на принципе SDIA) и не вызывает значительных технических трудностей; Формированию агрегатов способствует непрерывное перемешивание клеточной суспензии непосредственно в СО2-инкубаторе, что обеспечивается обладающим уникальными характеристиками шейкером ES-X (Khner AG, Швейцария; [Berglund C. M., et al., 2004];
3) Поддержанию клеток (а затем агрегатов) в суспензии способствует применение культурального пластика с особыми физическими свойствами;
4) На Этапе II протокола возможно продолжение индукции дофаминергической дифференцировки производных чЭСК за счет применения приспособленной (кондиционированной) питательной среды, обогащенной растворимыми факторами, секретируемыми стромальными клетками мыши линии РА6.
В результате отработки технологии, основанной на перечисленных выше ключевых факторах удалось получить однородную популяцию агрегатов клеток-производных чЭСК («трансплантационных единиц»), обладающих заданными свойствами и готовых
к непосредственной трансплантации (без диссоциации). В то время как технология Этапа I и диссоциации клеток папаином и аккутазой была отработана полностью как описано выше,
в ходе отработки технологии Этапа II, испытывали вариации всех ключевых параметров, включая следующие: 1) концентрацию клеточной суспензии: 50 000-500 000 кл/мл;
2) скорость перемешивания клеточной суспензии шейкером: 20-200 об/мин; 3) режим работы шейкера; 4) тип используемой питательной среды; 5) технология смены питательной среды; 6) параметры культурального пластика на подэтапах; 7) длительность каждого подэтапа
(II-a - II-в; по 2 или 3 дня) и Этапа II в целом (6, 7, 8 дней). Все эти параметры были отработаны в ходе пилотных экспериментов. После окончания Этапа I и диссоциации, суспензию клеток (во всех случаях жизнеспособность этих клеток составляла 66-88%) переносили в чашки Петри малого диаметра, изготовленных из пластика высокой адгезивности. Таким образом, на этом этапе (подэтап II-а) на суспензию клеток воздействовало перемешивание, но при этом клетки стремились прикрепиться дну чашки Петри. В результате, осуществлялась селекция клеток по адгезивным свойствам: высокоадгезивные стромальные клетки (контаминирующие клеточный пул стромальные клетки мыши линии РА6, используемые как фидерные на Этапе I), а вероятно и резидуальные недифференцированные чЭСК, прилипали к дну плашек, - в то время как менее адгезивные нейрональные/дофаминернические прогениторные клетки в большинстве включались в состав формируемых агрегатов клеток. После окончания подэтапа II-а, сформировавшиеся агрегаты клеток переносили из чашек Петри, изготовленных из пластика высокой адгезивности, в чашки Петри, изготовленные из пластика низкой адгезивности.
В результате, на этом этапе (подэтап II-б) продолжался рост агрегатов клеток, при этом проводилось отмывание мертвых клеток из суспензии. После окончания подэтапа II-б, сформировавшиеся агрегаты клеток продолжали культивировать в суспензионной культуре, заменяя питательную среду. На этом этапе (подэтап II-в) продолжался рост агрегатов клеток при непрерывном перемешивании.

Рис. 7. Получение чистой культуры «трансплантационных единиц»: день протокола 16+6. Использовались среда CNS (кондиционированная на клетках линии РА6 среда NS) и FGF-20 (1 нг/мл). Шкала = 100 мкм.
В результате исследования морфологии клеточных культур на подэтапах,
в соответствии с различными значениями перечисленных выше параметров была отработана технология, позволяющая получить значительное число однородных, пригодных для непосредственной трансплантации «трансплантационных единиц» (ТЕ) (Рис. 7). Иммуноцитохимическое исследование клеточного состава «трансплантационных единиц» подтвердило то, что в них широко представлены клетки нейронального ряда, в том числе экспрессирующие тирозингидроксилазу (ТН) дофаминергические нейроны. Также было выявлено, что находящимися в составе «трансплантационных единиц» клетками экспрессируются и другие маркеры, в том числе маркеры пролиферирующих клеток (Ki67, фосфорилированный гистоновый белок Н3 (РН3)) и плюрипотентных клеток (Nanog), нейронов (нестин, -III-тубулин) и дофаминергических нейронов (питуитарный гомеобоксный белок 3 (PITX3), альдегиддегидрогеназы 1 (Aldh1a1), белка сходного
с ядерным рецептором 1 (Nurr1)); при этом экспрессия маркера плюрипотентных клеток Oct3/4 обнаружена не была. Содержание нейронов (оценивалось по экспрессии
-III-тубулина) и дофаминергических нейронов (оценивалось по экспрессии ТН в ТЕ) было значительным: большой интерес представляла при этом их высокая концентрация в «корковых» областях ТЕ. Показательно, что содержание нейронов и дофаминергических нейронов в ТЕ значительно увеличивалось как при удлинении Этапа I протокола (16 или
18 дней), так и при добавлении в питательную среду FGF-20, являющимся, как показано выше, важным фактором стимуляции пролиферации производных чЭСК и выживаемости дофаминергических нейронов. Если в ТЕ сформированных в течение 16+6 дней обнаруживали 47,0+5,0% нейронов, в том числе 2,0±0,4% дофаминергических нейронов, - то в ТЕ сформированных в течение 18+6 дней обнаруживали уже 64,0+5,0% нейронов, в том числе 4,2+0,7% дофаминергических нейронов. При добавлении же в питательную среду FGF-20 (в концентрации 1 нг/мл), наблюдали дальнейшее увеличение содержания дофаминергических нейронов в ТЕ (до 13,0±2,0%). При этом увеличение содержания
в ТЕ нейронов было незначимым, что ещё раз свидетельствует в пользу специфичности действия FGF-20. Несколько репрезентативных ТЕ использовали для измерения количества содержащихся в них клеток: для этого проводили диссоциацию ТЕ в растворе аккутазы и подсчетывали число клеток в образовавшейся суспензии, с расчетом среднего значения.
В наших экспериментах, для протокола длительностью 16+6 дней, среднее значение числа клеток в одной ТЕ составило: для варианта, основанного на применении питательной среды NS и FGF-20 - 700 клеток/ТЕ; для варианта, основанного на применении питательной среды СNS (среды NS, кондиционированной на клетках линии РА6) - 14 800 клеток/ТЕ; и для варианта, основанного на применении питательной среды СNS и FGF-20 - 28 800 клеток/ТЕ.
В течение 6 недель после подготовки животных модельных в отношении локализованной, ассоциированной с возрастом нейродегенеративной патологией,
им производили инъекции ТЕ, являющихся агрегатами производных чЭСК линии SA002, содержащими дофаминергические нейроны (длительность протокола = 16+6 дней) (Рис. 7). Используя технику стереотаксической хирургии, модельным животным вводили в область полосатого тела число «трансплантационных единиц» эквивалентное 100 000 жизнеспособных клеток (общий объем около 4 мкл). Трансплантация ТЕ потребовала отработки особой техники введения основанной на применении «воздушного замка», при этом после завершения каждой трансплантации иглу шприца промывали и лаваж исследовали под микроскопом: это позволяло идентифицировать случаи, когда часть ТЕ оставались в игле шприца, и таким образом объем трансплантата снижался по сравнению
с запланированным. В ходе серий трансплантаций, техника трансплантации ТЕ была отработана, и суммарное время операции снизилось с 3-4 часов до 40 мин - 1,5 часов на серию из 6-8 крыс. При этом были сформированы следующие экспериментальные группы общей численностью 65 крыс: 1) Трансплантация «трансплантационных единиц», сформированных в присутствии FGF-20; 2) Трансплантация «трансплантационных единиц», сформированных без добавления FGF-20. В каждой серии трансплантаций проводили диссоциацию репрезентативных ТЕ для установления числа содержащихся в каждой ТЕ клеток: в соответствие с этим, каждой проводили трансплантацию 4-10 ТЕ. Так, например,
в отдельных сериях были получены следующие репрезентативные данные: 17 060 клеток/
ТЕ FGF20- (6 ТЕ/трансплантацию); 21 100 клеток/ТЕ FGF20- (5 ТЕ/трансплантацию);
19 800 клеток/ТЕ FGF20+ (5 ТЕ/трансплантацию); 11 658 клеток/ТЕ FGF20-
(9 ТЕ/трансплантацию), и т.д. Через 48 часов (2 суток) и через 2 недели после трансплантации, по 2 крысы из каждой группы в каждой серии умерщвляли для проведения гистологического исследования, направленного на изучение выживаемости и пролиферации донорских клеток. Остальных крыс использовали для исследования моторных функций в динамике, и умертвляли через 13-16 недель после трансплантации. В каждом случае, в ходе гистологического исследования выявляли наличие живых клеток донорского происхождения (т.е. клеток экспрессирующих человеческий ядерный белок, HNuc) в составе трансплантата, а также производных чЭСК экспрессирующих маркеры нейронов экспрессирующих нейрон-специфический ядерный антиген (NeuN), в том числе незрелых нейронов экспрессирующих -III-тубулин и дофаминергических нейронов экспрессирующих ТН. При этом отмечали наличие в составе трансплантата двух морфологически отличающихся друг от друга типов клеток, экспрессирующих тирозингидроксилазу (ТН+ клеток): крупных клеток с длинными отростками и относительно мелких клеток с короткими отростками. Во многих случаях отмечали наличие кластеров ТН+ клеток, в том числе содержащих значительное число клеток. В общей сложности, во трансплантатах многих животных, принадлежащих к обеим экспериментальным группам идентифицировали от 500 до 1000 экспрессирующих тирозингидроксилазу клеток донорского происхождения, т.е. переживших трансплантационную процедуру дофаминергических нейронов-производных чЭСК.
Этот результат, обусловленный принципом трансплантации ТЕ без диссоциации, в корне отличается от результата трансплантации клеточной суспензии (когда в трансплантатах идентифицировали порядка 10-50 живых ТН+ клеток). При этом общий объем трансплантата (клеточная суспензия или ТЕ) был равным (100 000 жизнеспособны клеток), а доля дофаминергических нейронов в составе гетерогенной клеточной популяции была сравнима: 4,4% и 7,4% для клеточной суспензии (длительность протокола 20 и 23 дня, соответственно) и 2,0% и 13,0% для «трансплантационных единиц» (длительность протокола 16+6 дней, без добавления FGF-20 и в присутствии FGF-20, соответственно). Двадцатикратное повышение выживаемости дофаминергических нейронов в ходе трансплантации полностью оправдывает усложнение протокола дифференцировки (двухэтапный протокол) и трансплантации (манипуляции с ТЕ в ходе собственно трансплантации) и прямо подтверждает то, что выживаемость донорских клеток является функцией не только числа пересаженных клеток, но и техники трансплантационной процедуры. Различий в числе выживших дофаминергических нейронов между группами животных-реципиентов трансплантации ТЕ сформированных в присутствии FGF-20 и без добавления FGF-20, выявлено не было. Чрезвычайно важным является отсутствие признаков развития быстро растущей, злокачественной тератомы (тератокарциномы) в сайте трансплантации ТЕ у всех животных-реципиентов. Это подтверждает то, что двухэтапный протокол формирования ТЕ адекватен в отношении удаления из клеточной среды большинства или всех резидуальных недифференцированных/плюрипотентных чЭСК.
Исследование динамики моторных функций животных модельных в отношении локализованной, ассоциированной с возрастом нейродегенеративной патологии не выявило статистически значимого улучшения в общем объеме групп реципиентов трансплантации ТЕ (эквивалент 100 000 жизнеспособных клеток). Между тем, отдельные модельные животные демонстрировали тенденцию к улучшению моторных функций, проявляющуюся в снижении соотношения между числом ипсилатеральных и контралатеральных поворотов животного
в течение ротометрического исследования (Рис. 8). Обнаружение данной тенденции, отличающей результаты эксперимента с трансплантацией «трансплантационных единиц»
от результатов всех проведенных ранее трансплантаций суспензии производных чЭСК, вероятнее всего обусловлены значительным повышением выживаемости донорских дофаминергических нейронов в ходе трансплантационной процедуры.

Рис. 8. Динамика моторных функций у модельных животных-реципиентов трансплантации «трансплантационных единиц», по данным ротометрического исследования перед трансплантацией (0 недель) и спустя 2, 6 и 8 недель. Показаны результаты для отдельных репрезентативных животных, продемонстрировавших тенденцию к улучшению моторных функций. Ипсилатеральный ответ рассчитан как соотношение между числом ипсилатеральных и контралатеральных поворотов в течение 90 минут исследования.
ВЫВОДЫ
1. Идентифицирована панель в составе 238 генов, контролирующих процессы пролиферации стволовых клеток в недифференцированном состоянии. Данная группа содержит в том числе новые «кандидатные» факторы контроля пролиферативного потенциала стволовых клеток.
2. Показано, что по совокупности свойств (а именно, доступности исходного клеточного материала; этическим препятствиям; сложности получения линий клеток; пролиферативному потенциалу; эффективности экспансии клеточного материала; уровню пластичности; иммуногенности; возможности направленной дифференцировки
в функциональные дофаминергические нейроны; и другим) эмбриональные стволовые клетки человека (чЭСК) линии SA002 наиболее удобны для размножения (экспансии) in vitro в интересах применения в качестве субстрата клеточной терапии локализованной возрастной нейродегенеративной патологии;
3. Выявлена биологическая роль фактора роста фибробластов 20 (FGF-20), проявляющейся в стимуляции пролиферации, дифференцировки и выживания дофаминергических прогениторных клеток в культуре in vitro. Применение этого фактора повышает эффективность протокола направленной дифференцировки чЭСК
в дофаминергические нейроны in vitro с 3,0+1,0% экспрессирующих тирозингидроксилазу клеток от общего числа клеток-производных чЭСК (базовый протокол) до 15,0+2,0% (применение FGF-20 в концентрации 1 нг/мл питательной среды);
4. Установлена обратная зависимость между длительностью пребывания чЭСК
в культуре in vitro (как функции степени их дифференцировки) и риском развития злокачественной тератомы (тератокарциномы) в сайтах трансплантации клеточного материала, что позволяет выбрать условия дифференцировки клеток, обеспечивающие минимизацию риска развития злокачественной тератомы у реципиентов, и является одним из решающих факторов возможности использования клеточного материала в лечении больных, в том числе пожилого и старческого возраста;
5. Разработан принципиально новый двухэтапный протокол дифференцировки чЭСК в функциональные дофаминергические нейроны in vitro (I этап - дифференцировка
в адгезионной культуре, II этап - в суспензионной культуре), который позволил получить однородную популяцию клеточных агрегатов с заданными характеристиками («трансплантационных единиц»), содержащих большое количество дофаминергических нейронов (до 13,0+2,0% от общего числа клеток) и не требующих диссоциации при трансплантации;
6. Разработана и апробирована в эксперименте с использованием модельных животных техника трансплантации «трансплантационных единиц», применение которой впервые дало возможность многократно (до 20 раз) повысить выживаемость дофаминергических нейронов в сайтах трансплантации, что позволило получить функциональный эффект у модельных животных и свидетельствует о перспективности применения разработанной технологии для лечения ассоциированных с возрастом нейродегенеративных заболеваний у человека.
Практические рекомендации
1. Учитывая общее постарение населения развитых стран, и связанный с этим рост распространенности локализованной, ассоциированной с возрастом нейродегенеративной патологии, а также учитывая постепенное снижение эффективности фармакотерапии проводимой в течение длительного времени, продолжение работ по разработке основанных на принципах клеточной терапии подходов к лечению таких заболеваний является в высшей мере целесообразным;
2. Эмбриональные стволовые клетки человека (чЭСК) могут быть использованы в качестве исходного субстрата клеточной терапии локализованной, ассоциированной с возрастом нейродегенеративной патологии. При этом целесообразно использовать линии чЭСК, способные эффективно пролиферировать вне присутствия фидерных (питающих) клеток ксеногенного происхождения (например линию SA002);
3. В ходе направленной дифференцировки чЭСК в дофаминергические нейроны in vitro, рекомендуется вводить в состав питательной среды фактор роста фибробластов 20 (FGF-20) в концентрации 1 нг/мл питательной среды;
4. Специализированную микрочиповую платформу “NeuroStem” целесообразно использовать в экспериментах по изучению фундаментальных биологических механизмов, лежащих в основе пролиферации, роста, дифференцировки и созревания клеток, а также процессов нейродегенерации, в том числе возрастной. Это является необходимым звеном работы по разработке основанных на принципах клеточной терапии подходов к лечению таких заболеваний;
5. В ходе разработки подходов к клеточной терапии локализованной, ассоциированной с возрастом нейродегенеративной патологии рекомендуется использовать производные чЭСК в формате не клеточной суспензии, а микроагрегатов, - то есть без диссоциации на этапе непосредственно предшествующем трансплантации. При этом рекомендуется использовать разработанный метод получения «трансплантационных единиц» обладающих комплексом заданных физических и биологических свойств.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ:
Статьи в журналах, включенных в Перечень ВАК Минобрнауки РФ
- Анисимов С.В. Клеточная терапия болезни Паркинсона: I. Трансплантация эмбриональной и взрослой ткани // Успехи геронтологии. - 2008. – T. 21 (4). – C. 575-592.
- Анисимов С.В. Клеточная терапия болезни Паркинсона: II. Применение соматических стволовых клеток // Успехи геронтологии. - 2009. - T. 22 (1). - C. 150-166.
- Анисимов С.В. Клеточная терапия болезни Паркинсона: III. Применение неонатальных, фетальных и эмбриональных стволовых клеток // Успехи геронтологии. - 2009. - T. 22 (2). - C. 296-315.
- Анисимов С.В. Клеточная терапия болезни Паркинсона: IV. Риски и перспективы // Успехи геронтологии. - 2009. - T. 22 (3). - C. 418-439.
- Получение дофаминовых нейронов из эмбриональных стволовых клеток человека in vitro/ И.В. Кожухарова, В.И. Земелько, З.В. Ковалева, Н.А. Пуговкина, Л.Л. Алексеенко, М.В. Харченко, Н.Д. Аксенов, А.Н. Шатрова, С.В. Анисимов, Т.М. Гринчук, И.И. Фридлянская, Н.Н. Никольский// Цитология. - 2010. - Т. 52 (10). - C. 875-882.
- Anisimov S.V. A large-scale screening of the normalized mammal mitochondrial gene expression profiles/ S.V. Anisimov// Genetical Research. – 2005. – V. 86 (2). – P. 127-138.
- Anisimov S.V. Cell-based therapeutic approaches for Parkinson’s disease: progress and perspectives/ S.V. Anisimov// Reviews in the Neurosciences. - 2009. - V. 20 (5-6). - P. 347-381.
- Anisimov S.V. Incidence of "quasi-ditags" in catalogs generated by Serial Analysis of Gene Expression (SAGE)/ S.V. Anisimov, A.A. Sharov// BMC Bioinformatics. – 2004. – V. 5(1). – 152.
- Anisimov S.V. Serial Analysis of Gene Expression (SAGE): 13 Years of Application in Research/ S.V.Anisimov// Current Pharmaceutical Biotechnology. - 2008. - V. 9 (5). - P. 338-350.
- Anisimov S.V. Serial Analysis of Gene Expression (SAGE). Editorial/ S.V. Anisimov// Current Pharmaceutical Biotechnology. – 2008. – V. 9 (5). – P. 337.
- Anisimov S.V. Risks and mechanisms of oncological disease following stem cell transplantation/ S.V. Anisimov, A. Morizane, A.S. Correia// Stem Cell Reviews and Reports. – 2010. – V. 6 (2). – P. 411-424.
- Differentiation of pluripotent embryonic stem cells into cardiomyocytes/ K.R. Boheler, J. Czyz, D. Tweedie, H.-T. Yang, S.V. Anisimov, A.M. Wobus// Circulation Research. - 2002. - Vol. 91 (3). - P. 189-201.
- Linkage of pluripotent stem cell-associated transcripts to regulatory gene networks/ K.V. Tarasov, G. Testa, Y.S. Tarasova, G. Kania, D.R. Riordon, M. Volkova, S.V. Anisimov, A.M. Wobus, K.R. Boheler // Cells Tissues Organs. – 2008. – V. 188 (1-2). – P. 31-45.
- «NeuroStem Chip»: a novel highly specialized tool to study neural differentiation pathways in human stem cell/ S.V. Anisimov, N.S. Christophersen, S.A. Correia, J.Y. Li, P. Brundin// BMC Genomics. – 2007. – V. 8 (1). – P. 46.
- SAGE identification of differentiation responsive genes in P19 embryonic cells induced to form cardiomyocytes in vitro/ S.V. Anisimov, K.V. Tarasov, D. Riordon, A.M. Wobus, K.R. Boheler// Mechanisms of Development. – 2002. – V. 117 (1-2). – P. 25-74.
- Signals from embryonic fibroblasts induce adult intestinal epithelial cells to form nestin-positive cells with proliferation and multilineage differentiation capacity in vitro. 1, Joint first authorship/ C. Wiese 1, A.Rolletschek 1, G. Kania, A. Navarrete-Santos, S.V. Anisimov, B. Steinfarz, K.V. Tarasov, S.A. Brugh, I. Zahanich, C. Ruschenschmidt, H. Beck, P. Blyszczuk, J. Czyz, J. Huebach, U. Ravens, O. Horstmann, L. St-Onge, T. Braun, O. Brustle, K.R. Boheler, A.M. Wobus// Stem Cells. – 2006. – V. 24 (9). – P. 2085-2097.
- Stern M.D. Estimation of biological complexity from SAGE catalogs: statistical limitations revealed by Monte Carlo simulation/ M.D. Stern, S.V. Anisimov, K.R. Boheler// Bioinformatics. – 2003. – V. 19 (4). – P. 443-448.
- Transplantation of human embryonic stem cell-derived cells to a rat model of Parkinson’s disease: effect of in vitro differentiation on graft survival and teratoma formation. 1, Joint first authorship; 2, Joint last authorship/ A. Brederlau1, A.S.Correia1, S.V. Anisimov, M. Elmi, G. Paul, L. Roybon, A. Morizane, P. Bergkvist, I. Riebe, U. Nannmark, M. Carta, E. Hanse, J. Takahashi, Y. Sasai, K. Funa, P. Brundin, P.S.Eriksson 2, J.Y.Li 2// Stem Cells. – 2006. – V. 24 (6). – P. 1433-1440.
Статьи в других журналах
- Fibroblast growth factor-20 increases the yield of midbrain dopaminergic neurons obtained from human embryonic stem cell cultures/ A.S. Correia, S.V. Anisimov, L. Roybon, J.Y. Li, P. Brundin/ Annals of Medicine// Frontiers in Neuroanatomy. – 2007. – V. 1 (1). – P. 4.
- Growth factors and feeder cells promote differentiation of human embryonic stem cells into dopaminergic neurons: a novel role for fibroblast growth factor-20/ A.S. Correia, S.V. Anisimov, J.Y. Li, P. Brundin// Frontiers in Neuroanatomy. – 2008. – V. 2 (1). – P. 26-34.
- Identification of molecules derived from human fibroblast feeder cells that support proliferation of human embryonic stem cells. 1, Joint first authorship/ S.V.Anisimov 1, N.S.Christophersen 1, A.S. Correia, V.J. Hall, I. Sandelin, J.Y. Li, P. Brundin// Cellular and Molecular Biology Letters. – 2011. – V. 16 (1). – 79-88.
- Stem cell-based therapy for Parkinson’s disease/ A.S. Correia, S.V. Anisimov, J.Y. Li, P. Brundin/ Annals of Medicine. – 2005. – V. 37 (7). – P. 487-498.
Монографии
- Анисимов С.В. Микрочиповая технология в диагностике сердечно-сосудистых, гематологических и эндокринных заболеваний/ С.В. Анисимов// Нанотехнологии в биологии и медицине (Ред. Е.В.Шляхто). - СПб.: Любавич. – 2009. - C. 71-100.
- Anisimov S.V. Selecting a proper platform for microarray experiment// In: Stem Cell Applications in Diseases/ Anisimov S.V. (Eds. W.B.Burnsides, R.H.Ellsley). - NOVA Publishers, USA. – 2008. – P. 1-10.
- Anisimov S.V. Selecting a proper platform for microarray experiment// In: Oligonucleotie Array Sequence Analysis/ Anisimov S.V. (Eds. M.K.Moretti, L.J.Rizzo). - NOVA Publishers, USA. – 2008. – P. 27-36.
- Being realistic about human embryonic stem cell-based therapy of Parkinson’s disease/ S.V. Anisimov, A.S. Correia, J.Y. Li, P. Brundin// In: Parkinson's Disease and Movement Disorders. 5th Edition/ J.J.Jankovic, E.Tolosa (Eds). - Philadelphia, PA, USA: Lippincott Williams & Wilkins. – 2006. – P. 642-652.
- SAGE analysis of embryonic stem cells/ K.V. Tarasov, S.V. Anisimov, Y.S. Tarasova, D.R.Riordon, A. Younes, M.D. Stern, A.M. Wobus, K.R. Boheler// In: Serial Analysis of Gene Expression: Current Technologies and Applications/ S.M.Wang (Ed). - Norwich, UK: Horizon Scientific Press. – 2005. – P. 207-228.
Методические руководства
- Anisimov S.V. Application of DNA microarray technology to gerontological studies/ S.V. Anisimov// Methods in Molecular Biology. – 2007. - V. 371. – P. 249-265.
- Anisimov S.V. Serial Analysis of Gene Expression (SAGE): Detailed protocol and analysis/ S.V. Anisimov, K.V. Tarasov, K.R. Boheler// In: Cell Biology: A Laboratory Handbook. 3rd Edition/ J.Celis (Ed). - Academic Press, San Diego, CA, USA. – 2005. – P. IV.14.1-IV.14.10.
Тезисы докладов
- Анисимов С.В. Клеточные технологии: реалии сегодняшнего дня/ С.В. Анисимов// Матер. конф. IV-я Международная конференция «Медицина долголетия и качества жизни». 17-18 апреля 2008 г., Москва, Россия. –2008 – C. 54.
- Анисимов С.В. Основные принципы восстановительной и заместительной клеточной терапии/ С.В. Анисимов// Матер. V-й Международной конф. «Медицина долголетия и качества жизни». 28-29 января 2009 г., Москва, Россия. – 2009. – C. 34.
- Анисимов С.В. Практические аспекты возможного применения эмбриональных стволовых клеток в клинике/ С.В. Анисимов// Всероссийская научно-практическая конф. с международным участием «Высокотехнологичные методы диагностики и лечения заболеваний сердца, крови и эндокринных органов». Артериальная Гипертензия. 23-24 мая 2008 г., Санкт-Петербург, Россия. – 2008. – T. 14 (1). – C. 93.
- Разработка способов направленной нейрональной дифференцировки эмбриональных стволовых клеток (ЭСК) человека, исключающих клетки с генетическими изменениями и/или туморогенным потенциалом/ Н.Н. Никольский, В.А. Поспелов, Т.М. Гринчук, Г.Г. Полянская, Т.В. Поспелова, С.В. Анисимов, Н.А. Михайлова, М.С. Лянгузова, И.В. Кожухарова, Т.А. Крылова, Л.Л. Алексеенко, З.В. Ковалева, Н.А. Пуговкина, В.С. Романов, А.М. Кольцова, Г.С. Синева, И.И. Суворова, Н.А. Виноградова, А.Д. Харазова, А.А. Заварзин, В.Ф. Синицина, Т.В. Серговская //Сб. тез. Итоговой конференции по результатам выполнения мероприятий за 2009 г. в рамках приоритетного направления «Живые системы» ФЦП «Исследования и разработки по приоритетным направлениям развития научно-технологического комплекса России на 2007-2012 годы». 25-27 ноября 2009 г. Москва, Россия. –2009. – С. 26-27.
- Anisimov S.V. Application of microarray technology to experimental neurology/ S.V. Anisimov// From Neuron to Neurology and Psychiatry. The Baltic Summer School. University of Copenhagen, The Panum Institute. - Copenhagen, Denmark. - August 14-26, 2005. - Delegate manual. – 2005. – P. 30.
- Developing human embryonic stem cell-based therapy for Parkinson’s disease/ S. Correia, A. Brederlau, S.V. Anisimov G., Paul L., Roybon, P. Eriksson, P. Brundin, J.Y. Li//Abstract book. Meeting of SRC-SDARF-JDRF Joint Programme in Stem Cell Research. - June 16-17, 2005 Sigtuna, Sweden. – 2005. – P.19.
- Developing human embryonic stem cell-based therapy for Parkinsons disease/ S. Correia, A. Brederlau, S.V. Anisimov, G. Paul L., Roybon, P. Eriksson, P. Brundin, J.Y. Li// International Society for Stem Cell Research 3rd Annual Meeting. - San Francisco, CA, USA. - June 23-25, 2005. - Delegate manual. – 2005. – P.85.
- Discovering dopaminergic differentiation-associated gene expression profile alterations in an immortalized human mesencephalic cell line using specialized custom microarray/ N.S. Christophersen, S.V. Anisimov, B. Juliusson, J. Jrgensen, P. Brundin// Abstract book. Meeting of SRC-SDARF-JDRF Joint Programme in Stem Cell Research. - June 16-17, 2005. Sigtuna, Sweden. – 2005. – P.18.
- Discovering dopaminergic differentiation-associated gene expression profile alterations in an immortalized human mesencephalic cell line using/ Christophersen N.S., Anisimov S.V., Juliusson B., Jrgensen J., Brundin P. //International Society for Stem Cell Research 3rd Annual Meeting. - San Francisco, CA, USA. - June 23-25, 2005. - Delegate manual. – 2005. – P. 181.
- Identification of Gene Transcripts with Profiles Unique to Pluripotent Mouse R1 Embryonic Stem Cells Serial Analysis of Gene Expression/ K.R. Boheler, S.V. Anisimov, K.V.Tarasov, D. Tweedie, A.M. Wobus// Abstract Book. Stem Cells: Origins, Fates and Functions. Keystone, CO. March 17-23, 2002, Keystone Symposia. – 2002. – P.62.
- In search of stemness: expression profile of ES (R1 and D3) and EC (P19) cell lines/ K.V.Tarasov, S.V. Anisimov, Y.S. Tarasova, A.M. Wobus, K.R. Boheler//Abstr. From Stem Cells to Therapy. Steamboat Springs, CO. - March 29 - April 3, 2003. - Keystone Symposia. – 2003. –P.114.
- Survival and neuronal differentiation of human embryonic stem cells after grafting into a Parkinsonian rat model/ S. Correia, A. Brederlau J.Y., Li, L. Roybon, S.V. Anisimov, G. Paul, P. Eriksson, P. Brundin// Abstr. 4th Forum of European Neuroscience. - Lisbon, Portugal. – July, 10-14, 2004. – P. A019.13.
СПИСОК СОКРАЩЕНИЙ, ИСПОЛЬЗУЕМЫХ В АВТОРЕФЕРАТЕ
БА - болезнь Альцгеймера
БДТЛ - болезнь [диффузных] телец Леви
БП - болезнь Паркинсона
ДМСО - диметилсульоксид
ДНКаза I - деоксирибонуклеаза I
ИГХ - иммуногистохимический анализ
ИЦХ - иммуноцитохимический анализ
мЭСК - эмбриональные стволовые клетки мыши
мЭФ - эмбриональные фибробласты мыши
ППК - первичные половые клетки
ПФА - параформальдегид
ПЦР-ОТ - полимеразная цепная реакция с обратной транскрипцией
СК - стволовые клетки
ТЕ - трансплантационные единицы
ЦНС - центральная нервная система
чНФ - фибробласты крайней плоти новорожденных
чЭСК - эмбриональные стволовые клетки человека
ЭСК - эмбриональные стволовые клетки
6-OHDA - 6-гидроксидофамин
bFGF - основной фактор роста фибробластов
DMEM - среда Игла в модификации Дюльбекко
DOPAC - 3,4-дигидрофенилуксусная кислота
FGF-2 - основной фактор роста фибробластов
FGF-20 - фактор роста фибробластов 20
HBSS - сбалансированный солевой раствор Хэнкса
LIF - лейкемический ингибиторный фактор
SAGE - серийный анализ генетической экспрессии
SCF - фактор стволовых клеток
SDIA - индуцирующая активность, ассоциированная со стромальными клетками
TrBlue - трипановый синий
АНИСИМОВ Сергей Владимирович. Применение стволовых клеток в терапии возрастной нейродегенеративной патологии (экспериментальное исследование)//Автореф. дис. … докт. мед. наук: 14.01.30; 03.03.04 - СПб., 2010. - 40 с.
| Подписано в печать «___»_______2011. Формат 60*84 1/16. Бумага офсетная. Печать офсетная. Печ. л. 1,0. Тираж 100 экз. Заказ___. |
Отпечатано с готового оригинал-макета.
ЗАО «Принт-Экспресс»
197101, С.-Петербург, ул. Большая Монетная, 5 лит. А.