

Селекция яровой твердой пшеницы в среднем поволжье
На правах рукописи
МАЛЬЧИКОВ ПЕТР НИКОЛАЕВИЧ
Селекция яровой твердой пшеницы в Среднем Поволжье
06.01.05 – селекция и семеноводство
Автореферат
диссертации на соискание ученой степени
доктора сельскохозяйственных наук
Кинель - 2009
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Среднее Поволжье традиционно является регионом производства высококачественного зерна твердой пшеницы. Биоклиматический потенциал этой агроэкологической зоны позволяет стабильно выращивать зерно твердой пшеницы с содержанием белка 16-17,5%, конкурентоспособное на мировом рынке. В тоже время значительные колебания условий среды, почти ежегодное действие стрессовых факторов, видовая устойчивость к которым твердой пшеницы ниже, чем мягкой, увеличивают коммерческий риск при её возделывании. В результате в период рыночных реформ посевы твердой пшеницы в России уменьшились почти в четыре раза и стабилизировались в последние годы в пределах 0,5 млн. га. (Васильчук Н.С., 2001; Евдокимов М.Г., 2006).
Основными факторами, дестабилизирующими производство твердой пшеницы, являются засуха, высокие температуры, корневые гнили, болезни листьев, колоса и повреждение вредителями. Наряду с агротехническими средствами, эффективным и экономически целесообразным способом снижения их вредоносности считается применение устойчивых сортов. Еще П.Н.Константинов (1923) считал создание устойчивых сортов основным средством борьбы с засухой. По мнению А.А.Жученко (1988, 2008) на современном этапе развития земледелия важнейшей задачей науки является мобилизация адаптивного потенциала растений. Наличие большого внутривидового разнообразия, возможности привлечения генетической изменчивости других видов пшеницы определяют в обозримом будущем селекцию твердой пшеницы на устойчивость к абиотическим и биотическим факторам среды, продукционную способность и качество как перспективную и актуальную задачу сельскохозяйственной науки в регионе.
Цель исследований - определить эффективные пути селекции яровой твердой пшеницы и создать высокопродуктивные, устойчивые к абиотическим и биотическим стрессам, качественные сорта, приспособленные к условиям Среднего Поволжья и Урала.
Задачи исследований:
- определить факторы среды, лимитирующие продукционный процесс твердой пшеницы и оценить возможности снижения их вредоносности селекционно-генетическими средствами;
- изучить особенности формирования и функционирования ассимиляционного аппарата в онтогенезе различных по продуктивности и адаптивности сортов;
- провести изучение ростовых процессов, структуры растений и донорно-акцепторных отношений в период налива зерна различных генотипов;
- оценить взаимодействие фотосинтетических, ростовых процессов, потребления и использования азота и фосфора при формировании урожая зерна;
- выявить в коллекции и создать новые генетические источники и доноры хозяйственно-ценных признаков;
- определить наиболее целесообразные изменения признаков в процессе селекции, обосновать количественные и генетические параметры моделей сортов для Средневолжского региона;
- оптимизировать методы подбора компонентов для гибридизации и отбора в гибридных и селекционных питомниках ;
- создать высокопродуктивные, адаптированные к условиям Среднего Поволжья, высококачественные сорта и оценить экономическую эффективность их возделывания.
Научная новизна работы. Впервые в условиях Среднего Поволжья исследованы генотипические особенности формирования и функционирования ассимиляционного аппарата, донорно-акцепторных отношений в период налива зерна, ростовых процессов, потребления и использования при формировании урожая зерна азота и фосфора. Методами факторного анализа определена система взаимодействия признаков, характеризующих продукционный процесс и формирование урожая зерна. Выявлены источники и созданы новые доноры высокоэффективных генов, контролирующих элементы продуктивности, уровень гомеостаза, устойчивость к патогенам (мучнистая роса, бурая ржавчина, пыльная головня), жаростойкость процессов гаметогенеза, выполненность соломины, высоту растений, качество зерна. Идентифицированы базовые генотипы, несущие коадаптированный блок генов, обеспечивающий их приспособленность к условиям Среднего Поволжья. Определены параметры моделей сортов для Среднего Поволжья. Предложена схема оптимизации селекционного процесса с применением различных агрофонов, элементов экологической селекции и эффективных методов оценки гомеоадаптивности. Созданы новые высокоурожайные сорта: Безенчукская 200, Безенчукская степная, Безенчукская 205, Марина. Они приспособленные к условиям Нижнего, Среднего Поволжья и Урала и включены в Государственный реестр селекционных достижений России по этим регионам.
Основные положения, выносимые на защиту:
- роль признаков продукционного процесса в формировании генотипических различий по урожайности зерна
- доноры хозяйственно-ценных признаков
- модели сортов яровой твердой пшеницы
- стратегия отбора генотипов в гибридных популяциях и селекционных питомниках
- новые сорта яровой твердой пшеницы для регионов Поволжья и Урала.
Практическая значимость и реализация результатов исследований. В результате селекционно-генетических исследований предложены источники и доноры ряда признаков, модели сортов, стратегия отборов, что позволяет существенно повысить эффективность селекционного процесса и сократить сроки создания сортов.
Четыре сорта решением Государственной комиссии Российской Федерации по испытанию и охране селекционных достижений, допущены к использованию: Безенчукская 200 – с 2002 года в Средневолжском, с 2003г. – в Уральском регионах; Безенчукская степная – с 2004 г. в Нижневолжском, Средневолжском и Уральском регионах; Безенчукская 205 – с 2008г. в Нижневолжском, Средневолжском и Уральском регионах; Марина – с 2009г. в Средневолжском и Уральском регионах. Сорта Безенчукская 207 и Безенчукская Нива проходят государственные испытания с 2008 и 2009 гг. соответственно.
Апробация работы. Основные материалы диссертационной работы доложены на международных (Саратов, 1997, 2004; Казань, 2001; Краснодар 2002; Самара, 2003;), Всероссийских (СтПетербург, 1998; Москва, 2002; Пенза 2003) и региональных (Саратов 1995; Кинель 1997; Немчиновка, 1998; Пенза 2001, 2002) научно-методических, координационных и научно-практических конференциях и опубликованы в 45 работах, в том числе одна монография, получено четыре авторских свидетельства и пять патентов.
Объём и структура диссертации. Диссертационная работа изложена на 403 страницах печатного текста, содержит введение, 8 глав, выводы, предложения для практической селекции и производству; включает таблиц, рисунков. Библиографический список включает наименований, в том числе иностранных источников.
Выражаю глубокую благодарность доктору с.-х. наук А.А.Вьюшкову, кандидату с.-х. наук М.Г.Мясниковой, доктору биологических наук В.В.Сюкову, лаборантам-исследователям Л.А.Родиной, Е.П.Трофимовой за ценные советы, содействие и помощь в проведении экспериментальных работ и написании рукописи.
СОДЕРЖАНИЕ РАБОТЫ
1. УСЛОВИЯ, ОБЪЕКТЫ и МЕТОДИКА ПРОВЕДЕНИЯ ИССЛЕДОВАНИЙ
Экспериментальная часть работы выполнена на опытном поле Самарского НИИСХ в селекционном севообороте лаборатории селекции яровой твердой пшеницы.
Поля экспериментального участка расположены в области черноземной степи на территории, которая входит в центральную агроклиматическую зону Самарской области. По содержанию легкорастворимых солей, почвы относятся к почти не засоленным, за исключением карбонатно-солончаковых пятен и солонцов. Содержание гумуса варьирует в пределах 3,0-5,0%.
В годы исследований (1989-2008) условия влагообеспеченности и температурного режима сильно варьировали. В этот период наблюдались три очень сильные засухи 1995, 1998гг. (весенне-летняя) и 2001г (летняя), три сильных засухи 1996, 2005г. (весенне-летняя) и 2007 (весенняя) и две средних летних засухи 2002 и 2006гг. Относительно благоприятные для формирования урожайности условия имели место в 1990, 1992, 1993, 1994, 1997, 2000г. и 2003г.
В 1996, 1999, 2004, 2005, 2008 гг., на процессы формирования урожайности значительное влияние оказали эпифитотии листовых пятнистостей и фузариоза колоса, в 2005г. наблюдалось эпифитотия бурой ржавчины, в 1994, 2004 гг. мучнистой росы, в 2003 г. сильное полегание посевов.
Таким образом, метеорологические условия, на фоне которых проводились исследования, были контрастными и в целом включали весь спектр лимитирующих факторов среды, распространенных в регионе Среднего Поволжья. Это позволило объективно оценить исследуемый материал и всесторонне оценить перспективы создания сортов твердой пшеницы максимально адаптированных к этим условиям.
Все эксперименты в годы исследований проводились на полях селекционного севооборота Самарского НИИСХ по чистому пару и зерновому предшественнику – озимая рожь и овес на зерно (зябь). Применялась общепринятая агротехника для центральной зоны Самарской области. Посев опытных массивов, включавших делянки 25м2, проводили на глубину 6-8 см сеялкой СН-10Ц в агрегате с трактором Т-25.
Посев мелкоделяночных опытов - площадь делянки 1,0м2, 0,2м2, проводили сеялкой СКС-6-10 (кассетный вариант), сеялкой гнездового посева СПР-2 и вручную на однорядковых делянках. Сорта и гибриды, включенные в системные скрещивания (диаллельная и топкроссная схема), высевались на однорядковых делянках (2,5см х 20см) в 2-х, 3-х кратной повторности с рендомизированным размещением вариантов.
Объектами исследований были:
1. коллекционные образцы различного эколого-географического происхождения, представлявшие репрезентативную выборку из стран СНГ (Россия, Казахстан, Украина), Средиземноморья и Северной Америки, CIMMYT;
2. изогенные по выполненности соломины линии, предварительно созданные методом отбора полостебельных и выполненностебельных сибсов – потомств гетерозиготного растения от последнего беккроссного скрещивания;
3. аналоги сортов Харьковская 46, Харьковская 9 Безенчукская 139 с генеми редукции высоты растений – Rht1, Rht Anh, Rht Az;
4. гибриды F1- F4, полученные по схемам прямых диаллельных, тестерных и парных скрещиваний, гибридные популяции старших поколений;
5. сорта и селекционные линии Самарского НИИСХ.
Выполненность стебля определяли по шкале K.L. Lebsock, E.J.Koch (1968) - каждый стебель разрезали в четырех местах, оценивали выполненность средней части первого, второго, третьего, и четвертого междоузлий (сверху от колоса) в баллах: 1 балл - соломина выполнена на 20%; 2 - на 40%; 3 - на 60%; 4 - на 80%; 5 - на 100%. Сумма баллов четырех междоузлий дает индекс выполненности соломины.
Накопление и распределение биомассы по органам изучались по методическим рекомендациям НИИСХ Юго-Востока (Кумаков В.А. и др., 1982). Для этого в фазы кущения, трубкования, завершения роста соломины в длину, созревания в каждой повторности отбирали пробу растений в трех точках общей площадью 0,6м2, которая включала 80-100 растений. Площадь листьев (ПЛ) определяли расчетным методом, используя данные удельной поверхностной плотности зеленых листьев из каждой пробы и абсолютно-сухой массы зеленых листьев на 1м2. Фотосинтетический потенциал (ФП) и чистую продуктивность фотосинтеза (Ф.ч.пр.) в онтогенезе растений рассчитывали, опираясь на значения ПЛ на начало и конец периода, его продолжительности в сутках и прироста биомассы на 1м2.
Элементы структуры урожая определяли на выборке растений (50 штук), случайно отобранных из пробного снопа, взятого в трех местах делянки общей площадью 0,6м2. Пробные снопы отбирали с трех повторений. Выборка растений с повторения в мелкоделяночных опытах составляла 20-30 штук.
Для гибридологического анализа признаков отбирали растения на специально высеянных делянках в блоке с родительскими сортами и стандартом, в количестве 50-500 штук в зависимости от наличия семян и комбинации скрещивания. Гибридологический анализ признаков «содержание каротиноидов», «устойчивость растений к мучнистой росе, бурой ржавчине», «высота растений» проводили в F2 и F3 – потомствах, случайно отобранных в F2 растений.
Тип реакции растений на внедрение Puccinia recondita определяли по шкале E.B.Mains, H.S.Jackson (1926), степень поражения по R.F.Peterson et.al. (1948). Тип реакции на Blumeria graminis определяли по В.Г.Новохатко, А.Н.Борисенко (1977), степень поражения по Э.Э.Гешеле (1978). Заражение расой 17 Ustilago tritici проводили модифицированным вакуумным аппаратом В.И.Кривченко.
Стекловидность, натурная масса зерна и масса 1000 зерен определялись по общепринятым методикам (Фирсова М.Н., Попова Е.П., 1981). Содержание азота, фосфора в растительных образцах, белка, клейковины в зерне, показатель седиментации (SDS – вариант), содержание каротиноидных пигментов проводились в лаборатории зерна и массовых анализов Самарского НИИСХ по общепринятым прописям.
Однофакторный дисперсионный анализ в рендомизированных блоках проводили по Б.А.Доспехову (1979). Диаллельный анализ генетических параметров проводили по B.I.Hayman (1954, 1956). Генетический анализ количественных признаков с использованием генерационных средних (шестипараметорная модель) проводили по B.J.Hayman (1958). Параметры адаптивности, стабильности, оценивали по методикам, предложенным А.В.Кильчевским, Л.В.Хотылевой (1997), Н.А.Соболевым (1980), В.В.Хангильдиным (1976), C.S.Lin, M.R.Binns (1991), S.A.Eberhart, W.A.Russel (1966), В.А.Драгавцевым и др. (1984), G.I.Wricke (1965), С.Р.Мартынова (1990). Факторный анализ матриц генотипических корреляций проведен по методике Харман Г. (1972). Анализ коэффициентов путей Райта осуществляли по С.П.Мартынову (1978).
Оценку параметров среды, как фона для отбора, рассчитывали по методике А.В.Кильчевского, Л.В.Хотылевой (1985).
Расчеты проводились на РС АТ 486 с использованием пакета селекционно-ориентированных программ «Agros –2», разработанных под руководством докт.биол.наук С.П.Мартынова.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
2. ИСТОРИЯ СЕЛЕКЦИИ ЯРОВОЙ ТВЕРДОЙ ПШЕНИЦЫ В ПОВОЛЖЬЕ И НА УРАЛЕ
В этом разделе диссертации, основываясь на литературных данных, изложены история, методы и результаты селекции твердой пшеницы в научно-исследовательских учреждениях Поволжья и Урала – Самарском НИИСХ, НИИСХ Юго-Востока, Краснокутской СОС, Оренбургском НИИСХ, Башкирском НИИСХ, Поволжском НИИСС, Всероссийском НИИОЗ.
3. ФАКТОРЫ СРЕДЫ И ГЕНЕТИЧЕСКИЕ СИСТЕМЫ, ЛИМИТИРУЮЩИЕ УРОЖАЙНОСТЬ ЗЕРНА
Ритм внешних условий среды в Среднем Поволжье в период вегетации яровой пшеницы отличается сильным варьированием и не обнаруживает определенной закономерности, что значительно осложняет типизацию лет и селекционный процесс в целом. Эти же обстоятельства определяют многовекторность селекции и ее адаптивный характер. Поэтому очень важной предпосылкой успешной селекции является удачный выбор нескольких ключевых признаков из того множества, которое имеет перспективу улучшения (Мережко А.Ф., 2005).
Наиболее сильное отрицательное влияние на урожайность оказывают высокие температуры. Уравнение множественной линейной регрессии, описывающее зависимость урожайности сорта Харьковская 46 от температуры и осадков в период от третьей декады мая до второй декады июля включительно за 30 лет наблюдений, отчетливо это демонстрирует: Y = 118,2 – 0,58 Х1 + 0,23 Х2 – 0,84 Х3 - 0,82 Х4 – 0,57 Х5 – 2,34 Х6 - 0,006 Х7 + 0,12 Х8 – 0,04 Х9 – 0,03 Х10 – 0,03 Х11 – 0,04 Х12, где Х1 - Х6 среднедекадные температуры, а Х7 – Х12 среднедекадное количество осадков за указанный период. Наибольший отрицательный вклад вносит температура во второй декаде июня и во второй декаде июля.
Анализ коэффициентов путей Райта, характеризующих связи этих метеорологических параметров с урожайностью сорта Харьковская 46 за тот же период подтвердил определяющее влияние температурного режима на продукционный процесс и урожай зерна. Наиболее значительный прямой эффект наблюдался во взаимосвязи урожайности и температуры для двух периодов – вторая декада июня и вторая декада июля. Значимый отрицательный коэффициент корреляции между урожайностью и температурой в 3-й декаде мая обусловлен прямым эффектом температуры в этот период и косвенными эффектами температуры во второй декаде июня и второй декады июля. Следовательно негативный эффект температуры в 3-й декаде мая значительно возрастает при условии действия высоких температур в течение 2-х последующих, указанных выше декад июня и июля.
Положительный прямой эффект высоких температур в 1-й декаде июня полностью снимается косвенными отрицательными эффектами высоких температур во 2-й декаде июня и 2-й декаде июля. Другими словами сочетание повышенных температур в первой декаде июня и умеренных температур во 2-й декаде июня и июля положительно отражается на урожае. Возможно это связано с благоприятным действием повышенной температуры в 1-й декаде июня на процессы нитрификации, поставляющие в доступной форме азот растениям находящимся в этот момент стадии (трубкование) интенсивно роста. Роль осадков за изученные периоды скорее нейтральна или носит зависимый характер от температурного режима. Результаты исследований этих факторов в других регионах России позволяют сделать почти аналогичные заключения (Евдокимов М.Г.,2006; Васильчук Н.С., 2001;. Грязнов А.А., 2002; Крючков А.Г. 1997).
Таким образом, жаростойкость в селекции адаптированных к абиотическим стрессам сортов твердой пшеницы в Среднем Поволжье имеет первостепенное значение.
Методологические проблемы, которые необходимо решить для эффективной селекции на жаростойкость сводятся к определению предельных значений высоких температур, действие которых максимально дифференцирует сортовую популяцию, и подбору адекватных географических пунктов для оценки и отбора устойчивых генотипов.
Среди биотических факторов в Среднем Поволжье значительный вред урожаю твердой пшеницы наносят бурая ржавчина, мучнистая роса, листовые пятнистости и фузариоз колоса. Наибольшая вредоносность наблюдается при возникновении параллельной эпифитотии, когда патогенная популяция представляет сложный комплекс возбудителей. Такая ситуация сложилась на опытном поле Самарского НИИСХ в условиях 2005 года. Вредоносность консортной популяции патогенов сопоставима с действием высоких температур или сильной почвенной и атмосферной засухи.
Значительный вред урожаю зерна в отдельные годы могут наносить скрытостебельные вредители (шведская муха, стеблевая блоха), хлебные пилильщики, клоп черепашка (Вьюшков А.А.,2004; Глуховцев В.В, 2009) и полегание посевов.
На основе многолетних данных по урожайности стандартных сортов (Харьковская 46, Безенчукская 139, Безенчукская 182) и вредоносности биотических и абиотических стрессоров определен иерархический ряд генетических систем, несовершенство которых в наибольшей степени лимитирует стабильность продуктивности и негативно влияет на адаптивность сортов.
Актуальными направлениями в селекции на адаптивность признаны следующие (в порядке убывания значимости):
- жаростойкость процессов гаметогенеза и образования зерна;
- устойчивость ростовых процессов к обезвоживанию растительных тканей и активность корневой системы;
- устойчивость к листовым пятнистостям и корневым гнилям (Fusarium, Helminthosporium), мучнистой росе (Blumeria graminis) и бурой ржавчине (Puccinia recondita);
- устойчивость к полеганию.
Приведенное ранжирование генетических систем достаточно условно. Это связано с тем, что наиболее вредоносные факторы среды – засуха и высокие температуры воздуха при их максимальном воздействии на продукционный процесс не могут быть полностью устранены, или даже существенно уменьшены селекционными методами. В тоже время эффекты неблагоприятных условий, вызывающих полегание посевов, могут быть полностью преодолены в процессе селекции низкорослых, среднерослых и неполегающих сортов.
4. ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ФОРМИРОВАНИЯ ПРОДУКТИВНОСТИ
Эффективность селекции по оптимизации формирования фотосинтетического аппарата и структуры растений может быть достаточно высокой при наличии научно-обоснованного прогноза соответствующих изменений. Ретроспективный анализа результатов эволюции признаков в процессе селекции позволяет это делать (Кумаков В.А., 1985). В своих исследованиях мы использовали эволюционный подход, дополнив его анализом высокопродуктивных (трансгрессивных) сортов и их родителей. В некоторых случаях ограничивались анализом одного родителя - либо наиболее продуктивного, либо внесшего в родословную нового сорта основной вклад.
Интегральные признаки продукционного процесса. Сравнение проводили по группам: 1) спелости; 2) “родитель - потомок” 3) эволюционного ряда. Группы спелости формировались следующим образом: среднеранние - к-38, 214с-94, среднепоздние - Безенчукский янтарь, Гордеиформе 814, Леукурум 1690, Гордеиформе 1674. В группе “родитель- потомок”, исследованы сорта: Безенчукская 182 и его родители Харьковская 46 и Безенчукская 105; Гордеиформе 1434, Безенчукская степная и их родители – Безенчукская 182, Саратовская золотистая, Гордеиформе 740; Памяти Чеховича и родитель, внесший основной вклад (65,625 %) в его генотип – Саратовская золотистая.
Эволюционные тенденции и ретроспективный анализ провели от сорта Гордеиформе 432 до современных сортов и селекционных линий. В результате двухлетнего эксперимента выявлены преимущества над ним всех сортов по зерновой продуктивности с единицы посевной площади, выходу зерна из общей биомассы (К.хоз) и чистой продуктивности фотосинтеза. Наиболее продуктивные сорта Безенчукская 182, Безенчукская степная, Памяти Чеховича, Валентина, сочетали высокие значения общей биомассы и К.хоз. Существенное увеличение общей надземной биомассы, наблюдавшееся только у этих сортов, обусловлено возрастанием чистой продуктивности фотосинтеза (Безенчукская 182, Безенчукская степная), фотосинтетического потенциала (Валентина) и их кумулятивным эффектом (Памяти Чеховича).
Трансгрессии по зерновой продуктивности, обнаруженные в группах «родитель-потомок» были обеспечены либо возрастанием только биологического урожая надземной массы, либо одновременного повышения биомассы и К.хоз. (табл.1). Наиболее ярким проявлением трансгрессии по урожаю зерна с единицы посевной площади является сорт Безенчукская 182, полученный на основе сортов Харьковская 46 и Безенчукская 105. Преимущество по зерновой продуктивности сорта Безенчукской 182 над родителями составило 40-45 % и определялось значимым его преимуществом по биомассе и К.хоз. Различия по ФП и Ф.ч.пр. оказались недостоверными. Поскольку величина общей надземной биомассы зависит от мощности, продолжительности и продуктивности функционирования ассимиляционного аппарата, т.е. от ФП и Ф.ч.пр., то очевидно, что роль фотосинтетических показателей в формировании сортовых различий по биомассе неоднозначна по годам. В засушливом 1995 году значимое преимущество Безенчукской 182 наблюдалось по ФП, в благоприятном по увлажнению, температурному и фитосанитарному режиму 1997 году по Ф.ч.пр. В острозасушливом – с длительной весенне-летней засухой 1998 году достоверных различий между сортами по ФП и Ф.ч.пр. не наблюдалось, хотя абсолютные значения у Безенчукской 182 были несколько больше, чем у родительских сортов.
1. Основные показатели продуктивности и фотосинтетической деятельности сортов в группах “родитель-потомок”
| Сорт | Год Райони- -рования | Показатели продуктивности и фотосинтетической деятельности, в % к одному из родительских сортов | ||||
| Убиол. | Ухоз. | Кхоз. | ФП | Ф.ч.пр. | ||
| I. Сорта: Харьковская 46, Безенчукская 105, Безенчукская 182* (в среднем за 1995, 1997, 1998, 2000, 2001 гг.) | ||||||
| Харьковская 46 | 1963 | 100,0 | 100,0 | 100,0 | 100,0 | 100,0 |
| Безенчукская 105 | 1965 | 100,1 | 105,5 | 102,1 | 102,1 | 97,7 |
| Безенчукская 182 | 1993 | 121,1 | 145,7 | 124,0 | 111,8 | 108,3 |
| НСР0,05 | 20,7 | 35,4 | 15,3 | Ff <Ft | Ff <Ft | |
| II. Cорта: Безенчукская 182, Саратовская золотистая, Гордеиформе 740, Гордеиформе 1434 (в среднем за 1997, 1998, 2000 гг.) | ||||||
| Безенчукская 182 | 1993 | 100,0 | 100,0 | 100,0 | 100,0 | 100,0 |
| Саратовская золотистая | 1993 | 79,7 | 68,2 | 86,6 | 90,4 | 93,1 |
| Гордеиформе 740 | - | 74,0 | 67,1 | 84,4 | 95,4 | 87,8 |
| Гордеиформе 1434 | - | 111,7 | 113,9 | 101,4 | 111,9 | 102,4 |
| НСР0,05 | 10,3 | 10,1 | 12,1 | 13,1 | 11,1 | |
| III. Cорта: Безенчукская 182, Саратовская золотистая, Гордеиформе 740, Безенчукская степная (в среднем за 2000, 2001 гг.) | ||||||
| Безенчукская 182 | 1993 | 100,0 | 100,0 | 100,0 | 100,0 | 100,0 |
| Саратовская золотистая | 1993 | 90,3 | 76,6 | 84,0 | 78,7 | 114,6 |
| Гордеиформе 740 | - | 99,6 | 86,6 | 91,0 | 98,9 | 100,7 |
| Безенчукская степная | 2004 | 113,6 | 115,3 | 103,2 | 111,2 | 102,1 |
| НСР0,05 | 15,9 | 12,1 | 9,0 | 11,5 | 13,5 | |
| IV.Сорта: Саратовская золотистая, Памяти Чеховича (в среднем за 2000, 2001гг.) | ||||||
| Сарат.золотистая | 1993 | 100,0 | 100,0 | 100,0 | 100,0 | 100,0 |
| Памяти Чеховича | - | 115,3 | 138,2 | 121,2 | 144,4 | 79,9 |
| НСР0,05 | 15,9 | 12,1 | 9,0 | 11,5 | 13,5 | |
Примечание * жирным шрифтом выделены сорта потомки
В 2000 году - среднем по уровню влияния условий среды на процессы формирования урожая, сорт Безенчукская 182 достоверно превосходил по ФП родительский сорт Безенчукская 105, достоверных различий с другим родителем Харьковской 46 по этим признакам не обнаружено. В 2001 году при благоприятных условиях в период вегетативного роста и засухе во второй половине вегетации наблюдалось значимое преимущество сорта Безенчукская 182 над родительскими сортами по ФП.
Сорта Гордеиформе 1434 и Безенчукская степная превосходили лучшего родителя – сорт Безенчукская 182 по урожайности зерна на 13-15 %, что значительно меньше, чем в предыдущем случае, где сорт Безенчукская 182 сам рассматривался как результат трансгрессии. Тем не менее, сорт Гордеиформе 1434 довольно отчетливо проявил тенденции усиления ФП в 1997 г., Ф.ч.пр. в 1998г., Ф.ч.пр. и К.хоз. в 2000 году. Безенчукская степная превосходила Безенчукскую 182 в 2000 г. по Ф.ч.пр. и К.хоз., в 2001 г. по ФП.
Главное преимущество сорта Памяти Чеховича над сортом Саратовская золотистая – внесшем основной вклад в его генотип, заключается в значительно более высоком выходе зерна из общего урожая надземной массы – К.хоз.
Таким образом, если рассматривать I и IV группы сортов, где трансгрессия по урожаю зерна 38,2-40,2 % то легко обнаружить значительный вклад в эту величину признака К.хоз. В то же время ФП и Ф.ч.пр. имеют неоднозначный эффект и их роль в формировании зерновой продуктивности зависит от условий года. Однако в большинстве случаев более продуктивные (трансгрессивные сорта) превосходят по какому-либо из этих показателей лучший родительский сорт, что можно считать результатом повышения пластичности компонентов фотосинтетической деятельности в процессе селекции. При этом наблюдается определенная закономерность в изменчивости признаков у сортов в зависимости от условий года. В благоприятных для ростовых процессов условиях более продуктивные сорта при равных значениях ФП с низкоурожайными сортами включают (на уровне ценоза, растения, листа или хлоропласта) механизмы активной ассимиляции и имеют в целом за вегетацию более высокие значения Ф.ч.пр. В условиях засухи продуктивные сорта выделяются по ФП.
Сильное влияние на продукционный процесс твердой пшеницы оказывает продолжительность вегетации сорта. Здесь выявлена четкая зависимость изменений фотосинтетических параметров от скороспелых до среднепоздних сортов. Оптимальный баланс в соотношении ФП и Ф.ч.пр. и максимальные значения К.хоз. характерны для группы среднеспелых сортов. Сокращение периода всходы – колошение на 5-7 дней и всего вегетационного периода на 4-6 дней сопровождается значимым увеличением показателей Ф.ч.пр. и заметным снижением ФП и К.хоз. Последовательное увеличение периода всходы – колошение на 3-4 и 5-6 дней приводит к снижению Ф.ч.пр. и К.хоз и увеличению ФП. Значение имеет то обстоятельство, что сорта, более поздние по продолжительности периода до колошения и в целом всей вегетации, не отличаются по урожаю зерна от сорта Безенчукская 182. Это оставляет надежду на результативность селекционного улучшения сортов среднепозднего биотипа.
Высокий уровень пластичности интегральных признаков продукционного процесса у современных сортов во многом объясняется особенностями их формирования в онтогенезе в зависимости от условий среды.
Формирование листовой поверхности. За 9 лет измерения площади листьев (1995-2004гг) в течение 3-х лет максимальный листовой индекс в посевах сорта Харьковская 46 превышал 4,0 м2/м2, что можно рассматривать как способность сортов этой группы спелости в Среднем Поволжье при благоприятных условиях с достаточно высокой частотой формировать оптимальную площадь листьев (Ничипорович А.А., 1977). Однако этот вывод не противоречит возможности и целесообразности селекционного улучшения этого признака. Объектом селекционного воздействия в данном случае будет динамика формирования ПЛ в онтогенезе и в зависимости от условий среды. Особенности этого процесса в предшествующий период ярко проявились при изучении сортов в острозасушливые (1995, 1998) и благоприятные (1997, 2001) годы.
В 1995 году высокопродуктивный и засухоустойчивый сорт Безенчукская 182 превысил сорт Харьковская 46 по биологическому урожаю на 37,2 %, урожаю зерна на 87,8% и К.хоз на 34,7%. Почти такое же преимущество наблюдалось и над сортом Безенчукская 105. В основе этого превосходства, как уже было отмечено выше, стало значительное увеличение у Безенчукской 182 фотосинтетического потенциала. Анализ динамики листовой поверхности в онтогенезе сортов показывает, что основное преимущество более продуктивного сорта приходится на период трубкование – цветение. Причем это преимущество по ЛП не пропорционально распределяется между главным и боковыми побегами (рис.1).


Рис. 1. Динамика листовой поверхности главных и боковых побегов на 1м2 посева в онтогенезе сортов твердой пшеницы в условиях 1995 года.
Доминирующая роль главного побега в формировании ассимиляционного аппарата листьев у сорта Безенчукская 182 на протяжении всего периода их роста очевидна. Большая по абсолютной величине листовая поверхность боковых побегов сорта Харьковская 46 не в состоянии компенсировать относительно слабое развитие листьев главных побегов. Более того, сама модель формирования ассимиляционного аппарата путем резкого увеличения абсолютного и относительного вклада листьев боковых побегов, начиная с момента кущения оказалась в условиях засухи 1995 года неэффективной – различия между сортами по урожаю зерна боковых побегов носили случайный характер и были незначительны по величине - 1,9, 2,2, 2,7 г/м2 для Харьковской 46, Безенчукской 182 и Безенчукской 105 соответственно.
В еще более жестких условиях засухи 1998 года преимущество продуктивных сортов над Харьковской 46 по ПЛ главных побегов в период трубкование-цветение подтвердилось. При этом почти такое же превосходство в динамике листовой поверхности над Харьковской 46 показал малопродуктивный сорт Безенчукская 105. Его высокая засухоустойчивость в период вегетативного роста утрачивается при формировании и наливе зерна.
Таким образом, усиленный рост листовой поверхности главного побега от трубкования до формирования зерна в условиях сильной засухи является лишь предпосылкой для относительного преимущества сорта по урожаю зерна. Он в процессе рекомбиногенеза должен быть комплементарно дополнен повышенной способностью колоса к аттракции и реализации накопленного фотосинтетического потенциала в период налива зерна.
В условиях 1997 года - самого благоприятного для роста и продуктивности отмечена положительная связь площади листьев главного побега на 1 м2 в период от трубкования до колошения с урожайностью зерна и основными элементами ее структуры. Наиболее высокие и значимые коэффициенты корреляции площади листьев главного побега в колошение, наблюдались с числом зерен на растении, К.хоз и урожаем зерна (0,65; 0,69; 0,62 соответственно). Аналогичной зависимости листовой поверхности боковых побегов и урожайности не обнаружено. Несмотря на значительный вклад боковых побегов, различия по урожайности между сортами формировались исключительно благодаря изменчивости продукционного процесса главных побегов. В 1997 году имело положительное значение увеличение листовой поверхности главных побегов в период формирования числа зерен в колосе - решающего компонента межсортового варьирования урожайности при минимальных воздействиях лимитирующих факторов в период налива зерна.
В условиях 2001 года сорт Безенчукская 182 превосходил по листовой поверхности главных побегов Харьковскую 46 в течение всего периода вегетации, но почти не отличался от Безенчукской 105 и даже уступал стародавнему сорту Гордеиформе 432. Особенность сорта Гордеиформе 432 – накапливать исключительно большую листовую массу главного побега – заметила еще И.В.Красовская (1946). Современные высокоурожайные сорта: Безенчукская степная, Памяти Чеховича, Валентина и Безенчукская 182 мало отличались по динамике листовой поверхности в благоприятных для роста условиях, сложившихся в 2001 году, от сорта Гордеиформе 432. Различия сортов по площади листьев обнаружились в период цветения - созревания.
Возможно, именно эта особенность современных сортов позволяет им формировать более крупное зерно. В тоже время преимущество по урожайности зерна обеспечивается у них не только крупнозерностью, но и числом зерен на 1м2, числом зерен в колосе и К.хоз., т.е. процессами морфогенеза. Эти признаки в условиях 2001 г. не зависели от величины листовой поверхности – соответствующие коэффициенты корреляции в период от кущения до цветения были незначимы (табл.2).
2. Коэффициенты генотипической корреляции площади листьев в онтогенезе с урожайностью зерна и ее элементами, 2001 г.
| Площадь листьев по фазам онтогенеза | Урожай зерна, г/м2 | Число растений на 1м2 | Число зерен на 1м2 | Число зерен в колосе | Масса 1000 зерен, г. | К.хоз., % |
| Кущение | -0,07 | -0,50 | -0,13 | 0,45 | 0,07 | 0,15 |
| Трубкование | -0,15 | -0,18 | -0,18 | 0,14 | -0,08 | -0,41 |
| Цветение | 0,58 | 0,22 | 0,58 | 0,47 | 0,34 | 0,41 |
Продолжительность жизни листьев. Базовой величиной этого показателя является продолжительность вегетационного периода. Обычно более длительный период ассимиляции наблюдается у позднеспелых сортов, что не всегда положительно отражается на урожае зерна. Значение как селекционный признак имеет увеличение продолжительности функционирования ассимиляционного аппарата без увеличения общей продолжительности вегетации (Кумаков В.А., 1971). Увеличение продолжительности жизни листьев без удлинения вегетации сортов может определяться активными и продолжительно функционирующими меристемами, т.е. гормональным статусом, корректирующим старение растительного организма. В репродуктивный период у зерновых культур, когда стебель завершает рост, остается два акцептора ассимилятов – колос и корневая система. Постепенно влияние колоса на фотосинтетический аппарат становится определяющим. Вместе с тем продолжительность жизни листьев зависит от засухоустойчивости и развития корневой системы (Игошин А.П., 1984; Woodword R.G., Rawson H.M., 1976; Кумаков В.А.,1985; Чиков В.И., 1987).
Взаимосвязь продолжительности жизни 2-х верхних листьев с числом зерен в колосе, колоске и засухоустойчивостью, в условиях засухи 1998 года, была средней для всех признаков - 0,31; 0,31; 0,45 соответственно. В условиях 2001 года - благоприятных для роста листьев и засушливых для налива зерна эта взаимосвязь была - средней для числа зерен в колосе и колоске - 0,47; 0,57, и довольно тесной (0,76) для засухоустойчивости. Положительная связь числа зерен в колосе и колоске с продолжительностью жизни листьев в оба года свидетельствуют о сложном взаимодействии этих признаков с засухоустойчивостью в процессе совместного влияния на фотосинтетический аппарат. Этот результат объясняется селекцией на высокую продуктивность в условиях неустойчивого увлажнения. В процессе селекции улучшились параметры продуктивности колоса и засухоустойчивости современных сортов. В условиях сильнейшей засухи 1998 года сорта Гордеиформе 1434 и Гордеиформе 814 имели самую длительную продолжительность жизни листьев верхнего яруса. В то же время сорта Саратовская золотистая и к-38, отличаясь высокой засухоустойчивостью, не выделяются в такой же степени по продолжительности жизни листьев. Сорта Памяти Чеховича, Безенчукская степная, Безенчукская 182 обладают хорошей засухоустойчивостью и имеют высокую озерненность колоска. Эти особенности позволяют им при широком варьировании условий среды, эффективно поддерживать в активном состоянии необходимую ассимиляционную поверхность листьев.
Чистая продуктивность фотосинтеза. В.Г.Конарев (2001), отметив эволюционный консерватизм молекулярных механизмов фотосинтезирующих систем хлоропласта, подчеркнул отсутствие сколь-либо существенного эффекта от попыток воздействовать на них с целью селекционного улучшения. Обсудив возможные пути модернизации молекулы рибулезобифосфаткарбоксилазы (поиск субъединиц с высокой карбоксилазной и низкой оксигеназной активностью) и перспективы переключения растений С3 – типа фотосинтеза на С4 – тип, он пришел к заключению, что «реален и наиболее актуален в решении проблемы углеродного обмена поиск путей реализации больших, по сути неограниченных, возможностей фотосинтеза в формообразовательных процессах, направленных на улучшение индекса урожайности и получение максимального выхода продукции». В этой связи очень важно правильное толкование причин тех сортовых различий на уровне агроценоза, которые довольно часто наблюдаются по Ф.чпр. и интенсивности фотосинтеза.
Как уже отмечено выше роль признака Ф.ч.пр. в формировании биологического и хозяйственного урожая, изученных сортов твердой пшеницы, неоднозначна. Все современные сорта превышают старый сорт Гордеиформе 432 по Ф.ч.пр. в целом за вегетацию и в зависимости от условий среды меняют ранги относительно друг друга. Сравнение динамики Ф.ч.пр в онтогенезе сортов Безенчукская 182 и Гордеиформе 432 (рис.2), отчетливо показывает, что основное преимущество Безенчукской 182 по этому показателю формируется в период от цветения до созревания. 
Рис.2. Динамика Ф.ч.пр. в онтогенезе сортов Безенчукская 182 и Гордеиформе 432
Отсутствие различий между сортами в период трубкование – цветение, объясняется интенсивным ростом соломины в этот период, масса которой у низкопродуктивного сорта Гордеиформе 432 в благоприятных условиях достигала максимальных величин среди изучавшихся сортов.
Анализ динамики Ф.ч.пр. в онтогенезе сортов Харьковская 46, Безенчукская 105 и Безенчукская 182 на основе данных многолетнего эксперимента показывает, что достоверные различия между минимальным и максимальным значением наблюдается в репродуктивный период.
Во всех случаях определения этого признака в момент кущения не обнаружено сортовых различий. Даже сравнение сортов - скороспелых и позднеспелых, с разным вкладом в общую листовую поверхность боковых побегов, не дает значимых различий при расчете величины чистой продуктивности на всю поверхность листьев. Эти различия становились существенными только в период налива зерна.
Таким образом, ростовые и эпигенетические процессы в данном случае, видимо, контролируют фотосинтетическую функцию и формирование урожая, хотя при определенных условиях урожайность может лимитироваться и фотосинтезом (Мокроносов А.Т., 1982).
В целом необходимо констатировать, что работы по целенаправленному поиску форм с повышенной фотосинтетической активностью необходимо проводить и в случае их обнаружения использовать как исходный селекционный материал (В.А.Кумаков, 1985).
Ростовые процессы, структура растения и донорно-акцепторные отношения. Взаимодействие источника и акцептора ассимилянтов рассматривается как основной фактор детерминации фотосинтеза на организменном уровне (Курсанов А.Л., 1982; Мокроносов А.Т., 1981; 1982; 1983). Ограничивающим может быть и тот, и другой компонент. Предполагается, что лучшее понимание этих процессов и приложение к селекционно-растениеводческой практике приведет к новому скачку в урожайности (Davies D.R., 1977; Кумаков В.А., 1980, 1982).
Очевидно, что относительная активность метаболических процессов органов (рост, дыхание) определяет доминирующий вектор в потоке ассимилятов, начиная от хлоропласта. Бесспорным является участие в этом процессе гормонов ауксиновой и цитокининовой природы (Мокроносов А.Т., 1983; Дамиш В.,1983;Чиков В.И., 1987; Киризий Д.А.,1995). Также необходимо учитывать возможности растений в процессе роста и развития переключать связи донор – акцептор между отдельными органами и изменять соотношения экспортно-импортной функции самих органов (Киризий Д.А.,1995). Ряд исследователей из CIMMYT, США, Австралии, используя для анализа специальных экспериментов многомерные регрессионные уравнения, пришли к заключению, что улучшение баланса между донором и акцептором в процессе селекции – эффективный подход в повышении урожая надземной биомассы и КПД ФАР (Reyndolds M.P., Pietragalla J. et al., 2008). Задача состоит в том, чтобы определить, чем ограничивается накопление сухого вещества на каждом конкретном этапе развития – донором или акцептором (акцепторами). По мнению А.Т.Мокроносова (1982) хлоропласт способен длительное время работать в режиме гиперфункции, что особенно важно для перспектив создания крупноколосых форм пшеницы, имеющих широкое отношение колос / листья в момент цветения. В то же время необходимо учитывать, что высокая эффективность фотосинтеза в начале вегетации приводит к образованию дополнительных листьев и побегов, что не всегда целесообразно.
В связи с этим в своих исследованиях мы стремились определить связь сортовых различий по интенсивности ростовых процессов в онтогенезе с продуктивностью и оценить перспективы их оптимизации в процессе селекции.
Накопление и распределение биомассы в период от всходов до колошения. Величина общей биомассы характеризует интенсивность синтетических процессов агроценоза на единице площади, функции роста органов описывают векторные «приоритеты» в ростовых процессах генотипов. При образовании сухого вещества надземной массы решающее значение имеет не ее количество, а динамика образования. Генотипические особенности начинают проявляться на самых ранних этапах развития и наблюдаются в течение всего онтогенеза на уровне синтетических, физиолого-биохимических и эпигенетических процессов, которые затем могут трансформироваться в сортовые различия по урожайности (Вркоч Ф., 1984).
В таблице 3 приведены данные за шесть лет изучения трех сортов в период от всходов до кущения, которые позволили довольно четко определить сортовые «приоритеты» в ростовых процессах в этот период и идентифицировать следующие два биотипа твердой пшеницы. Первый биотип характеризуется значимым превалированием в ростовых процессах боковых побегов с момента начала их роста и независимо от конкретной динамики лимитирующих факторов среды. Представителем этого биотипа является сорт Харьковская 46. Второй биотип, представленный сортами Безенчукская 105 и Безенчукская 182, характеризуется доминированием в ростовых процессах главного побега.
3. Накопление биомассы и распределение ее по органам в период от всходов до кущения, в среднем за: 1995, 1997, 1998, 2000, 2001, 2003 годы
| Сорт | Биомасса, г/м2 | Функции роста органов растений, % | |||
| общая | главных побегов | стебли главных побегов | листья главных побегов | Боковые побеги | |
| Харьковская 46 | 50,7 | 29,1 | 19,6 | 44,3 | 36,1 |
| Безенчукская 105 | 47,8 | 34,0 | 25,2 | 50,3 | 24,5 |
| Безенчукская 182 | 48,6 | 35,4 | 25,9 | 51,7 | 22,4 |
| НСР0,05 | Ff < Ft | Ff < Ft | 3,3 | 3,1 | 4,6 |
| m, % | 2,57 | 5,39 | 4,51 | 2,01 | 5,29 |
Таким образом, к моменту кущения у представителей разных биотипов в ростовых процессах определяются доминирующие векторы. К первому биотипу, кроме сорта Харьковской 46, отнесены - Безенчукский янтарь и селекционные линии Гордеиформе 814 и Гордеиформе 1674. Перспективы распространения сортов этого биотипа имеются в регионах с прохладной весной и осадками во второй половине вегетации (Западная Сибирь, Алтай, Оренбургская область). Безенчукский янтарь включен в госреестр сортов России с допуском к использованию по 9 региону, где наблюдается именно такой ритм варьирования погодных условий.
Большинство изученных генотипов в условиях Среднего Поволжья отличаются достаточно сильным доминированием в ростовых процессах главных побегов. К этой группе отнесены сорта Безенчукская 105, Гордеиформе 432, Безенчукская 182, Безенчукская 139, Безенчукская степная, Памяти Чеховича, Валентина, Саратовская золотистая, Степь 3, Саратовская 59, Безенчукская 205, Безенчукская 200, селекционные линии Гордеиформе 740, Гордеиформе 1434, Леукурум 1690. Сорта этой группы достаточно легко отделить от первой группы в период кущения. Сорта Безенчукская степная и Памяти Чеховича имеют относительно более жесткое доминирование главного побега в момент кущения, которые в очень благоприятных для кущения условиях 2001 и 2003 годов не превысили 30% рубеж в доле боковых побегов. Посевы засухоустойчивых сортов обычно имеют уменьшенную листовую поверхность в начале вегетации, за счет этого сохраняется больше запасов влаги в почве к цветению и эмбриональному росту зерна чрезвычайно чувствительному к недостатку влаги (Ничипорович А.А., 1972; Richards R.A., 1983). Это позволяет удерживать приемлемые величины К.хоз. и урожая зерна (Richards R.A., Townley –Smith T.F., 1987). В данном случае необходимый эффект достигается за счет жесткого ограничения роста боковых побегов осуществляемого на генетическом уровне.
Динамика общей надземной биомассы на 1 м2 в течение онтогенеза в среднем за ряд лет отчетливо демонстрирует превосходство высокопродуктивных сортов по приростам сухого вещества в период налива зерна. После момента цветения весь прирост надземной биомассы аккумулируется в зерне, основные генотипические различия по урожайности зерна формируются в зависимости от интенсивности роста именно в этот период. Тем не менее, факторы, определяющие эти различия в росте, связаны с морфогенетическими процессами, проходящими в период от всходов до цветения. В этот период формируется колос, определяются его размеры, акцепторная емкость и аттрагирующая сила. К моменту окончания роста стебля в длину (цветение) наиболее урожайные сорта имели значительно более высокую потенциальную продуктивность колоса, выраженную величиной его «стартовой» массы, чем сорта низкоурожайные и принадлежащие к ранним периодам селекции (табл.4).
4. Распределение сухой массы по органам главного побега в цветение
| Сорт | «Стартовая» масса колоса, мг | Доля органов в общей сухой массе побега, % | ||
| листовые пластинки | соломина с листовыми влагалищами | колос | ||
| В среднем за 1995, 1997, 1998, 2000, 2001, 2003 годы | ||||
| Харьковская 46 | 194,2 | 21,8 | 58,6 | 19,6 |
| Безенчукская 182 | 282,8 | 21,5 | 55,5 | 23,0 |
| Безенчукская 105 | 255,7 | 22,0 | 57,7 | 20,3 |
| НСР0,05 | 53,0 | Ff < Ft | Ff < Ft | 1,96 |
| m, % | 6,53 | 1,97 | 1,33 | 2,87 |
| В среднем за 1997, 1998, 2000 годы | ||||
| Харьковская 46 | 185,3 | 20,1 | 61,1 | 18,8 |
| Безенчукская 182 | 251,6 | 19,6 | 58,0 | 22,5 |
| Безенчукская 105 | 221,6 | 20,6 | 59,9 | 19,4 |
| Гордеиформе 1434 | 283,2 | 19,3 | 57,4 | 23,3 |
| Саратовская золотистая | 201,4 | 19,7 | 61,3 | 19,0 |
| Гордеиформе 740 | 244,4 | 19,1 | 61,5 | 19,3 |
| Безенчукский янтарь | 203,5 | 18,5 | 61,5 | 20,0 |
| Безенчукская 200 | 226,9 | 19,6 | 59,9 | 20,5 |
| Гордеиформе 814 | 215,6 | 19,9 | 60,9 | 19,3 |
| НСР0,05 | 45,3 | Ff < Ft | Ff < Ft | 2,59 |
| В среднем за 2000, 2001 годы | ||||
| Гордеиформе 432 | 268,1 | 17,9 | 63,6 | 18,5 |
| Харьковская 46 | 193,5 | 17,8 | 64,4 | 17,8 |
| Саратовская золотистая | 261,2 | 18,6 | 60,8 | 20,6 |
| Безенчукская 182 | 311,3 | 17,2 | 62,0 | 20,9 |
| Гордеиформе 1674 | 332,0 | 15,7 | 63,2 | 21,1 |
| Безенчукская степная | 319,6 | 20,0 | 59,3 | 20,7 |
| Валентина | 360,1 | 17,8 | 61,7 | 20,5 |
| Памяти Чеховича | 320,7 | 18,3 | 59,5 | 22,2 |
| НСР0,05 | 41,9 | 1,66 | 1,82 | 1,25 |
| m, % | 4,94 | 3,02 | 0,93 | 1,96 |
Напомним, что под «стартовой» массой понимается масса колоса в момент окончания роста стебля. Кроме того, у высокопродуктивных генотипов в момент цветения отмечено более выгодное для колоса распределение сухого вещества между колосом и вегетативной частью побега. Причем увеличение доли колоса проходило как за счет доли стебля, так и листьев. Доминирующим у среднерослых сортов (Памяти Чеховича, Безенчукская степная) был процесс снижения доли стебля. Среди высокорослых сортов наиболее высокая доля колоса в фазу цветения наблюдалась у сортов Безенчукская 182 и Гордеиформе 1434, характерной чертой которых является повышенная озерненность колоса и колоска.
Следовательно одновременное снижение высоты растений и увеличение озернённости колоса может привести в процессе селекции к получению крупноколосых с высоким потенциалом продуктивности сортов.
Донорно-акцепторные отношения между органами в период налива зерна. Итак, увеличение потенциала колоса может создать проблемы в период формирования и налива зерна, если модифицированные сорта не будут располагать соответствующим фондом ассимилянтов. Очевидно, что вероятность развития такого сценария, будет больше, если уже существующие сорта с различным уровнем напряженности в системе донор – акцептор в период налива зерна проявляют недостаточность в реализации потенциала колоса. Для поиска ответа на вопрос, что лимитирует урожайность в репродуктивную фазу развития - синтетические возможности или емкость запасающих органов, В.А.Кумаков и др. (1982, 2000) предложили методику количественного измерения и оценки этих процессов в зависимости от условий среды и генотипа. Эта методика позволяет на основе баланса сухого вещества побега, коэффициента реализации продуктивности колоса (КРК), степени востребованности структурных ассимилянтов для налива зерна, проводить ранжирование сортов по их адекватности агроэкологическим условиям и определять в каждом конкретном эпизоде перспективность тех или иных преобразований селекционного материала. Рассмотрим данные, полученные на сортах Безенчукская 105, Харьковская 46, Безенчукская 182 в 1995, 1997, 1998 и 2001 годы, два из которых были острозасушливыми (1995,1998) и два (1997, 2001) благоприятными по гидротермическому режиму. Близкими по величине в контрастных условиях среды были отношения сухой массы колоса и вегетативных органов главного побега. Значительные различия наблюдались по отношению масс колоса и листьев (табл.5). В отличие от данных, приведенных В.А.Кумаковым (1985) и относящихся к мягкой пшенице, в нашем эксперименте засуха не только не приводила к увеличению отношения масс колоса и листьев, а даже значительно уменьшала его. Возможно, это связано с различной удельной поверхностной плотностью листьев разных видов пшеницы, которая сильно зависит от фазы развития, возможностей эвакуации ассимилянтов из листьев, среды и генотипа.
5. Соотношение масс колоса, листьев и вегетативных органов в цветение
| Сорт | 1995,1998гг. | 1997, 2001гг. | Среднее за 4 года | |||
| к/л | к/во | к/л | к/во | к/л | к/во | |
| Харьковская 46 | 0,73 | 0,26 | 1,27 | 0,27 | 1,00 | 0,26 |
| Безенчукская 182 | 0,85 | 0,32 | 1,49 | 0,32 | 1,17 | 0,32 |
| Безенчукская 105 | 0,64 | 0,24 | 1,48 | 0,30 | 1,05 | 0,27 |
| Среднее | 0,74 | 0,27 | 1,41 | 0,30 | ||
Сокращения: к- колос, л-листья, во-вегетативные органы
Итак, засушливые условия не вызывают у твердой пшеницы увеличения отношений колос/ листья и колос/ вегетативная масса в целом. В среднем за четыре года более широкими эти соотношения были у сорта Безенчукская 182. Следовательно, можно предположить, что нагрузка на единицу ассимиляционного аппарата в период налива зерна у этого сорта выше, а вероятность включения механизмов вторичного использования структурных веществ вегетативной сферы (реутилизации) больше. В условиях засухи можно ожидать проявления недостаточности общего фонда ассимилянтов для реализации потенциала продуктивности колоса, по которому Безенчукская 182 значительно превосходит Харьковскую 46 и Безенчукскую 105. Анализ динамики изменения сухой массы главного побега за период цветение – восковая спелость показывает, что это не так. Сорт Безенчукская 182 во всех ситуациях выделяется по приросту массы колоса, т.е. имеет самый высокий КРК и не обнаруживает относительной недостаточности фонда ассимилятов в период налива зерна.
Даже в условиях засухи (1995,1998) сорт Безенчукская 182 заметно опережал Харьковскую 46 и Безенчукскую 105 по приростам биомассы в период от цветения до восковой спелости и коэффициенту реализации колоса (табл.6). В 1998 году у этого сорта на фоне интенсивной реутилизации, наблюдавшейся у сортов Харьковская 46 и Безенчукская 105, отмечен значительный прирост биомассы главного побега.
6. Изменение сухой массы главного побега сортов яровой твердой пшеницы за период цветение – восковая спелость в засушливые годы
| Сорт | Прирост сухой массы | Изменение массы вегетативных органов | ||||
| всего побега, мг | в том числе колоса (КРК) | мг | % к исходной массе в цветение | |||
| мг | % к стартовой массе | |||||
| 1995 год | ||||||
| Харьковская 46 | 29,5 | 165,2 | 220,2 | -136,5 | -21,1 | |
| Безенчукская 182 | 287,9 | 359,0 | 291,7 | -54,5 | -7,0 | |
| Безенчукская 105 | 28,2 | 196,0 | 221,1 | -178,1 | -22,7 | |
| 1998 год | ||||||
| Харьковская 46 | 28,1 | 139,4 | 220,7 | -77,4 | -12,9 | |
| Безенчукская 182 | 325,9 | 316,7 | 328,5 | +94,8 | +16,1 | |
| Безенчукская 105 | -57,9 | 126,9 | 210,7 | -139,5 | -21,4 | |
В благоприятных условиях 1997 и 2001годов «стартовая» масса колоса незначительно варьировала по годам – 405,1 мг и 410,7 мг соответственно в среднем по всем трем сортам. КРК в 2001 году был меньше, чем в 1997 году на 10,0 % в относительных величинах. При этом реутилизация наблюдалась только у сорта Безенчукская 105. У сортов Харьковская 46 и Безенчукская 182, напротив отмечен значительный прирост биомассы вегетативных органов главного побега. Этот факт следует рассматривать, как проявление высоких потенциальных возможностей фотосинтеза и в то же время недостаточной аттрагирующей активности колоса в сложившихся условиях, которая объясняется значительной стерилизацией цветков высокими температурами в период цветения – формирования зерна.
Сорт Гордеиформе 1434 при более крупном колосе и побеге в целом, унаследовал от Безенчукской 182 те же пропорции между колосом и вегетативными частями растения. Практически одинаковыми были у этих сортов КРК и источники ассимилянтов в период налива зерна в 1997 году. В условиях засухи 1998 года Гордеиформе 1434 при большем числе зерен в колосе, используя возможности реутилизации, тем не менее, снизил относительно Безенчукской 182 КРК и общий прирост побега от цветения к восковой спелости. Возможно, что это связано с различиями по вегетационному периоду – Гордеиформе 1434 колосится и созревает на 1-2 дня позднее.
Итак, селекция на повышение потенциала продуктивности колоса, при одновременном увеличении функции его роста к моменту цветения и акцепторной нагрузки на фотосинтетический аппарат в период налива зерна, не приводит к отрицательным последствиям для его реализации, если сопровождается ростом общей адаптивности и засухоустойчивости в частности. Современные, продуктивные сорта к моменту цветения при той же или даже сниженной массе соломины формируют значительно более крупный колос, чем низкопродуктивные сорта и сорта предыдущих этапов селекции. Ранги сортов по соотношению колос / соломина в цветение, почти не меняются и восковую спелость зерна рис.5, 6.
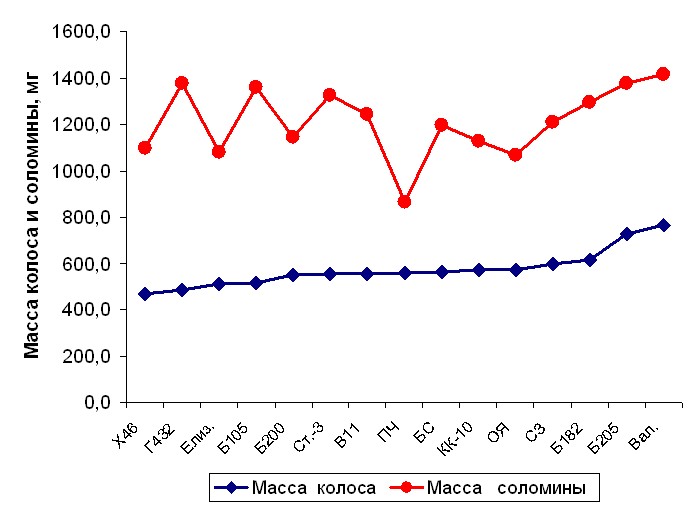
Рис.5. Зависимость массы колоса от массы соломины различных сортов твердой пшеницы в момент цветения, чистый пар, 2004 год.

Рис. 6. Зависимость массы колоса от массы соломины различных сортов твердой пшеницы в момент созревания, чистый пар, 2004 год.
Сокращения: Х-Харьковская, Г-Гордеиформе, Б- Безенчукская, КК- Краснокутка, ОЯ- Омская янтарная, СЗ – Саратовская золотистая, ПЧ- Памяти Чеховича, Елиз.-Елизаветинская, Ст.- Степь 3, В –Воронежская, С-степная, Вал. – Валентина.
Широкое отношение масс колос / соломина, наблюдалось для среднерослых сортов Памяти Чеховича и Омская янтарная, как в цветение, так и в момент созревания. Учитывая, что эти сорта имеют сокращенный на 2-4 дня период всходы – колошение, когда формируется потенциал продуктивности колоса, дальнейшая эволюция селекционного материала на их основе по пути оптимизации параметров вегетационного периода и увеличения продуктивности колоса вполне реальная перспектива.
Динамика потребления и использования на формирование урожая минеральных элементов. Наиболее типичной ситуацией в полевых посевах Поволжья и Урала при обилии солнечного света является дефицит влаги и минеральных веществ, особенно азота (Колосков П.И., 1971, Абдрашитов Р.Х., 2003). Азот дефицитен во всех почвах России, включая черноземные (Мишустин Е.Н. и др., 1983). Поэтому в этих условиях необходимо ясное представление того, каким образом модифицировать сортовые популяции, чтобы посевы наиболее эффективно использовали имеющиеся ресурсы местообитания для повышения хозяйственного урожая. На Западе интенсивно разрабатывается идея отбора сортов типа ”low input varieties” – т.е. сортов с низким потреблением, не требующих при возделывании интенсивного вложения капитала и дающих максимальную прибыль при минимуме затрат (Dambroth M., Bassam N., 1983; Bassam N., 1998; Brancourt – Hulmel M., Doussinault G, Lecomte C. et al, 2003). Такие сорта крайне необходимы в Поволжье и на Урале.
Изучение морфологии и архитектоники корневых систем в онтогенезе растений с целью определения сортовых различий, а тем более проведение селекционных оценок и отборов по этим параметрам крайне затруднительно и почти невозможно в рамках существующих схем селекционного процесса. Однако состояние надземных органов их оводненность, чистая продуктивность фотосинтеза и вынос минеральных элементов косвенно указывают на активность корневой системы.
Динамика ежедневных приростов биомассы и выноса минеральных веществ с единицы площади по межфазным периодам сортами твердой пшеницы носит сложный и неоднозначный характер. Нам удалось в течение трех лет – 1995 (сильная засуха), 1997 (очень благоприятный год), 1998 (очень сильная засуха) детально изучить эти процессы у трех сортов: Харьковская 46, Безенчукская 105, Безенчукская 182. Кроме того, данные, полученные при изучении 10 сортов в 1997, 1998 гг., использованы в системном анализе формирования генотипических различий по урожайности методом главных компонент.
В 1995 году значительное преимущество сорта Безенчукская 182 по продуктивности общей биомассы и зерна формировалось за счет более интенсивного накопления сухого вещества в периоды трубкование – начало колошения - цветение и формирование зерна – созревание. Интенсивность выноса азота, рассчитанная по количеству ежедневного поступления его в надземные органы растений, в целом соответствовала динамике прироста биомассы. Аналогичного преимущества Безенчукской 182 по фосфору не обнаружено. Предположение о наличии у Безенчукской 182 фосфорэффективной системы синтетических и ростовых процессов не подтвердилось в период налива зерна.
Очень благоприятные условия 1997 года, особенно в начале вегетации стимулировали интенсивное накопление биомассы всеми сортами – в момент трубкования биомасса на 1м2 превышала на ~30,0% уровень ее накопления в 1995 году в момент созревания. При этом по выносу фосфора надземными органами сорта почти не отличались, вынос азота у сорта Безенчукская 182 был существенно выше, чем у двух других сортов. Вынос азота у сортов Харьковская 46 и Безенчукская 182 к фазе трубкования составил 9890,5 и 10988,5 мг/м2 соответственно, что равно 99,0 и 109,9 кг/га, т.е. близко к фонду минерального азота, накапливаемого в почве на черноземах чистым паром. Если учесть, что потенциальная скорость потребления азота растениями обычно выше скорости, с которой он становится доступным в результате деятельности микроорганизмов (Томпсон Л.М., Троу Ф.Р., 1982), то очевидно, что к моменту трубкования запасы азота в почве в посевах этих сортов в условиях 1997 года, были исчерпаны. В результате поступление азота у них в надземную часть прекратилось. Анализируя полученные результаты необходимо учитывать, что именно к фазе колошения нитратредуктазная активность корней и надземной части выравнивается, после колошения до восковой спелости корни превалируют в этом процессе. Кроме того, корневая система твердой пшеницы в сравнении с мягкой имеет более длительный ассимиляционный период, и она вносит более напряженный характер конкуренции за ассимиляты между корнями и зерновками в конце вегетации. К моменту восковой спелости в корнях твердой пшеницы остается 14,0 % азота, мягкой - 5,0 % (Шер К.Н., Кумаков В.А., Чернов В.К.., 1998). По данным Б.В.Березина (1984) доля корневой системы в общих дыхательных затратах растений твердой пшеницы по сравнению с мягкой выше на протяжении всего периода вегетации и особенно в репродуктивный период. Эти факты свидетельствуют о высокой функциональной активности корневой системы твердой пшеницы, которая даже в ответственные периоды формирования зерновой продуктивности может доминировать в донорно-акцепторных отношениях с надземной частью и отвлекать для активизации собственного метаболизма ее ресурсы. Значительное отставание сорта Безенчукская 105 по выносу азота, наблюдавшееся в 1997году в период от кущения до трубкования, видимо, не до конца исчерпало его запасы в почве, и он продолжал интенсивно поступать в надземную часть после трубкования. Умеренное потребление азота сортом Безенчукская 105 в период всходы - трубкование связано с более низким уровнем водопотребления, так как эти процессы сопряжены физиологически и совпадают во времени (Климашевский Э.Д., 1991).
Вынос фосфора надземными органами растений к моменту трубкования колебался в зависимости от сорта в пределах 38,1 -39,4 кг/га, что почти в два раза превышает вынос фосфора за всю вегетацию 1995 года. Поскольку поглощение фосфора осуществляется в основном корневыми волосками при непосредственном контакте с поглощающей поверхностью и зависит от густоты и разветвленности корней, можно предположить, что различий между сортами по этим параметрам корневой системы в рассматриваемый период не было. Очевидно также, что к моменту трубкования у сортов сформировалась мощная вторичная корневая система, но, несмотря на это, чтобы обеспечить потребности растущего растения корневая система должна была, продолжать интенсивно расти. Это связано с известной прямой зависимостью между процессами роста клеток корня, новообразованием цитоплазмы и поглощением веществ (Сабинин Д.А., 1957).
Продолжение интенсивных ростовых процессов и формирования урожая зерна в этой ситуации было обеспечено временным снижением суточных приростов сухого вещества надземной части и транслокацией азота и фосфора из нее в корневую систему. В последующий период от начала колошения до завершения роста стебля (цветение), наблюдался интенсивный вынос азота и фосфора. В период налива зерна наибольшие приросты биомассы, вынос азота и особенно фосфора отмечены у сорта Безенчукская 182.
В условиях сильной засухи 1998 года наиболее продуктивный и засухоустойчивый сорт Безенчукская 182 по ежедневным приростам биомассы в течение всего периода от всходов до цветения занимал промежуточное положение между Харьковской 46 и Безенчукской 105. Последние сорта проявили сильную изменчивость по исследуемому признаку, чередуя периоды интенсивного и депрессивного роста. В целом за всю вегетацию Безенчукская 182 по накоплению биомассы значимо превысила оба сорта. Характер межсортовых различий в интенсивности потребления и ежедневного выноса азота по межфазным периодам довольно точно соответствовал динамике прироста биомассы. По выносу фосфора в целом за всю вегетацию сорта Харьковская 46 и Безенчукская 105 уступили Безенчукской 182 на 23,4 и 30,9 % соответственно. Следует обратить внимание на то, что преимущество в потоке фосфора в надземную часть сорта Безенчукская 182 определилось в период от начала колошения и до созревания. Именно в этот период дефицит влаги достиг максимальных значений, следовательно, выживаемость и продуктивность растений зависели от скорости роста корней в глубину и освоения ими новых еще неиссушенных почвенных горизонтов. Известно, что фосфор сдерживает рост корней в глубину и усиливает их ветвление – т.е. меняет морфологию корневой системы для эффективного освоения ресурсов в верхних почвенных горизонтах.
Азот в наибольшей степени из всех макроэлементов влияет на рост корней, ускоряя их проникновение в глубину (Черны В., Ферик М., 1984). В связи с этим становятся понятными различия в динамике выноса минеральных элементов и их транслокации в корневую систему
В условиях сильной засухи различия между сортами по продуктивности агроценоза находятся в прямой зависимости от потребления и использования воды. Учитывая это можно предположить, что наблюдаемый параллелизм в интенсивности ежедневных приростов биомассы и выноса азота в свою очередь свидетельствует о сходстве динамики водопотребления и выносов азота, для набора исследуемых сортов твердой пшеницы. В условиях засухи более урожайны те посевы, которые формируют глубоко проникающую первичную корневую систему, состоящую из зародышевых и колеоптильных корней. По данным П.Н.Богданова (1946) слабое развитие корней – основная причина неполного потребления и использования посевом даже тех скудных запасов влаги, которыми располагает посев в засушливый год.
Таким образом, более высокая засухоустойчивость Безенчукской 182, видимо, связана с энергичной работой первичной корневой системы. Отсутствие преимущества этого сорта над менее продуктивными сортами по надземной биомассе и выносу азота в начальный период вегетации является косвенным свидетельством ускоренного роста корневой системы в это время. Основное преимущество более интенсивных сортов по выносу минеральных элементов наблюдается в предрепродуктивную и репродуктивную фазу, экстенсивные сорта выделяются на ранних этапах. К моменту трубкования сорта, стабильно формировавшие высокоозерненный колос, отдают «предпочтение» более активному потреблению азота, чем фосфора. В условиях засухи у них отношение валового азота к валовому фосфору в надземной биомассе составляло 5,0 –5,5, а у сортов с меньшим числом зерен (к ним относятся в основном низкопродуктивные формы) 4,5-4,7. К группе сортов с высоким отношением N/P отнесены: Безенчукская 182, Безенчукская степная, Гордеиформе 1434, Валентина. В период от трубкования до цветения вектор «предпочтения» у продуктивных сортов (с высокоозерненным колосом), резко менялся в сторону усиленного потребления фосфора, что вполне объяснимо с точки зрения предстоящих энергозатрат в период формирования, роста и налива зерновок (Кумаков В.А., и др., 1983).
Все современные сорта лучше, чем более ранний сорт Безенчукская 139 используют фосфорно-калийные и минимальные дозы азотных удобрений. Тот факт, что максимальные прибавки получены на фоне PK(30), где априори продукционный процесс в части связанной с минеральным питанием должен лимитироваться азотом, говорит о более эффективном усвоении современными сортами его почвенных ресурсов и лучшем использовании в ростовых процессах. Эти данные подтверждают наш вывод о том, что современные сорта твердой пшеницы отличаются высокой засухоустойчивостью и одновременно потенциальной продуктивностью, отзывчивостью на благоприятный комплекс среды. Тем не менее, повышенные дозы азота (N90) они используют хуже, чем Безенчукская 139. Преимущество продуктивных сортов на этих фонах снижается до уровня контрольного варианта без удобрений, или даже несколько ниже его. Это говорит о том, что в разряд лимитирующих факторов у продуктивных сортов перешли ростовые процессы (Macy P., 1936; цит. по Томпсон Л.М., Троу Ф.Р., 1982). Расчет эффективности использования азота и фосфора сортами в зависимости от условий года дал следующие результаты. По эффективности использования в ростовых процессах поглощенного азота в целом за вегетацию значительная изменчивость наблюдалась в засушливых условиях - продуктивные сорта характеризовались более высокими показателями. Коэффициент корреляции между массой сухого вещества синтезированного на 1 мг поглощенного азота и надземной биомассой с 1м2 в 1998 году значим на 5,0 % уровне (r = 0,55). В благоприятном 1997 году такой зависимости не наблюдалось r = 0,07. Аналогичной дифференциации сортов по использованию фосфора не обнаружено (r=0,02 в засуху и r= -0,075 в благоприятных условиях). При этом концентрация минеральных элементов в листьях и стебле у продуктивных, интенсивных сортов была ниже, чем у сортов экстенсивных и относящихся к ранним периодам селекции. Однако в мякине концентрация минеральных элементов была выше у высокопродуктивных сортов. Возможно, это связано с длительностью фотосинтетической активности колосковых, цветковых чешуй, остей, колоскового стержня и в связи с этим поздним включением механизмов реутилизации минеральных элементов из них в зерно. Полученные данные вполне объяснимы. Интенсивность фотосинтеза колоса твердой пшеницы в два раза выше, чем мягкой (Кумаков В.А. и др., 1990). Потребность в ассимилятах у крупноколосых сортов больше, поэтому период фотосинтеза колоса у них более длителен, чем у сортов с мелким колосом.
Таким образом, можно предположить, что в экстремальных условиях засухи более высокая эффективность использования азота в ростовых процессах детерминирована генетическими системами засухоустойчивости. Отзывчивость продуктивных сортов во влажные годы обеспечивается высоким валовым выносом и потреблением минеральных веществ, что также определяется уровнем адаптивности. Очевидно, что современные сорта твердой пшеницы не обладают специфическими генными системами отзывчивости ростовых процессов на повышенный уровень питания и удобрения.
Система взаимоотношений фотосинтеза, роста, потребления минеральных веществ при формировании генотипических различий по урожайности. Для выявления закономерностей формирования урожая на основе этих процессов применялись статистические методы многомерного анализа.
Метод главных компонент (Харман Г., 1972) предназначен для совместного анализа взаимосвязанных признаков. Он позволяет весь комплекс признаков распределить на группы путем нахождения нескоррелированных между собой главных компонент. В процессе их анализа выделяются так называемые гипотетические факторы, представляющие собой сложные системы, отличающиеся глубоким внутренним взаимодействием входящих в них признаков и существенной независимостью от других выделенных систем. Системы, в которых два или более признака связаны между собой обратной зависимостью, т.е. компенсируют изменение уровней друг друга, являются автокомпенсаторными или обладающими свойством авторегуляции своей эффективности (Осипов Ю.Ф., 2000). Группы тесно связанных между собой признаков могут быть выражены как один. При этом часть избыточных признаков отсеивается, что ведет к оптимизации селекционного процесса.
По данным экспериментов, проведенных в контрастные по гидротермическому режиму условиях на 10 сортах- Харьковская 46, Безенчукская 105, Безенчукская 182, Гордеиформе 1434, Саратовская золотистая, Гордеиформе 740, Гордеиформе 1479, Валентина, Степь 3, определена система взаимодействия признаков, характеризующих продукционный процесс в динамике онтогенеза, с урожайностью зерна (табл.7). Установлено три уровня взаимосвязи. Первый уровень включает признаки, объединенные в группу тесной, стабильной и независимой от условий среды взаимосвязью. Второй и третий уровни объединяют признаки, обнаруживающие тесную зависимость в условиях засухи и оптимального гидротермического режима соответственно. На первом уровне взаимосвязи признаков, кроме урожайности зерна, находятся: функция роста колоса в цветение, К.хоз и количество поглощенного фосфора в период трубкование – цветение. По отношению к ним автокомпенсаторным является признак «содержание белка в зерне». На втором уровне к признакам первой группы, положительно связанных с урожаем зерна, добавляются – число зерен в колосе, азотный уборочный индекс, площадь листьев главного побега в кущение и количество поступивших в надземную часть от цветения до созревания азота и фосфора. Автокомпенсаторными признаками на этом уровне были: «содержание белка в зерне», «функция роста боковых побегов в цветение», «Ф.ч.пр. в период трубкование - колошение». Третий уровень взаимосвязи включает - число растений на 1м2, вынос Р2О5 и азота в период кущение - трубкование, надземную биомассу в цветение, площадь листьев главного побега в цветение и все признаки первой группы. К группе автокомпенсаторных признаков на этом уровне относятся - содержание белка в зерне, вынос азота и Р2О5 в период всходы-кущение, надземная масса в кущение, площадь листьев главного побега в кущение.
7. Уровни взаимозависимости урожайности зерна и морфофизиологических признаков (по результатам факторного анализа)
| Уровень | Характер взаимосвязи | Группы признаков | |
| основная | автокомпенсаторная к основной | ||
| I | независимый от условий среды | 1.Урожай зерна агроценоза 2.Функция роста колоса в цветение 3.К.хоз. 4.Вынос агроценозом Р2О5 в период трубкование – цветение | 1. Содержание белка в зерне |
| II | проявляющийся в условиях засухи | Признаки первого уровня плюс: 1.Число зерен в колосе 2.Азотный уборочный индекс 3.Площадь листьев главного побега в кущение 4.Вынос агроценозом Р2О5 и азота в период цветение–созревание | 1.Содержание белка в зерне 2.Функция роста боковых побегов в цветение 3.Ф.ч.пр. в период трубкование- колошение |
| III | проявляющийся при оптимальном гидротермическом режиме | Признаки первого уровня плюс: 1.Число растений на 1м2 2.Вынос Р2О5 и азота в период кущение-трубкование 3. Надземная биомасса в цветение 4.Площадь листьев главного побега в цветение | 1.Содержание белка в зерне 2. Вынос азота и Р2О5 в период всходы-кущение 3. Надземная масса в кущение 4. Площадь листьев главного побега в кущение |
5. ГЕНЕТИЧЕСКИЕ ИСТОЧНИКИ И ДОНОРЫ ХОЗЯЙСТВЕННО-ЦЕННЫХ ПРИЗНАКОВ.
Доноры генов гомеостаза и элементов продуктивности. В Среднем Поволжье в процессе селекции твердой пшеницы сформировался устойчивый блок коадаптированных генов, который, контролируя формирование неспецифического гомеостаза, обеспечил повышение урожайности и устойчивости сортов (Вьюшков А.А., 2004; Мальчиков П.Н., Вьюшков А.А., Мясникова М.Г., Чиганцев Н.П., 2007). Задача селекции состоит в том, чтобы не только сохранить его, но и обеспечить эволюцию. Поэтому на современном этапе при создании исходного материала для селекции коммерческих сортов за основу берется базовый генотип, несущий коадаптированный блок генов (Хангильдин В.В., 1978). Базовые генотипы, как и все другие, не являются идеальными, могут нести нежелательные признаки - отличаться от оптимальных величин по вегетационному периоду, устойчивости к болезням, полеганию, качеству зерна. Для исправления их недостатков методами беккроссов, парных скрещиваний вводятся соответствующие гены, названные В.В.Хангильдиным (1978) реализаторами.
В группу базовых включают генотипы, проявившие в сортоиспытании высокую степень гомеоадаптивности, под которой понимается весь спектр взаимодействия генотип – среда (Сюков В.В., 2003). Еще одно не менее важное требование к этой группе – наличие коадаптированных блоков генов, контролирующих основой комплекс признаков, и наследуемых потомством сцеплено. Функционирование таких блоков генов оценивается исторически – по результатам селекции и сортообразующей способности или в системных скрещиваниях по комбинационной способности и генетическим параметрам. Последняя процедура наиболее предпочтительна, поскольку позволяет идентифицировать базовый генотип до его морального устаревания.
Наличие в исходном материале сортов несущих блоки коадапатированных генов является необходимым условием успешной селекции. Поиск базовых генотипов – носителей таких блоков важная задача селекции. Для ее выполнения необходимо решение двух последовательных исследовательских задач. Первая - идентификация гомеоадаптивных сортов, вторая - определение среди них генотипов, хорошо передающих потомству свои генетические особенности в различных условиях среды. Для решения первой задачи необходимо применение соответствующей методологии, апробированной в данных агроэкологических условиях. C.S.Lin, M.R.Binns в диаллельных скрещиваниях генотипов костра безостого изучили наследование параметров стабильности, отнесенных ими к следующим четырем типам: 1 - средовая варианса каждого генотипа не связанная с изменчивостью всего исследуемого набора сортов; 2 - эковаленса G. Wricke; 3 - варианса отклонений от линии регрессии по S.A.Eberhart, W.A.Russel; 4 - показатель Pi C.S.Lin, M.R.Binns (1988). К наследуемым параметрам отнесены 1 и 4 типы стабильности, не наследуемыми и, следовательно, бесполезными для селекции 2 и 3 типы. Последние два параметра также имели довольно тесную взаимосвязь.
Используя многолетние данные урожайности восьми сортов по разным предшественникам и фонам (всего 12 сред), были изучены следующие методы статистической оценки гомеоадаптивности: bi – пластичность по Eberhart, Russel (1966);Экорегрессия (варианса отклонений от линии регрессии) по Eberhart, Russel; ai – коэффициент мультипликативности (Драгавцев В.А. и др., 1984); Hom – коэффициент гомеостатичности (Хангильдин В.В. и др., 1978); St2 – коэффициент стабильности (Cоболев Н.А., 1980); W – эковаленса (Wricke G., 1965); Pi – мера превосходства сорта (Lin C.S., Binns M.R., 1988); ОАС – общая адаптивная способность; 2CАС- варианса специфической адаптивной способности; Sgi - относительная стабильность генотипа (Кильчевский А.В., Хотылева Л.В., 1997);
Методом двухфакторного дисперсионного анализа выявлены значимые эффекты среды, генотипов и их взаимодействия. Большинство изученных параметров дают информацию близкую по результатам ее толкования в координатах понятия «гомеоадаптивность». Только два параметра (2di, Wi) имеют значительные отклонения в характеристике сортов, подтверждая результаты C.S.Lin, M.R.Binns (1988).
Большинство параметров вошедших в первую корреляционную плеяду относятся к первому типу, т. е характеризуют признак на основе средовой вариансы (Hom, 2CAC, Sgi, ai, St2). Все эти параметры тесно связаны с OACi – характеризующей линейный уровень различий признака сорта со всей совокупностью исследуемого набора сортов и СЦГi – рассчитываемой на основе OACi и среднего по опыту значения Sgi. Кроме того, в эту корреляционную плеяду входит параметр Pi, относящийся также к наследуемому 4 типу стабильности.
Попытка определить в диаллельных скрещиваниях по B.I.Hayman наследование параметра экостабильности G.Wricke оказалась неудачной из-за нестабильности его проявления у генотипов и незначимости различий между вариантами. Достоверные различия между сортами и гибридами получены по коэффициенту мультипликативности (аi), характеризующему гомеостатичность признака «масса зерна с колоса».
Первые три места по гомеостатичности изучаемого признака заняли Безенчукская 205, Памяти Чеховича, Безенчукская степная, 5 и 6 место Гордеиформе 1734 и Безенчукская 202 соответственно. Безенчукская 182 имела промежуточное значение между крайними вариантами. Параметры Wr-Vr и Wr+Vr показали соответствие наследования признака аддитивно-доминантной модели. Коэффициент корреляции между выраженностью признака у сортов и суммой Wr+Vr был высоким R= 0,763 и указывал на положительное действие доминантных генов, усиливавших гомеостатичность. Все компоненты генетической дисперсии были значимы на 5 % уровне. Доминирование было полным параметр - (H1/D) близок к единице, наблюдалась значительная ассиметрия частот доминантных и рецессивных аллелей - параметр Н2/4Н1, значительно отклонялся от 0,25, параметр F был больше нуля. Доминантных генов в популяции больше, чем рецессивных. Параметры Fr значимы для всех сортов (Fr Памяти Чеховича и Безенчукской 205 самые большие по величине). Наиболее эффективными донорами доминантных генов гомеостатичности могут быть сорта Безенчукская 205, Памяти Чеховича. Таким образом, достаточно выраженный генетический контроль коэффициента мультипликативности, позволяет его и тесно коррелирующие с ним параметры отнести к группе наследуемых признаков и использовать их для эффективной оценки адаптивности и стабильности генотипов твердой пшеницы.
Из всего набора методов оценки адаптивности генотипов и дифференцирующей способности среды комплекс параметров А.В.Кильчевского, Л.В. Хотылевой (1997) следует признать наиболее информативным. Результаты нашего эксперимента организованного и выполненного по этой методике в 2005-2006 гг. в 6 средах распределенным по двум географическим пунктам (Безенчук, Волгоград) и включавшего 30 лучших генотипов селекции Самарского НИИСХ, позволяют дать им полную характеристику по параметрам гомеоадаптивности. Сортами, сочетающими высокую продуктивность со стабильностью формирования урожая, оказались: Гордеиформе 1761, Памяти Чеховича, Безенчукская 205, Леукурум 1751, Безенчукская 207, Леукурум 1753, Безенчукская Нива, Марина, Безенчукская степная, Гордеиформе 1732. Все сорта, по величине СЦГi, включенные в список продуктивных и стабильных являются кандидатами для использования их в качестве базовых генотипов. Лучшие из них (Памяти Чеховича, Безенчукская 205, Безенчукская степная ) изучены в диаллельных скрещиваниях по методике Хеймана в условиях Безенчука (2004, 2005гг.) и Волгограда (2005 г.) по признакам продуктивности растений. Эти сорта, как было показано выше, имеют наибольшую концентрацию доминантных генов гомеостатичности признака «масса зерна с колоса». Генетико-статистическая характеристика сортов по основным элементам продуктивности растений дополняет эту информацию и формирует целостное представление о наличии коадаптированных блоков генов.
Наибольший интерес представляли признаки «число зерен с растения», «К.хоз.» и «масса 1000 зерен». В условиях Волгограда по всем признакам получены высокозначимые различия между родительскими сортами. По всем признакам отмечен положительный эффект доминантных генов. По признакам «число зерен с растения» и «масса 1000 зерен» наблюдалось полное доминирование в локусах (H1/D) 1,0, по К.хоз. неполное доминирование(H1/D) < 1,0. Несмотря на преобладание доминирования, аддитивные эффекты в наследовании признаков были значимы на 5% уровне. Значимая ассиметрия частот доминантных и рецессивных аллелей (параметр H2/4H1) наблюдалась только по К.хоз.
В условиях Безенчука в 2004-2005 гг. генетические параметры, определенные на этом же наборе сортов и диаллельных гибридах F1 (2004г) и F2 (2005г.), показали, отсутствие значимых эффектов генов на признак число зернен с колоса главного побега в оба года исследований. При этом в обоих случаях по признаку получены значимые на 5 % уровне различия между сортами и гибридами. В наследовании признака «К.хоз.» превалировали доминантные гены, проявившие значимые аддитивные эффекты и действовавшие в направлении отбора. Показатель степени доминирования (H1D) значительно превышал единицу. Весь комплекс параметров свидетельствует о хорошей наследуемости признака и целесообразности отбора из гибридных популяций в поздних поколениях. Аддитивные эффекты генов превалировали в оба года в контроле массы 1000 зерен. В 2004 году рецессивные, а в 2005 году доминантные гены увеличивали признак. Прогнозируется высокий эффект отбора в ранних поколениях гибридных популяций.
Наибольшее количество доминантных генов с аддитивным эффектом, контролировавших признак «К.хоз» в 2004 году имели сорта Памяти Чеховича и Безенчукская 205, наименьшее их количество было у сортов Безенчукская 202 и Гордеиформе 1734. Безенчукская 182 по количеству доминантных генов занимала промежуточное положение между крайними вариантами. Безенчукская степная по их частоте в локусах была близка к первой группе сортов (Памяти Чеховича, Безенчукская 205). Поскольку коэффициент корреляции между выраженностью признака у родителей и уровнем доминантности ( R ) был невысоким, можно предположить, что в 2004 году часть рецессивных генов действовали положительно увеличивая признак. В 2005 году в контроле признака « К.хоз» у сортов Памяти Чеховича и Безенчукская степная принимали участие максимальное количество доминантных генов. Безенчукская 202 имела минимальное их количество, у Гордеиформе 1734 и Безенчукской 182 формирование признака проходило под действием равного числа доминантных и рецессивных аллелей. Максимальное количество положительно действовавших в 2004 году на массу 1000 зерен рецессивных генов имел сорт Безенчукская 202. Однако если учесть, что коэффициент корреляции между доминированием и величиной признака у родителей был невысокий можно предположить наличие у других сортов положительно действовавших на признак доминантных генов. В 2005 году доминирование было сильнее, но его уровень также был недостаточен для однозначной системы генов. Тем не менее, максимальное количество доминантных генов имели сорта Безенчукская степная, Памяти Чеховича, Безенчукская 205. Рецессивные аллели преобладали у сорта Безенчукская 182.
Таким образом, в условиях Волгограда на формирование всего комплекса признаков оказывает влияние хорошо скоррелированная с ним физиологическая система жаростойкости гаметогенеза и засухоустойчивости ростовых процессов, контролируемая доминантными генами с сильными аддитивными эффектами. В условиях Безенчука, несмотря на значительную схожесть рангов сортов по развитию признаков с результатами, полученными в Волгограде, все признаки контролировались разными генетическими системами, а по числу зерен в колосе эффекты среды превысили эффекты генов.
Полученные результаты объясняются высоким уровнем адаптивности к агроэкологическим условиям Безенчука сортов, включенных в исследования. В условиях более сильной дифференцирующей способности среды г.Волгограда, исследуемая сортовая популяция обнаружила не только новые свойства сортов и более высокий уровень генотипической изменчивости, но и лежащую в их основе иную генетическую вариабельность.
Сорта Памяти Чеховича, Безенчукская степная и Безенчукская 205 являются донорами доминантных генов жаростойкости и засухоустойчивости, которые проявляют свое действие в соответствующих условиях в виде положительных эффектов на компоненты продуктивности растений. Эти же сорта в условиях Безенчука превосходят сорт Безенчукскую 182 по концентрации положительно действовавших на К.хоз и массу 1000 зерен аллелей и являются носителями доминантных генов обеспечивающих гомеостатичность формирования признака «масса зерна с колоса» в разнообразных условиях среды.
Проведенные исследования доказывают, что в процессе селекции сортов Памяти Чеховича, Безенчукская 205, Безенчукская степная, произошло объединение двух генетических пулов твердой пшеницы – саратовского и безенчукского, функционировавших до этого относительно автономно. Новые сорта с «безенчукскими генами» унаследовали высокую продуктивность, а с «саратовскими» - высокую засухоустойчивость. Более того, они превосходят саратовские сорта по засухоустойчивости (по многолетним испытаниям в условиях Волгограда) и сорт Безенчукскую 182 по урожайности в относительно благоприятных условиях. В данном случае получена своеобразная трансгрессия по адаптивности (гомеостатичности) в результате комплементарного взаимодействия генов засухоустойчивости и высокой продуктивности. Механизм этого взаимодействия может быть следующим. Засухоустойчивость способствует реализации потенциала продуктивности сортов твердой пшеницы, в период вегетации которой на Юго-Востоке почти во все годы случаются периоды засухи и высокотемпературные стрессы. В то же время сама продуктивность, которая по определению П.А.Генкеля (1967) является одним из факторов засухоустойчивости, положительно воздействует на нее через активацию ростовых и синтетических процессов.
Исходный материал и генетические доноры качества зерна. Весь спектр признаков качества зерна твердой пшеницы распределяется на три группы. Первая включает признаки, которые определяются при анализе зерна - стекловидность, натура, масса 1000 зерен, содержание клейковины и белка, зольность, цвет зерна, число падения. Вторая – признаки, оценивающие качество крупки – цвет и наличие спексов (темных вкраплений). Третья – признаки, характеризующие реологические свойства теста и качество макаронных изделий (Васильчук Н.С., 2001; Голик В.С., Голик О.В., 2007).
Стекловидность. По нашим наблюдениям наиболее сильная дифференцирующая способность по стекловидности зерна характерна для обедненных азотом фонов, формируемых по предшественникам озимая рожь и овёс на зерно без внесения удобрений. В этих условиях стекловидное зерно имели сорта: Харьковская 46, Безенчукская 182, Безенчукский янтарь, Безенчукская 200, Безенчукская степная, Безенчукская 205, Саратовская золотистая, Ник, Валентина, Жемчужина Сибири, Ангел, Краснокутка 10. Сорта, имеющие продолжительные периоды всходы – колошение и налива зерна, в этих условиях в отдельные годы заметно снижают процент стекловидности в сравнении с посевом по чистому пару. Кроме того, на бедных фонах отчетливее проявляется способность сортов давать стекловидное зерно с хорошим цветом при перестое после созревания. В этих условиях преимущество уже имеют более поздние сорта. В целом по многолетним наблюдениям в условиях Безенчука высокий уровень стекловидности зерна и стабильность этого признака характерны для сортов среднеспелого биотипа. Большинство современных сортов твердой пшеницы способны формировать зерно с высокой стекловидностью в широком диапазоне условий среды. Возможным механизмом сохранения признаков качества зерна на высоком уровне (содержание белка, клейковины, стекловидность) в процессе селекции на урожай, кроме увеличения уровня адаптивности и выноса азота, является более интенсивная реутилизация азота из стебля. Во всяком случае, в местностях (на фонах) с низкой стекловидностью зерна и при отсутствии азотной подкормки отбор по признаку эффективен (Hdjichristodoulou A., 1979; цит. по Голик В.С., 1986). По мнению В.С.Голик и О.С Голик (2008) стекловидность – один из наиболее реализуемых признаков. Он, являясь наследственным признаком (Porceddu E., 1998), достаточно легко передается потомству.
Натурная масса зерна. Зерно с высоким натурным весом обладает хорошими мукомольными качествами, имеет высокий выход семолины хорошего качества (Васильчук Н.С., 2001). Кроме того, себестоимость перевозки единицы веса зерна с низкой натурой повышается, транспортные расходы снижают конкурентоспособность конечной продукции, производимой из такого зерна. В России согласно ГОСТа 9353-90 для первого класса показатель натуры зерна должен быть не ниже 770, для второго - 750 и третьего, четвертого и неклассной – 710г/л.
Натура зерна зависит как от условий выращивания, так и от генотипа сорта. Используя методику А.В.Кильчевского и Л.В.Хотылевой (1997) нами проведена оценка 13 сортов конкурсного сортоиспытания в восьми средах. Изучение проведено в течение четырех лет (2004-2007гг.) по предшественникам: чистый пар и овес на зерно. Результаты оценки параметров адаптивной способности и стабильности признака «натура зерна» показали значительные различия сортов. Наибольшими эффектами ОАСi обладали сорта Безенчукская Нива, Марина, Безенчукская степная и Безенчукский янтарь. Самые низкие показатели ОАСi имели сорта - низкорослый Гордеиформе 1732 (ген Rht1) и среднерослый Памяти Чеховича (ген RhtAnh). Выявлена слабая отрицательная связь между величиной признака ui + vi и стабильностью 2САСi (r = -0,42), что означает снижение стабильности при увеличении признака в процессе селекции. Однако абсолютные показатели натуры зерна лучших по средним показателям сортов в большинстве сред также были одними из самых высоких. Относительная стабильность генотипа Sgi колебалась от 2,8% у Памяти Чеховича до 5,9 % у Гордеиформе 1732. Цель селекции повышение величины признака и стабилизация его формирования в различных средах достигается отбором генотипов с высокой ОАСi, средними показателями стабильности (2САСi = 554,1-794,1; Sgi.= 3,2-3,8) и близким к единице параметром Эберхарда – Рассела (bi = 1,01-1,13). Этими величинами оценивается признак у сортов - Марина, Безенчукская Нива, Безенчукская степная и Безенчукский янтарь.
Благоприятный прогноз для отбора генотипов с высокой натурой зерна получен нами в 2005 году при анализе генетических параметров Хеймана на основе диаллельных скрещиваний адаптированных к местным условиям сортов, но значимо различающихся по изучаемому признаку. Анализировались шесть сортов и их прямые диаллельные гибриды F2. Доминантные гены увеличивали признак (R= -0,711). Доминирование в локусах было неполным, аддитивный компонент был значим ((H1/D = 0,731). Наблюдалась незначительная ассиметрия в частотах доминантных и рецессивных аллелей (H2/ 4H1 = 0,22) c преобладанием доминантных генов ((4DH1+ F) / (4DH1- F) = 1,00). Число групп генов, по которым различались родители, равнялось двум. Наибольшее количество доминантных генов имели сорта: Безенчукская 205, Гордеиформе 1734, Безенчукская 202, Безенчукская 182. Сорт Безенчукская степная наряду с доминантными генами имел рецессивные гены. Сорт Памяти Чеховича имел только рецессивные гены, которые обусловили формирование у него зерна с относительно низкой натурной массой. Все генетические параметры достоверны, имеет место идеальная ситуация для проведения селекционной работы по совершенствованию признака.
Масса 1000 зерен. Этот признак является не только значимым компонентом продуктивности растений, но и важным критерием качества зерна и его технологичности. Крупное зерно повышает выход крупки (семолины) при помоле. Это связано с отношением эндосперм / отруби, которое у крупного зерна шире (Quick J.S., Williams N., 1972). В данном случае отбор селекционного материала по массе 1000 зерен опосредованно увеличивает выход крупки. По данным Н.С.Васильчука (2001) признак легко поддается селекционному улучшению. Результаты наших исследований наследования массы 1000 зерен в диаллельных скрещиваниях, приведенные выше, подтверждают этот вывод. Во всех средах (Волгоград 2005 г., Безенчук 2004, 2005 гг.) аддитивный компонент был значим, что позволяет прогнозировать хороший эффект отбора, начиная с ранних поколений.
Среди, изученных нами инорайонных сортов наибольшая частота встречаемости крупнозерных форм наблюдалась в саратовском и краснокутском материале. Сорта Саратовская золотистая и Валентина (НИИСХ Юго-Востока), вовлеченные нами в селекционный процесс, показали высокую способность давать потомство с крупным зерном. Сорт Валентина показал высокую комбинационную способность по признаку в двутестерных скрещиваниях (Мясникова М.Г., 2006). С привлечением генетической плазмы сортов Валентина и Саратовская золотистая получены крупнозерные сорта Безенчукская 200, Безенчукская степная, Памяти Чеховича, Безенчукская 205, Марина, Безенчукская 207, Безенчукская Нива. Потенциал этих сортов по массе 1000 зерен равен 48-50 грамм.
Дальнейшее наращивание потенциала признака признано нецелесообразным (Мальчиков П.Н., Вьюшков А.А.,.Мясникова М.Г., 2005). В связи с этим основные усилия в селекционной работе предпочтительнее сосредоточить на повышении стабильности признака в различных условиях среды. По нашим данным лучшими сортами в этом плане являются: Марина, Валентина, Безенчукская 182, Безенчукская степная, Краснокутка 10, Краснокутка 12, Безенчукская Нива, Безенчукская 207.
Содержание белка в зерне. Для сортов с высоким содержанием белка в зерне характерно повышенное количество ресурсов азота растения, приходящееся на единицу массы зерна растения, являющегося показателем обеспеченности зерна азотом (ПозN), который отрицательно коррелирует с признаком «К.хоз». Поэтому современные продуктивные сорта с высоким К.хоз имеют пониженное содержание белка в зерне.
В представлениях А.Н.Хохлова (1991) весь процесс жизнедеятельности растений определяется динамичным балансом двух функций: созданием урожая репродуктивной частью и «поддержкой» активного состояния вегетативных органов и, прежде всего корневой системы. Соответственно этому он макроструктуру суммарного фотосинтетического потенциала (МФП) выражает как сумму репродуктивно ориентированного фотосинтетического потенциала (ФПР) и фотосинтетического потенциала поддержки (ФПП), обслуживающего потребителей нисходящего тока – вегетативную сферу и корни. Чрезмерное увеличение мощности ФПР в ущерб мощности ФПП может привести к снижению снабжения азотом настолько, что нарушает функционирование ФПР и подрывает основы формирования урожая. В случае же доминирования ФПП колос формируется слабый, сила его «запроса» не в состоянии доминировать в процессах распределения азота и потенциал корневой системы остается нереализованным. Показана сильная зависимость между пропорциями 2-3 верхних ярусов стебля и МФП. Оптимальным признано отношение линейных размеров подколосоносного междоузлия к колосоносному 1,0-1,1.
В наших экспериментах содержание белка в зерне у современных сортов твердой пшеницы, выращенных по предшественнику овес на зерно, незначимо отличалось от величины, показанной высокобелковистым сортом Харьковская 46. В опыте по чистому пару, только два сорта – Марина и Безенчукская 182 статистически достоверно уступили сорту Харьковская 46 по этому признаку. Чистый пар, обладая большими запасами азота, уменьшает как средовое, так и генотипическое варьирование признака, «сглаживая» различия между сортами. Кроме того, в посевах по чистому пару обнаружено значимое преимущество всех современных сортов над Харьковской 46 по признаку «К.хоз». По предшественнику овес на зерно достоверное превосходство над Харьковской 46 по К.хоз. имели сорта: Памяти Чеховича, Безенчукская степная, Марина.
Наибольший интерес для селекции из всех изученных сортов, представляет сорт Памяти Чеховича. Он сочетает в своем генотипе высокий К.хоз (относительное преимущество над Харьковской 46 составляет 30 %) и высокое содержание белка - различия с сортом Харьковская 46 незначимы в экспериментах по двум предшественникам.
Высокий выход зерна у этого сорта обусловлен функционированием гена редукции высоты растений – Rht Anh, унаследованным им от мексиканского сорта Аnhinga. Довольно значительное снижение высоты растений, уменьшило у этого сорта количество азота, накапливаемого до цветения в вегетативной массе и реутилизируемого в зерно в период налива. Следовательно, можно предположить, что значительная часть белка в зерне у сорта Памяти Чеховича синтезируется благодаря активному функционированию корневой системы в постфлоральный период и поступлению азота из почвы в зерно.
Изучение соотношения длины верхнего междоузлия и суммы нижележащих междоузлий стебля, характеризующее (по А.Н.Хохлову, 1991) структуру МФП, показало, что у среднерослых сортов Безенчукская степная и Памяти Чеховича оно было стабильным при выращивании по двум предшественникам. При этом по предшественнику овес на зерно у среднерослых сортов оно было достоверно меньше, чем у Безенчукской 139 и Безенчукской 182, что указывает на значительную роль генотипа в формировании структуры их МФП.
Итак, ряд современных сортов твердой пшеницы, обладающих высоким потенциалом продуктивности, который в значительной степени обеспечен высокими значениями К.хоз., способны компенсировать негативные эффекты этого признака на содержание белка в зерне. Этот результат демонстрирует реальность эволюционных каналов, обеспечивающих рост урожайности твердой пшеницы путем увеличения К.хоз. и одновременно стабилизирующих признак «содержание белка в зерне» на оптимальном уровне. В связи с этим большое значение имеет характеристика исходного материала по адаптивности и стабильности признака «содержание белка в зерне». Кроме того, для успешной селекции важен выбор фона для отбора с высокой дифференцирующей способностью среды.
В девяти опытах (пять по пару, четыре по овсу на зерно) изучено 12 сортов. По эффектам общей адаптивной способности признака выделились сорта: Харьковская 46, Безенчукская 200, Безенчукская 139, Безенчукская 207. Значения этого параметра у сортов Гордеиформе 1732, Безенчукская Нива и Марина варьировали ниже средней величины для исследуемой популяции сортов. Близкими к средней величине эти значения были у остальных сортов – Безенчукский янтарь, Безенчукская 182, Безенчукская степная, Памяти Чеховича. Самыми нестабильными (параметр 2 САСi ) были сорта: Безенчукская Нива, Безенчукская 207, Марина, Харьковская 46, Безенчукская 200. Наиболее стабильными были сорта Безенчукская 182 и Гордеиформе 1732. По параметру относительной стабильности (Sgi ) самым стабильным сортом следует считать Безенчукскую182. Не выявлено связи между уровнем развития признака Ui +Vi и его стабильностью 2 САСgi (r=0,17). Коэффициент корреляции между 2 (G*E)gi и САСgi был средним, что свидетельствует о наличии эффектов дестабилизации. Превалирование эффектов дестабилизации подтверждают коэффициенты Kgi, которые у большинства сортов были больше единицы. Коэффициент нелинейности Lgi колебался от 0,3 до 1,6, что указывает на неоднозначный ответ сортов по характеру линейности на изменение среды.
Судя по величине bi (параметр Эберхарда – Рассела) наибольшей отзывчивостью на среду обладают сорта: Марина, Безенчукская 200, Безенчукская степная, Безенчукская Нива, Памяти Чеховича. В этом списке нестабильные в изучавшихся условиях среды сорта Марина и Безенчукская Нива. Это свидетельствует о том, что улучшение среды в значительной степени может компенсировать недостаточное накопление белка в зерне этих сортов в неблагоприятных средах.
Очевидно, что задачей селекции на содержание белка в зерне в Среднем Поволжье является одновременный отбор образцов на ОАС и стабильность. Оценка и отбор генотипов по селекционной ценности генотипов (СЦГi), обеспечивает ее выполнение. Лучшим сортом по этому параметру является Безенчукская 182. Высокое значение параметров СЦГi и уровня признака (Ui+Vi) сорта Харьковская 46 нивелируются низкой его урожайностью.
В целом необходимо подчеркнуть, что тенденция снижения накопления белка в зерне с ростом продуктивности современного сортимента твердой пшеницы Самарского НИИСХ, имеет место. Однако в условиях хорошего агрофона все сорта накапливают оптимальное количество белка в зерне – около 16 %.
Максимальное развитие признака (> 16,0%) обеспечили паровые предшественники в 2005 - 2007 годах, минимальное (< 14,0%) предшественник овес на зерно в эти же годы. Наибольшая дифференцирующая способность проявилась в средах – «пар 2003 год» и «овес на зерно 2005 год». Единственная среда, которая соответствуют по классификации Е.Н.Синской (1963) анализирующему фону – «пар 2003 год». Таким образом, применяемые фоны в селекции на содержание белка в зерне в целом соответствуют задачам и целям в этой области.
Качество макаронных изделий. Анализ готовых макаронных изделий является заключительным этапом качественной оценки зерна твердой пшеницы. Государственная комиссия Российской Федерации по сортоиспытанию сельскохозяйственных культур определила количественные критерии, характеризующие готовые изделия по их прочности, цвету (блеску), коэффициенту разваримости и потери сухих веществ при варке макаронных изделий. Варочные свойства макаронных изделий зависят от содержания белка, клейковины и ее качества (Matsuo R.R., Dexter J.I., 1980; Долгалев М.П., Тихонов В.Е., 2005). Качество клейковины, определяемое ее реологическими свойствами, имеет более тесную связь с варочными свойствами макарон (Feillet P., 1988). В свою очередь реологические свойства и сила клейковины тесно взаимосвязаны с компонентами спектра глиадинов – 45 и 42. Сорта твердой пшеницы по аллеломорфному составу локусов (Cli-B1 / CluB3) обычно распределяются на два типа. Первый тип объединяет – глиадин 42 (-42), ассоциированный с низкомолекулярным глютенином первого типа (LMW – 1). Второй тип – глиадин 45 (-45) ассоциирован с низкомолекулярным глютенином второго типа (LMW -2). Компонент глиадиновой фракции белка 45 функционально не связан с сильной клейковиной, его маркирующие свойства объясняются тесным сцеплением с субъединицей 2 низкомолекулярных глютенинов (Pogna N.E. et al, 1990; Porceddu E. et al, 1998).
В большинстве лабораторий, ведущих селекцию сортов твердой пшеницы с учетом качества клейковины, используется комплекс методов оценки реологических свойств теста. Обычно он включает тесты по индексу деформации клейковины (ИДК), SDS-седиментации, оценку на фаринографе и миксографе. Между этими параметрами установлена тесная корреляционная связь (Васильчук Н.С., 2001). Наиболее производительный и простой тест – SDS-седиментация. В Самарском НИИСХ принята следующая схема селекции сортов с высоким качеством клейковины и кулинарных свойств макарон. Изучение рабочей коллекции, сортов межстанционного, конкурсного сортоиспытания и базовых генотипов проводится с определением параметров: прочности сухих макарон, цвета макарон, разваримости макарон, потери сухих веществ при варке, количества клейковины, ИДК, SDS–седиментации, микросографической оценки, числа падения и содержания каротиноидных пигментов. В предварительном сортоиспытании и контрольном питомнике определяются SDS–седиментация и содержание каротиноидных пигментов. Отбор элитных колосьев и линий в селекционных питомниках проводится с учетом качества клейковины родительских сортов и маркирующего его признака - окраски колосковых чешуй (Кудрявцев А.М. и др., 1993). Учитывая хорошую наследуемость признаков качества клейковины и макарон, и то обстоятельство, что в ранних поколениях селекционный материал наиболее интенсивно бракуется по степени адаптивности, принятая схема наиболее рациональна с позиций экономики и трудовых затрат.
Комплексная многолетняя оценка коллекционного материала позволила выделить источники высоких реологических свойств теста - Харьковская 9, Харьковская 13, Харьковская 15 (УкрНИИРСиГ), USMP 13, Vic (США), Гордеиформе 740, Гордеиформе 942, Гордеиформе 1732 (Самарский НИИСХ), Саратовская золотистая, Валентина, Ник (НИИСХ ЮВ) и др.
В процессе селекции произошло существенное улучшение прочности макарон. Большинство современных сортов по данным многолетнего эксперимента достоверно превышают по этому показателю Харьковскую 46 и Безенчукскую 139. По цвету макарон выделяется только сорт Безенчукская 207, который достоверно превышает старые и некоторые современные сорта. Если учесть, что по содержанию каротиноидов в зерне современные сорта значительно превосходят Харьковскую 46 и Безенчукскую 139, то можно предположить, что цвет макарон зависит не только от количества каротиноидов в зерне, но в значительной степени от других генетических систем. По коэффициенту разваримости и потере сухих веществ достоверных различий между изученными сортами не получено, но оптимальной величине по ГОСТу (3,0…4,5) соответствовали значения признака у сортов Безенчукская 207 и Памяти Чеховича.
В межстанционном сортоиспытании, где кроме сортов Самарского НИИСХ в течение 3-х лет изучались высококачественные сорта НИИСХ Юго-Востока и сорт Оренбургская 10, получены схожие с вышеприведенными результаты. Подтвердилось превосходство современных сортов по прочности макарон и их цвету. По коэффициенту разваримости достоверных межсортовых различий не получено. Оптимальным параметрам разваримости соответствовали данные сортов Безенчукская 182, Памяти Чеховича, Оренбургская 10, Саратовская золотистая и Безенчукская степная. По цвету макарон выделились сорта Безенчукская степная, Саратовская золотистая и Ник. В этом эксперименте получены значимые различия по величине потерь сухого вещества при варке. Минимальные потери сухого вещества наблюдались у сортов Памяти Чеховича (4,6%), Саратовская золотистая (4,9%), Валентина (5,1%) и Ник (5,4%). Все сорта по этому показателю соответствовали требованиям мирового рынка, согласно которым потеря сухих веществ при варке не должна превышать 7,0%.
Сорта Безенчукская степная, Памяти Чеховича и Марина во всех ситуациях обладали способностью образовывать клейковину, качество которой по тесту SDS – седиментации, превышая уровень 40,0 мл, соответствовало современным требованиям мирового рынка.
Содержание каротиноидов в зерне не зависело от предшественника. Большинство современных сортов значимо превосходят по величине этого признака Харьковскую 46 и Безенчукскую 139. Признак незначительно варьировал по годам, обнаруживая симмиллярную реакцию сортов на условия среды. Ранги сортов при этом практически не меняются. Содержание каротиноидов в зерне во многом определяет цвет макаронных изделий. В то же время рядом исследователей убедительно показано, что цвет макаронных изделий зависит от двух генетических систем, одна из которых контролирует синтез желтого пигмента, другая активность окислительных ферментов (Созинов А.А., 1985; Вьюшков А.А., 1998). Таким образом, высокое содержание каротиноидов в зерне является только предпосылкой, или лишь одним необходимым условием получения из такого зерна макаронных изделий лимонно-желтого цвета. Несмотря на то, что в конкурсном сортоиспытании Самарского НИИСХ с 1998 года испытываются сорта с высоким содержанием каротиноидов в зерне, превышающие по этому показателю стандартные сорта (Харьковская 46, Безенчукская 139, Безенчукская 182) на 20-50 %, только в 50,0% опытов наблюдалась значимая связь между признаками (табл.8). Четкого влияния предшественника на эту связь не наблюдалось. Наиболее вероятно влияние условий налива и созревания зерна (температурный режим, осадки. В пользу этого предположения указывают данные 2005 года, когда связь между признаками была достоверной на 5 % уровне значимости во всех блоках сортоиспытания по обоим предшественникам. Аналогичный вывод сделан М.Г.Евдокимовым (2006) для условий Западной Сибири.
8. Коэффициенты корреляции между содержанием каротиноидов и цветом макарон. конкурсное (КСИ), межстанционное (МСИ) сортоиспытание.
| Питом-ник | Годы и предшественники | ||||||||||
| 1998 | 2000 | 2003 | 2004 | 2005 | 2006 | 2007 | |||||
| пар | пар | пар | пар | зябь | пар | зябь | пар | зябь | пар | зябь | |
| КСИ Блок 1 | 0,31 | - | 0,41* | 0,18 | 0,32 | 0,44* | 0,73* | 0,76* | 0,15 | 0,02 | 0,47* |
| КСИ Блок 2 | - | - | - | - | - | 0,84* | 0,72* | 0,02 | 0,47* | - | - |
| МСИ | - | 0,07 | - | - | - | 0,55* | - | - | - | - | - |
Сильное влияние на цвет макаронных изделий может оказывать число спексов (темных вкраплений) в крупке. Основная причина их присутствия в крупке - наличие больного «черным зародышем» зерна, возбудителями которого являются грибковые патогены Bipolaris sorokiniana Sacc, Alternaria tenius Fr. и Alternaria triticina Pras) (Васильчук Н.С., 2001). Учитывая негативное влияние окислительных ферментов и спексов на цвет макаронных изделий, селекционные программы многих лабораторий предусматривают непрерывное увеличение концентрации каротиноидных пигментов (Васильчук Н.С., 2001; Голик В.С., Голик О.В, 2008). Наличие доноров существенно может повысить эффективность этого направления селекции. Начиная с 90-х годов ХХ века, широко используется как донор высокого содержания каротиноидных пигментов сорт Саратовская золотистая. Содержание каротиноидов в зерне этого сорта в большинстве испытаний в условиях Безенчука колебалось в пределах 600-700 мкг %, что в 1,5-2,0 раза выше, чем у большинства других сортов применявшихся в качестве исходного материала. Такой уровень межсортовых различий позволил нам изучить донорские свойства сорта Саратовская золотистая с применением генетико-биометрических методов (шестипараметорная модель и диаллельный анализ по Хейману) и гибридологического анализа с определением числа генов. В последнем случае в качестве тестеров применялись сорта Харьковская 9 и Гордеиформе 740, содержание каротиноидов в зерне у которых было в 2,1 и 1,8 раза меньше, чем у Саратовской золотистой.
Признак наследовался в результате аллельного взаимодействия генов по аддитивно-доминантному типу. Гибридологический анализ позволил выявить наличие у Саратовской золотистой двух сильных генов с аддитивным эффектом, что характеризует отличные донорские качества этого сорта по содержанию каротиноидов в зерне.
В селекционных программах Самарского НИИСХ в качестве источников высокого содержания каротиноидов использовались сорта: Саратовская золотистая, Vic (CША), ЛН-209-36, Светлана, Новодонская. Кроме того, базовые сорта Безенчукская 182, Безенчукский янтарь, Оренбургская 10, Гордеиформн 814, которые широко привлекались для гибридизации, имеют повышенное на 15-20 % содержание каротиноидов в зерне в сравнении с сортами Безенчукская 139 и Харьковская 46.
Наиболее результативным было применение сорта Саратовская золотистая. Именно с этим сортом связано резкое увеличение числа линий в конкурсном сортоиспытании с содержанием каротиноидов, на 25-50% превышающих стандарты (Безенчукская 139, Безенчукская 182). В последние годы число таких линий доведено до 40…60 %.
В процессе селекции получены линии с концентрацией каротиноидов в зерне сравнимой с сортом Саратовская золотистая. В том числе такие линии получены от скрещивания родительских форм, происходящих от сорта Саратовская золотистая, но имеющих по сравнению с ним значимо меньшее содержание каротиноидов в зерне. В частности популяция Безенчукская 200/ Гордеиформе 1434 дает значительный процент такого селекционного материала. Линия Леукурум 1755 этого происхождения стабильно превосходит стандарт Безенчукскую 182 на 40-50% и не уступает Саратовской золотистой. Аналогичные результаты получены по линии Леукурум 1756, происходящей от скрещивания Леукурум 1675/ Гордеиформе 1434. Сорт Леукурум 1675 унаследовал повышенное содержание каротиноидов от ЛН–209-36. Уровень их концентрации в этом сорте сравним с сортом Гордеиформе 1434.
Таким образом, обе линии представляют результат трансгрессии по содержанию каротиноидов. В первой линии (Леукурум 1755), видимо, произошло объединение (через Безенчукскую 200 и Гор.1434) генов Саратовской золотистой. Во втором случае трансгрессия получена в результате рекомбинации генов от Саратовской золотистой и ЛН–209-36. Учитывая, что Саратовская золотистая имеет два сильных гена, контролирующих синтез каротиноидов в зерне, можно предположить, что Безенчукская 200 и Гордеиформе 1434 являются носителями неаллельных генов. Предварительно им можно дать символы Yellow 1 и Yellow 2. Ген (или блок генов) от ЛН-209-36 неаллелен Yellow 2.
Возможности дальнейшего прогресса признака в процессе селекции на основе трансгрессий вполне реальны. Например, линия 653д-44, изучающаяся в конкурсном сортоиспытании Самарского НИИСХ на 25…28 % превышает по содержанию каротиноидов Саратовскую золотистую. Эта трансгрессия получена от скрещивания Памяти Чеховича с линией 356д-14, которая происходит от межвидовой гибридизации Безенчукская 182/ Лютесценс 1293 // Гордеиформе 740 /3/ Саратовская золотистая.
Исходный материал для селекции на устойчивость к хлебному пилильщику. Из всего многообразия признаков, обеспечивающих ту или иную степень устойчивости к вредителю, выполненность соломины паренхимной тканью - единственный признак способный надежно обеспечить защиту растений от его повреждений (Щеголев В.Н., 1931). Эффективность его использования в селекции зависит от наличия адаптированных доноров соответствующих генов. В Самарском НИИСХ на почти изогенных линиях, созданных на основе рекуррентного сорта Безенчукская 139 и трех доноров генов выполненности соломины (Апуликум 401, Um 6001, к-50989) в многолетнем эксперименте изучены эффекты этих генов на урожайность и ее компоненты, длину соломины, устойчивость к стрессовым факторам, качество зерна. Негативного воздействия выполненности соломины на все изученные признаки не наблюдалось. Установлено влияние степени адаптивности донора генов выполненности на продуктивность изогенных линий – самые продуктивные линии аналоги Безенчукской 139, получены на основе наиболее адаптированного донора Апуликум 401. Наследование признака в диаллельных скрещиваниях в большинстве ситуаций соответствовало аддитивно-доминантной модели. Гибридологический анализ признака позволил установить, что взаимодействие в расщепляющихся поколениях идет между следующими тремя генетическими системами: 1) генетическая система, имеющая один основной (сильный) ген, который можно обозначить SSt (Solidness stem) или систему дупликатных, дупликатно-комплементарных или комплементарных генов – 2 или реже 3 гена. Наличие этой генетической системы является условием формирования максимальной выполненности соломины паренхимной тканью и стабильности проявления этого признака в фенотипе; 2) генетическая система, состоящая из генов модификаторов – усиливающих действие первой генетической системы и в основном проявляющихся как гены антиингибиторы (супрессоры) – 1 или реже 2 гена. 3) генетическая система рецессивных генов-ингибиторов сердцевины стебля, которые имеются у всех изученных нами полостебельных генотипов в количестве 1 или реже 2-х генов. Находясь в гомозиготном состоянии рецессивные гены этой системы, подавляют действие первой генетической системы. В свою очередь гены этой системы могут быть подавлены генами антиингибиторами второй генетической системы. Гены (генетические системы) выполненности соломины, происходящие от разных доноров (Апуликум 401, Um 6001, к-50989) идентичны или аллельны. Полученные результаты предполагают высокую эффективность в селекции сортов твердой пшеницы с выполненной соломиной. Многолетние исследования позволили создать новый исходный материал твердой пшеницы с выполненной соломиной, включающий следующие образцы и линии: Апуликум 401, k-49441 (Um 6001), k-50989 (Мелянопус 120-73), k-18344, k-8786, k-17386, k- 6862, 2246Б-1вс, 2246Б-7вс, 2246Б-10вс, 2111Б-1вс, 2111Б-2вс, 2111Б-3вс, 2111Б-4вс, Гордеиформе 1462Б-1, Гордеиформе 1462Б-1-1, Гордеиформе 1462Б-1-2, Гордеиформе 1398Б-12, Гордеиформе 1734, Гордеиформе 1741, Гордеиформе 1743, Гордеиформе 1744. В государственный реестр селекционных достижений РФ, включены сорта с выполненной соломиной Безенчукская 205 и Марина.
Доноры генов редукции высоты растений твердой пшеницы. Целенаправленное снижение высоты растений с применением высокоэкспрессивных генов может иметь положительный эффект на потенциал продуктивности колоса, устойчивость к полеганию, параметры ценоза и возможно в некоторых ситуациях на засухоустойчивость (McCaig T.N, Morgan J.A, 1993; Лобачев Ю.В., 2003). В засушливых регионах необходимо учитывать степень засухоустойчивости исходного материала, качественные показатели зерна и конечной продукции. В Самарском НИИСХ были получены среднерослые и низкорослые аналоги районированных или хорошо приспособленных к местным условиям сортов и изучены в многолетних экспериментах. Изученный набор генов ранжировался в порядке убывания экспрессии таким образом: Rht 14 (исходный донор итальянский сорт Castelporsiano)>Rht 1 (исходный донор мексиканский сорт Coccorit 71) >Rht Az (исходный донор образец из Азербайджана k-46828) > Rht Anh (исходный образец мексиканский сорт Anhinga). Ген Rht 14 в условиях Поволжья, наиболее сильно снижая высоту растений, негативно влиял на адаптивность короткостебельных линий (Вьюшков А.А., Мальчиков П.Н., 1997; Вьюшков А.А., 2004). Поэтому в последующем основная селекционно-исследовательская работа проводилась с генами Rht 1, Rht Az, Rht Anh. Наиболее результативной она была с генами Rht Anh и Rht 1, причем в этих экспериментах Гордеиформе 942 (низкорослый аналог Харьковской 9) по влиянию на элементы продуктивности и качество зерна селекционного материала, оказался лучшим донором гена Rht 1. Скрещивание Гордеиформе 942 / Гордеиформе 1434 позволило создать сорт Гордеиформе 1732 признанный перспективным для передачи в систему госсортоиспытания. Оценка параметров адаптивности и стабильности по А.В Кильчевскому и Л.В.Хотылевой (1997), проведенная на основе 2-х летних экспериментов в Безенчуке и в Волгограде, позволила установить превосходство Гордеиформе 1732 над стандартом Безенчукская 182 по общей адаптивной способности, стабильности и селекционной ценности генотипа. Изучение гибридов от скрещиваний Гордеиформе 1732 с высокорослыми тестерами показало промежуточное наследование высоты растений в F1 с частичным уклонением в сторону высокорослости. В F2 растения распределились по высоте на три фенотипических класса - низкорослые, среднерослые и высокорослые в соотношении 1:2:1 (2 = 1,95-2,51), что соответствует моногенной модели контроля признака. По экспрессии гена короткостебельности в сорте Гордеиформе 1732 и его родословной можно уверенно утверждать, что этот ген Rht 1.
Включение в программы скрещиваний сорта Anser 10 (CIMMYT) позволило отобрать короткостебельные и низкорослые линии, устойчивые к болезням. Скрещивание линии 646д-37 (2214Б-17 / Anser 10), принадлежащей к низкорослому морфотипу с высокорослыми сортами обнаружило промежуточный тип наследования в F1 и расщепление в F2 в соотношении 1:2:1 при 2 = 1,23, что соответствует различиям между родителями по одному гену. Отсутствие расщепления в F2 по высоте растений в популяции Гордеиформе 1732 / 646д-37, свидетельствует об аллельности генов низкорослости этих сортов. Таким образом, линия 646д-37 является носителем и донором гена Rht 1.
Скрининг по адаптивности гибридных популяций, созданных от парных скрещиваний Апуликум 943 (носитель гена Rht Anh) и сортов Безенчукская 182, Оренбургская 10, Саратовская золотистая, и др., оказался малоэффективным и не привел к созданию конкурентоспособного материала с укороченной соломиной. Успешным было скрещивание 2125Б-4 /92д-4 // Саратовская золотистая, ставшее завершающим этапом сложной ступенчатой гибридизации. Линия 2125Б-4 получена отбором из F2 Cаратовская золотистая / T.dicoccum k-46995, линия 92д-4 с геном Rht Anh, отбором из F2 cложного гибрида Безенчукский янтарь // Саратовская золотистая / T.dicoccum k-46995 /3/ Апуликум 943 / Безенчукский янтарь. Результатом всей этой работы стало включение гена Rht Anh в высокопродуктивный и адаптированный к засушливым условиям Поволжья сорт Памяти Чеховича.
Многолетнее изучение (1998-2007) сорта Памяти Чеховича в конкурсном и межстанционном сортоиспытании выявило его положительные качества. Значимое превосходство над стандартами получено по скорости развития растений (-3,7 дня относительно Безенчукской 182 по продолжительности периода всходы - колошение), К.хоз и массе 1000 зерен, достаточно четко определяется действие гена Rht Anh на длину соломины. При анализе структуры урожая и глазомерных оценках обнаружена тенденция к более высоким, чем у стандартных сортов, показателям выживаемости растений в онтогенезе и их сохранности к уборке. В эпифитотийные по листовым пятнистостям годы (1996, 1999, 2004) проявил высокую устойчивость к поражению ассимиляционной поверхности патогенами. По признакам качества зерна и макаронных изделий сорт Памяти Чеховича полностью идентичен сорту Саратовская золотистая, который является одним из лучших в России. В процессе анализа высоты растений популяций F1 - F5 гибрида Леукурум 1751 / Памяти Чеховича, установлено доминирование высокорослости. Расщепление в F2 на два фенотипических класса соответствует теоретически ожидаемому расщеплению 3:1. Все растения среднерослого класса сохранили свой фенотип в последующих поколениях до F5, что подтверждает гипотезу контроля признака среднерослости у сорта Памяти Чеховича одним рецессивным геном. Тест на аллельность этого гена и генов среднерослости мексиканского сорта Anhinga (Rht Ahn) и Гордеиформе 944 (ген Rht Az ), подтвердил гипотезу об идентичности генов среднерослости сортов Памяти Чеховича и Anhinga (Rht Anh) и неаллельности генов RhtAnh и RhtAz. Кумулятивный эффект двух генов по предварительным оценкам снижает высоту растений на 25,0-30,0 %, что открывает новые возможности для моделирования признака от низкорослого с геном Rht 1 до высокорослого морфотипов (табл. 9).
9. Модельные параметры длины соломины твердой пшеницы в Среднем Поволжье в зависимости от генетических систем и условий среды
| Морфотип | Генетическая система | Длина соломины | |||
| пределы варьирования, см | % к высокорослому морфотипу | ||||
| min | max | min | max | ||
| Низкорослый | Rht 1 | 40,0 | 66,0 | 66,6 | 60,0 |
| Среднерослый, 1-й тип | Rht Anh + Rht Az | 45,0 | 77,0 | 70,5 | 70,0 |
| Среднерослый, 2-й тип | Rht Az | 50,0 | 88,0 | 83,3 | 80,0 |
| Среднерослый, 3-й тип | Rht Anh | 55,0 | 96,0 | 90,0 | 87,0 |
| Высокорослый | нет генов системы Rht | 60,0 | 110,0 | 100,0 | 100,0 |
Сорт Памяти Чеховича обладая донорскими свойствами среднерослости (носитель гена Rht Anh) и высокой засухоустойчивости открывает перспективы селекции жаростойких, засухоустойчивых, среднерослых, потенциально высокопродуктивных сортов твердой пшеницы для засушливых регионов России, традиционно возделывающих эту культуру.
Селекционные линии 688д-4 и 653д-44, полученные с применением в качестве одного из родителей сорта Памяти Чеховича, по фенотипу отнесены к среднерослому морфотипу. Гибридологическим анализом длины соломины растений F2 от скрещиваний этих линий с высокорослым тестером сортом Жемчужина Сибири установлено действие одного рецессивного гена (расщепление 3:1 при =2,14; и 2,98). Поскольку в происхождении этих линий только сорт Памяти Чеховича является носителем гена среднерослости, то правомерно предположение, что ген среднерослости линий 688д-4 и 653д-44 идентичен гену RhtAnh
Источники и доноры устойчивости к болезням. Значительная часть исследованных в Самарском НИИСХ образцов твердой пшеницы имела умеренную устойчивость к бурой ржавчине, что, вероятно, связано с наличием в их генотипах аллелей Lr-генов. Безенчукская 182, Саратовская золотистая, Валентина и Гордеиформе 740, отнесенные к группе слабопоражающихся генотипов, входят в родословную сортов Марина и Леукурум 1750, которые в условиях эпифитотии бурой ржавчины были практически иммунными. Вполне правомерно предположение, что в данном случае проявился аддитивный эффект малых генов родительских сортов. Гибридные растения F1 от скрещивания сортов Марина и Леукурум 1750 с восприимчивым сортом Памяти Чеховича отнесены к группе восприимчивых, но со значительно меньшей степенью поражения, чем сорт Памяти Чеховича. То есть в данном случае устойчивость носит рецессивный характер или промежуточный тип наследования по количественной оценке степени поражения. В F2 этих скрещиваний наблюдались три фенотипических класса в соотношении 1:6:9, что соответствует комплементарному взаимодействию двух генов. По типу иммунности их следует отнести к рецессивным генам, по степени поражения к генам с промежуточным типом наследования признака. Следовательно сорта Марина и Леукурум 1750 являются генетическими донорами устойчивости к бурой листовой ржавчине. Оба сорта хорошо адаптированы к условиям Среднего Поволжья, потенциально продуктивные и высококачественные генотипы.
Доноры устойчивости к мучнистой росе созданы в результате интрогрессии генов в твердую пшеницу от T.diccocum (k-46995) и T.timopheevi (через T.aestivum образец ВИРа - ИТ-3). В первом случае в начале была получена устойчивая линия 2200Б-1 (Саратовская золотистая / T.dicoccum k-46995 // Cаратовская золотистая / Гордеиформе 740). Скрещивание 2200Б-1 с потенциально продуктивным, но неустойчивым сортом Гордеиформе 1434 и отбор из F3 полученной популяции привели к созданию трех устойчивых к мучнистой росе линий Гордеиформе 1727, Гордеиформе 1738 и Гордеиформе 1739. Во втором случае - устойчивый к мучнистой росе образец мягкой пшеницы 1678 Б-21(Альбидум 653*2/ ИТ-3) с транслокацией от T.timopheevi был включен в межвидовую гибридизацию с твердой пшеницей. В результате из популяции 1678Б-21 // Саратовская золотистая / Гордеиформе 740 /3/ Саратовская золотистая, методом двукратного отбора была получена устойчивая к мучнистой росе линия твердой пшеницы 9Д-2-5. Её использование в селекционном процессе привело к созданию серии продуктивных с высоким качеством зерна линий: Леукурум 1751, Леукурум 1752, Леукурум 1753, 495Д-22 и др. Если учесть, что сорта твердой пшеницы, которые привлекались в скрещивания (Саратовская золотистая и Гордеиформе 740) сильно восприимчивы к мучнистой росе, можно констатировать успешную интрогрессию устойчивости к этому патогену от T.dicoccum и T.timopheevi в твердую пшеницу.
Гибридологический анализ шести устойчивых линий (Годеиформе 1727, Гордеиформе 1738, Гордеиформе 1739, Леукурум 1751, Леукурум 1753, 495д-22) проведен на гибридах F1 – F3, полученных от скрещивания с восприимчивым сортом – тестером Памяти Чеховича. В первом поколении наблюдалось промежуточное наследование признака с уклонением в сторону устойчивого родителя. Результаты анализа характера расщепления в F2 показали, что различия между исследуемыми сортами и тестером определяются одним геном.
Среди патогенного комплекса грибов, вызывающих листовые пятнистости, поражение колоса и зерна в большинстве случаев доминировали виды Alternaria sp. в меньшей степени гельминтоспориозные грибы - Dr. teres и B.sorokiniana, наиболее вредоносное развитие этих болезней сопряжено с доминированием в популяции фузариозных и гельминтоспориозных патогенов. Необходимо учитывать, что патогены объединенные в консортные патологические системы, характеризуются значениями вредоносности отличными от вредоносности отдельных видов патогенов (Вилкова Н.А., 2000). Поэтому в селекции на устойчивость к листовым пятнистостям и фузариозу колоса необходим поиск источников и создание доноров генов устойчивости к различным видам патогенов с перспективой их рекомбинации и получения стабильно устойчивых генотипов.
Ряд образцов из коллекции ВИРа (к-63825, к-64205, к-54962, к-45267,к-49438, к-54520), CIMMYT (Anser 10, Weter-hen-3, Aufan -11, Altar 84, Shag 9, Comb –Duck 6), Космодур (Франция), Харьковская 9, Л-337 (Укр.НИИРСиГ), Саратовсая 59, Саратовская 57 (НИИСХ Юго-Востока), Гордеиформе 942, Леукурум 1675, Безенчукская 205 (Самарский НИИСХ) проявили устойчивость к листовым пятнистостям и поражению колоса в эпифитотийные для этих болезней годы (1996, 1999, 2004, 2006, 2008).
Широкое использование этого исходного материала в селекции привело к созданию устойчивых (табл. 10), к листовым пятнистостям и болезням колоса сортов, сохранявших устойчивость в условиях эпифитотии. Родословные сортов отчетливо демонстрируют стабильную в ряду поколений эффективность генов устойчивости от исходных форм, проявившуюся на естественном инфекционном фоне. Особенно высокая эффективность характерна для группы сортов, унаследовавших устойчивость от Харьковской 9 и Памяти Чеховича. Гибридизация среднеустойчивых сортов может давать трансгрессии по устойчивости. Примером могут служить сорта Памяти Чеховича и Леукурум 1675.
В предыдущий период селекции твердой пшеницы были выделены источники устойчивости к пыльной головне: Ракета, Гордеиформе 10, Безенчукская 115, Безенчукская 121, Безенчукская 139, к-6807 (ВИР), Javardo (Португалия), Um 6001 (Канада) (Шестакова А.П., Вьюшков А.А., 1974). Используя сорта Javardo и к-6807, как доноры методом беккроссов получены (рекуррентные родители Харьковская 46 и Лнукурум 163) устойчивые к головне линии: Гордеиформе 718, Гордеиформе 719 (Javardo / 3* Харьковская 46), Гордеиформе 776 (Javardo / Харьковская 46 // Безенчукская 141), Эритромелян 898 (к-6807 / 2* Харьковская 46). На фоне искусственного заражения отобрана из гибридной популяции УкрНИИРСиГ устойчивая линия Гордеиформе 724, из популяций СибНИИСХ линии Гордеиформе 747 и Гордеиформе 748.
10. Сорта, устойчивые к листовым пятнистостям, болезням колоса и доноры генов устойчивости, входящие в их родословную.
| Сорт | Исходные формы и их доля (%) в геноме сорта | Доноры генов устойчивости |
| Памяти Чеховича | Саратовская золотистая (65,6), T.dicoccum k - 46995 (15,6), Безенчук. янтарь (12,5) Харьковская 46 (5,5), Anhinga (0,8) | Харьковская 46, Anhinga |
| 653д-44 | Памяти Чеховича (50,0), Безенчукская 182(12,5), Лютесценс 1293 (12,5), Гордеиформе 740 (12,5), Саратовская золотистая (12,5) | Памяти Чеховича, |
| Горд. 1674 | Саратовская золотистая (50,0), Апуликум 341 (25,0), Гордеиформе 740 (12,5), Харьковская 9 (12,5) | Харьковская 9, Апуликум 341 |
| 688д-4 | Памяти Чеховича (50,0), Гордеиформе 1674 (50,0) | Памяти Чеховича, Гордеиформе 1674 |
| Горд. 942 | Харьк. 9 (97,75), Харьк. 46 (6,20), Coccorit 71 (0.05) | Харьковская 9 |
| Горд. 1732 | Гордеиформе 942 (50,0), Гордеиформе 1434 (50,0) | Гордеиформе 942 |
| 103д-4 | Саратовская 57 (50,0), Апуликум 341 (12,5), Саратовская золотистая (18,75), Гордеиформе 748 (6,25), Светлана (12,5) | Саратовская57, Гордеиформе748, Апуликум 341 |
| 673д-1 | Безенчукская 182 (25,0), Д2011 (25,0), 103д-4 (50,0) | 103 д-4 |
| 646д-37 | Гордеиформе 748 (25,0), ЛН-209-36 (25,0), Anser 10 (50.0) | Гордеиформе 748, ЛН-209-36, Anser 10 |
| Леук. 1675 | Гордеиформе 748 (50,0), ЛН-209-36 (50,0) | Горд.748, ЛН – 209-36 |
В последствии в процессе создания и изучения изогенных по выполненности соломины линий получены серии устойчивых к пыльной головне и с выполненной соломиной линий из популяций 2246 Б (Безенчукская 139 * 6 / Апуликум 401), 2111Б ( Безенчукская 139 * 6 / к-50989), 1462 Б (Um 6001 / 7* Безенчукская 139), где все родительские сорта были устойчивы к пыльной головне. Линия 2246Б-1, как наиболее адаптированная была привлечена в гибридизацию с сортом Безенчукская 182. Это скрещивание дало устойчивые к головне и с выполненной соломиной линии Гордеиформе 1734, Гордеиформе1741, Гордеиформе 1742, которые используются в селекции на устойчивость к пыльной головне и как доноры выполненности соломины. Создание серии сортов, проявляющих на инфекционном фоне устойчивость к пыльной головне, унаследованную от линии 2246Б-1, позволяет нам отнести ее к донорам этого признака.
На основе Гордеиформе 724 была получена линия устойчивая к пыльной головне Леукурум 1690 (Горд.724 // Саратовская золотистая / T.dicoccum k-46995). Эта линия также на основании анализа родословной и стабильно устойчивого на инфекционном фоне потомства от скрещивания Леукурум 1690 / Леукурум 1649 отнесена к генетическим донорам устойчивости к пыльной головне.
6. МОДЕЛИ СОРТОВ.
Существуют различные подходы к созданию моделей сортов. В.А.Кумаков (1985) выделил четыре методологии - обобщение опыта селекции и экстраполяция тенденций развития признаков на перспективу, статистический анализ взаимосвязи признаков, построение математических моделей продукционного процесса и, наконец, экспериментально-теоретическое обоснование значимости отдельных признаков и свойств растений, т.е. методология, основанная на физиологических исследованиях.
Развитие частной генетики вида может выдвинуть на первый план генетическую модель сорта (.Крупнов В.А, 1981, 1987; Сюков В.В., 2003; Коваль С.Ф., Коваль В.С., Чернаков В.М. и др., 2005).
Проанализировав эффективность различных методов В.А.Кумаков (1985) пришел к заключению, что физиологический метод наиболее точен, так как несет высокий уровень научной обоснованности и доказательности в поиске резервов повышения продуктивности и устойчивости растений.
Физиологический подход применяется рядом зарубежных исследовательских центров (Donald C.M., 1968; Folltyn J., 1977; Nachit M.M., Sorrells M.E. et al, 1992; Reynolds, M.P., et al, 1998; 2000).
Прикладное значение модели возрастает, если в процессе ее создания учитывались принципы физиологического моделирования, выдвинутые в качестве атрибутивных компонентов процесса (Кумаков В.А., 1985). Из пяти принципов наибольшее значение для практической селекции имеют два: принцип зональности и принцип единства в многообразии.
Эти же принципы моделирования являются фундаментальными и для развития идеи ориентации селекционного процесса по пути осознанного формирования зональной системы сортов. Повышение уровня стабильности продуктивности в системе эколого-географического ландшафта возможно только при одновременной эксплуатации целого спектра генетически разнообразных, но высокоадаптированных сортов (Жученко А.А., 1994; Романенко, А.А., Беспалова Л.А., Кудряшов И.Н., Аблова И.Б., 2005).
Вышеприведенные обстоятельства определяют конкретные задачи моделирования перспективных морфотипов. По нашему мнению модель должна в первую очередь решить две задачи: 1) выделить зональные признаки и наметить перспективы их улучшения; 2) дифференцировать сортовые популяции на биотипы в соответствии с биологическими особенностями культуры, экологическими, технологическими, агроландшафтными и т.д. нишами, конкретной эколого-географической зоны и определить для каждого биотипа адекватный уровень признаков.
Большинству изученных физиологических признаков уже дана оценка. В этом разделе, полученные результаты будут интерпретированы в комплексе с другими признаками, не затронутыми предшествующим анализом.
Для разделения сортов на биотипы необходимо выделить признаки и свойства, определяющие или вносящие в это разделение основной вклад т.е. «каркасные» признаки по которым можно легко идентифицировать тот или иной биотип. Параметры вегетационного периода являются центральным звеном – образующим кластер признаков, формирующих биотипы. Поскольку в Поволжье и на Урале нет конкурентоспособных, скороспелых сортов, – у которых колошение наступало бы на 5-7 дней раньше, чем у Безенчукской 182, обосновывать детальные параметры модели скороспелого биотипа, видимо, некорректно. Этот вывод распространяется и на среднепоздний биотип. Основным типом сортов твердой пшеницы в большинстве регионов России является среднеспелый биотип (Янченко В.И., Мельник К.М., Розова М.А., 2001; Долгалев К.М., 2005; Евдокимов М.Г., 2006). Известные в Поволжье сорта Харьковская 46, Безенчукская 139, Безенчукская 182 имеют продолжительность периода всходы-колошение при многолетнем испытании в условиях Безенчука 45-50 дней и относятся к среднеспелому биотипу. Сорта Безенчукская 205, Памяти Чеховича, Безенчукская 207 в этих же условиях колосятся за 42-47 дней. По срокам созревания различия между этими сортами либо отсутствуют, либо сорта из последней группы созревают на один день раньше. Тем не менее, мы считаем, целесообразным разделить среднеспелый биотип на три – первый или ранний по дате колошения, второй и третий по дате колошения более поздние. Различия между вторым и третьим биотипами главным образом относятся к их реакции на плодородие. Второй биотип характеризуется высокой отзывчивостью на благоприятный комплекс среды. Достаточно легко достигает уровня продуктивности 5,0 т/га и имеет повышенную устойчивость к полеганию. Сорта этого биотипа обладают хорошей засухоустойчивостью, но уступают по этому параметру сортам первого биотипа. Представители третьего биотипа лучше используют имеющиеся ресурсы минерального питания в засушливых условиях (ГТК=6-7), при низком уровне их содержания в почве и обычно имеют значительное преимущество по урожайности над сортами второго биотипа при среднепопуляционном ее варьировании от 1,0 до 2,0 т/га.
Как уже говорилось выше для всех биотипов необходимо достаточное развитие зональных признаков. По результатам оценки значимости признаков в продукционном процессе, их взаимосвязей, выявленных корреляционным и факторным анализами, нами для трех биотипов сортов определены параметры 52 признаков по отдельным из них, представлены соответствующие им гены (табл.11).
11. Модели сортов (биотипов) твердой пшеницы для острозасушливых, средних и благоприятных по увлажнению условий среды
| Признак | Единица измерения | Пределы изменчивости | Уровень связи с урожаем* гены | |||
| Биотип | ||||||
| 1 | 2 | 3 | ||||
| 1 | 2 | 3 | 4 | 5 | 6 | |
| Урожай зерна в зависимости от условий : | ||||||
| Острозасушливые ГТК = 0,4-0,5 | т/га | 0,85 0,55 | - | - | ||
| Благоприятные годы, ГТК= 1 | т/га | - | 5,5** 4,6 | |||
| Средние, ГТК =0,7-0,8 | т/га | - | - | 2,9 2,38 | ||
| Признаки ростовых и продукционных процессов по периодам онтогенеза: | ||||||
| Вынос азота всходы – кущение | мг/м2 | 380 290 | 1800 1800 | 1100 1100 | -3 | |
| Вынос Р2О5 всходы - кущение | мг/м2 | 170 140 | 700 700 | 500 500 | -3 | |
| Вынос азота кущение–трубкование | мг/м2 | 1050 670 | >7000 6150 | >3600 2930 | +3 | |
| Вынос Р2О5 в кущение – трубкование | мг/м2 | 325 210 | >2200 1920 | >1200 | +3 | |
| Вынос азота трубкование – цветение | мг/м2 | >780 500 | >5150 4515 | >2650 2150 | ||
| Вынос Р2О5 трубкование –цветение | мг/м2 | >300 175 | >1850 1590 | >970 760 | +1 | |
| Вынос азота цветение – созревание | мг/м2 | >950 590 | 6000 5390 | >3150 2564 | +2 | |
| Вынос Р2О5 цветение – созревание | мг/м2 | >640 390 | 4000 3580 | >2100 1704 | +2 | |
| Ф.ч.пр. трубкование-колошение | г/м2*сут | 4,2 4,2 | 5,5 5,5 | 4.5 4,5 | -2 | |
| Ф.ч.пр.цветение-созревание | г/м2*сут | 5,5 5,0 | 8,0 7,0 | 6,5 6,0 | ||
| Ф.ч.пр. всходы-созревание | г/м2*сут | 5,2 4,5 | 7,5 6,0 | 6,0 5,7 | ||
| Листовой индекс ГП в кущение | м2/м2 | 0,5 0,45 | 1,4 1,4 | 0,65 0,68 | +2; -3 | |
| Листовой индекс ГП в цветение | м2/м2 | 0,9 0,8 | 4,0 3,5 | >1,90 1,84 | +3 | |
| Биомасса в кущение | г/м2 | 30 30 | 70 70 | 55 55 | -3 | |
| Продолжение таблицы 11 | ||||||
| 1 | 2 | 3 | 4 | 5 | 7 | |
| Биомасса в цветение | г/м2 | 155 135 | >700 650 | 500 440 | +3 | |
| Биомасса в созревание | г/м2 | >220 180 | 1220 1150 | 725 640 | ||
| Функция роста колоса в цветение | % | 27 22 | 27 22 | 24 20 | +1 | |
| Функция роста листьев -“- | % | 25 25 | 16 16 | 19 18 | ||
| Функция роста cтебля -“- | % | 43 48 | 32 37 | 39 44 | ||
| Функция роста БП кущение | % | 8 15 | 25 35 | 20 25 | -2 | |
| Функция роста БП цветение | % | 5 5 | 25 25 | 15 18 | -2 | |
| К.хоз. | % | 37-40 30 | 45 40 | 39-40 33.2 | +1 | |
| Продолжительность периода всходы – колошение, морфологические признаки и элементы структуры урожая: | ||||||
| Количество дней до колошения | 42-45 45-50 | 45-50 45-50 | 45-50 45-50 | Vrn1, Eps | ||
| Опушение колоса | + | - | - | |||
| Окраска колоса | Белая | Белая, красная | Белая, красная | |||
| Высота растений | см | 55 60 | 60-70 90 | 65-80 80 | Rht | |
| Выполненность соломины | балл | 18-20 8-10 | 18-20 8-10 | 18-20 8-10 | SSt | |
| Отношение длины ВМУ к сумме НМУ | <1,10 1,25 | <1,05 1,15 | <1,10 1,15 | |||
| Число растений в созревание | шт/м2 | 280 280 | >300 280 | >290 290 | +3 | |
| Продуктивное кущение | 1,03 1,03 | 1,30 1,30 | >1,15 >1,15 | |||
| Доля боковых побегов в урожае | % | 3 3 | 30 25 | 15 10 | ||
| Число колосков в колосе | шт. | 10 9 | 14 13 | 12 11,5 | ||
| Озерненность колоска | шт. | 0,87 0,62 | 2,3 2,2 | 1,8 1.64 | ||
| Число зерен в колосе | шт. | 8,7 5,6 | 32,3 29,4 | 21,6 18.9 | +2 | |
| Масса 1000 зерен | г. | 37 34 | 45 43 | 41 39,6 | ||
| Устойчивость к стрессовым факторам: | ||||||
| Реакция на Puccinia reconditа*** | тип/% | 2/5 2/7 | 0-2/0-5 2/7 | 2/5 2/7 | Lr | |
| Реакция на Blumeria graminis*** | тип/% | 0/0 4/30 | 0/0 4/30 | 1-2/5 4/30 | Pm | |
| Реакция на Ustilago tritici**** | тип/% | 0 60 | 0-5 60 | 0-5 60 | ||
| Продолжение таблицы 11 | ||||||
| 2 | 3 | 4 | 5 | 6 | ||
| Устойчивость к корневым гнилям | балл | 7 6 | 7 6 | 7 6 | ||
| Устойчивость к пятнистостям | балл | 7 6 | 7 6 | 7 6 | ||
| Устойчивость к фузариозу колоса | балл | 7 5 | 7 5 | 7 5 | ||
| Засухоустойчивость | балл | 8 6 | 8 6 | 8 6 | ||
| Жаростойкость | балл | 8 6 | 7 6 | 7 6 | ||
| Устойчивость к полеганию | балл | 7 5,5 | 9 5,5 | 8 5,5 | ||
| Признаки качества зерна и макарон : | ||||||
| Натурная масса несортированного зерна | г/л | 780 780 | 780 780 | 780 780 | ||
| Содержание белка в зерне | % | 14-16 15,3 | 15,0 15,0 | 15,0 15,0 | -1 | |
| Азотный уборочный индекс | % | 60-65 55 | 63-68 62 | 62-65 61 | +2 | |
| Содержание клейковины | % | >33 33-35 | 35 35 | 35 35 | ||
| SDS седиментация | мл | >40 30 | >40 30 | >40 30 | ||
| Содержание каротиноидов | мкг % | 600 400 | 600 400 | 600 400 | Yello | |
| Прочность макарон | г/см | >900 800 | >900 800 | >900 800 | ||
| Цвет сухих макарон | балл | 5 4 | 5 4 | 5 4 | ||
*1 – первый уровень (признаки имеют стабильную связь с урожаем независимо от условий среды); 2- второй уровень (признаки связаны с урожаем в условиях засухи); 3- третий уровень (признаки связаны с урожаем в условиях благоприятного гидротермического режима). Положительный или отрицательный знак указывает направление взаимосвязи.** - в числителе параметры моделей, в знаменателе -параметры сорта Безенчукская 182;*** - естественный фон; **** - искусственный инфекционный фон.
Для некоторых признаков в моделях предполагается достижение тех значений, которые уже реализованы у лучших сортов, но они распределены между ними и при этом не являются взаимно компенсаторными. Обязательным атрибутом сортов первого биотипа является высокая засухоустойчивость, устойчивость к пыльной головне, листовым пятнистостям, мучнистой росе и хлебному пилильщику, толерантность или слабая восприимчивость к бурой ржавчине. Для высоты растений и ее структуры оптимальными для первого биотипа являются значения, обеспечиваемые геном редукции высоты растений Rht Anh (доноры сорта Памяти Чеховича, 688д-4, 653д-44) (минимальный уровень) и параметры среднерослого сорта Безенчукская 205 (максимальный уровень). То же относится и к признаку «длина верхнего междоузлия» и его отношению к длине всех нижележащих междоузлий. Высота растений второго биотипа, может быть снижена в результате действия гена Rht 1 и комплементарного эффекта генов Rht Anh и Rht Az еще больше. Количественные признаки в зависимости от условий среды по результатам факторного анализа, как показано выше, образуют три кластера (уровня) взаимосвязи с урожайностью. Исходными данными для расчетов модельных значений этих признаков служили коэффициенты факторных нагрузок после их вращения методом варимакс. Базовой величиной считали прирост урожайности модельного объекта от уровня стандарта. Для сухих лет эта величина рассчитывалась по значению сорта Безенчукская 182 в 1998 году, для влажных в - 1997 году и для средних в - 2001году. В расчет брались только те признаки, которые имели значимую связь с урожайностью. Они увеличивались пропорционально увеличению урожайности. Отрицательно связанные с урожаем признаки изменяли незначительно, исходя из физиологической целесообразности, или оставляли на уровне стандарта (например, содержание белка в зерне). Предлагаемые модели не являются окончательными, для их оптимизации прежде всего требуется расширение спектра биотипов, вычленение и оценка перспектив развития признаков адаптивности комплементарных условиям конкретных мега- микрозон и возможностей их сочетания со свойствами сортов широкого ареала, повышение степени адекватности региональным ресурсосберегающим технологиям.
7. МЕТОДЫ ПОДБОРА РОДИТЕЛЬСКИХ КОМПОНЕНТОВ ДЛЯ ГИБРИДИЗАЦИИ И СПОСОБЫ РАБОТЫ С СЕЛЕКЦИОННЫМ МАТЕРИАЛОМ.
Подбор пар для гибридизации. Для устойчивой эволюции селекционного материала по пути повышения продуктивности и адаптивности селекционеру необходимо иметь достаточно представительную коллекцию базовых генотипов, обладающих высоким уровнем общего гомеостаза. Принципиально новые базовые генотипы, чаще всего характеризуют определенный этап в селекции, являются его наиболее яркими представителями и появляются в результате трансгрессий по признакам продукционного процесса. Соответствующий принцип подбора родительских компонентов для гибридизации обоснован Ю.Б.Коноваловым и Т.И.Хупацария (1996). Примером его реализации является получение сорта Безенчукская 182, который унаследовал оптимальный тип ростовых процессов от Безенчукской 105, а потенциал продуктивности колоса, видимо, от Харьковской 46.
Другим примером трансгрессии по признакам продукционного процесса является сорт Памяти Чеховича – продукт межвидовой, сложной ступенчатой гибридизации. При его создании проведено целенаправленное снижение высоты растений (ген Rht Anh) и введение на этапах сложной ступенчатой гибридизации от образца Tr.dicoccum k–46995 признака синхронности в развитии главного и боковых побегов. Эти признаки и генетический фон базового генотипа засухоустойчивого сорта Саратовская золотистая позволили повысить жаростойкость процессов гаметогенеза и крупность зерна, что обеспечивает ему в условиях засухи преимущество по числу зерен в колосе и на 1м2 посева, в благоприятные по увлажнению годы по массе зерновки и позволяет стабильно формировать посев с высоким выходом зерна из надземной биомассы ( К.хоз.). Генетико-статистическими исследованиями по диаллельной модели Хеймана установлена высокая наследуемость количественных признаков адаптивности (жаростойкость) и продуктивности сорта Памяти Чеховича, что увеличивает его надежность как базового генотипа.
Учитывая ограниченные возможности рекомбинационного процесса в парных (А / В) и сложных (А / В //С; А /В // С / Д) скрещиваниях эффективным остается метод ступенчатой гибридизации. Самая эффективная и надежная концепция гена может применяться только при наличии доноров необходимых генов. Работа по их созданию требует времени и дополнительных усилий. Между тем коллекция твердой пшеницы и генетически родственных ей видов отличается большим разнообразием. Поэтому концепции сорта и признака будут ещё длительное время востребованы в селекции твердой пшеницы. Если высокий уровень неспецифического гомеостаза базового генотипа рассматривать как сложный признак, то концепция использования базового генотипа объединяет два принципа подбора сортов для гибридизации – принцип гена и принцип признака.
В настоящее время Самарский НИИСХ располагает адаптированными к условиям Поволжья донорами генов редукции высоты растений, устойчивости к популяциям патогенов мучнистой росы, бурой ржавчины, пыльной головни, листовых пятнистостей, высокой температуре, высокого содержания каротиноидных пигментов в зерне и выполненности соломины, обеспечивающей устойчивость к хлебному пилильщику.
Таким образом, всю совокупность подходов к подбору родительских пар целесообразно использовать системно в зависимости от задач, селектируемых признаков и условий среды. Основываясь на многолетних данных, предлагается следующая система создания и отбора родительских генотипов. Она состоит из двух этапов.
Первый включает:
- создание базовых генотипов – используются принципы подбора пар по комплементарности продукционных процессов, эколого-географически отдаленные скрещивания, отдаленная гибридизация; предусматривает получение трансгрессий усиливающих общий гомеостаз формирования урожайности и потенциальную продуктивность; вновь созданные базовые генотипы могут иметь коммерческое значение;
- создание адаптированных доноров хозяйственно-ценных признаков; предусматривает широкое изучение новых отечественных и зарубежных сортов, коллекции твердой пшеницы и других видов из рода Triticum, применение метода беккроссов, сложной ступенчатой гибридизации при интрогрессии генов из других эколого-географических групп и видов; аналоги базовых генотипов могут иметь коммерческое значение.
Второй этап:
- включение в гибридизацию сортов из групп базовых генотипов, адаптированных доноров, аналогов базовых генотипов с интрогрессированными генами, сортов из коллекции; обязательным атрибутом является создание коммерческих сортов.
Отбор в гибридных популяциях и селекционных питомниках. Эффективность отбора из гибридной популяции и в селекционных питомниках во многом зависит от условий среды, в которых выполняется селекционный процесс. Нельзя или почти невозможно, если исключить случайности, создать сорт устойчивый к засухе, температурным стрессам, болезням, высокоурожайный, отзывчивый на агрофон и т.д., проводя селекцию в неадекватных условиях. Поэтому в селекции придается большое значение фону, на котором проводится отбор (Мичурин И.В., 1955; Frey K.J., 1964; Драгавцев В.А. и др., 1984; Жученко А.А., 2001; Драгавцев В.А., 2005). По способности выявлять изменчивость Е.Н.Синская (1963) подразделила фоны на три группы: анализирующий, или способствующий проявлению изменчивости; стабилизирующий, в котором полиморфизм не проявляется, но происходит закрепление достигнутых отбором изменений на предыдущем фоне; нивелирующий фон – угнетающий жизнеспособность всех биотипов в популяции, или нивелирующий различия между ними по другим причинам. На основании этих идей белорусскими учеными (Кильчевский А.В., Хотылева Л.В., 1985; Кильчевский А.В., 1986) разработаны математические методы оценки селекционных фонов или способы их классификации. Мы использовали данный подход, который основан на модели двухфакторного дисперсионного анализа для оценки дифференцирующей способности среды в двух экспериментах, в которых получены значимые эффекты взаимодействия генотип-среда.
Первый включал период 1998-2002 годы, в течение которого изучены восемь сортов. С 1998 по 2000 годы исследования выполнялись параллельно по двум фонам – пару и зерновому предшественникам (овес на зерно). В 2001 году эти же сорта, кроме зернового предшественника, высевались по пару в трех вариантах, которые дифференцировались в зависимости от агрофона почвенного участка. Второй эксперимент проведен в 2005-2006 годах в шести средах - в Безенчуке – по двум предшественникам (пар, овес на зерно) и в Волгограде (Нижневолжский НИИСХ) в оба года по пару. Изучалось 30 генотипов, представленных сортами конкурсного сортоиспытания. В обоих экспериментах изучался один признак – урожайность зерна с единицы площади.
В первом эксперименте между эффектами сред и их дифференцирующей способностью выявлена определенная зависимость. Наибольший полиморфизм наблюдался на продуктивных фонах (три фона по пару, один по зерновому предшественнику). Здесь же сильнее проявились эффекты дестабилизации ( Kek равен 2,90; 2,58; 2,41; и 1,49). Эти среды согласно классификации Е. Н.Синской (1958) можно отнести к анализирующим фонам. Все остальные фоны попали в класс стабилизирующих. Три среды две из которых представляли засушливый 1998 год, одна низкопродуктивный фон 2001 года, отличаются самыми неблагоприятными условиями (dk < -7,5), невысокими значениями 2 ДССk, сильным эффектом компенсации (Kek= 0,22-0,52), но относительная дифференцирующая способность этих сред (Sek =21,4-22,7) самая высокая в эксперименте. Это означает, что в данных средах, исследуемые популяции генотипов имеют достаточный полиморфизм для эффективного стабилизирующего отбора.
Во втором эксперименте максимальную урожайность генотипов обеспечила среда по чистому пару в 2006 г., минимальная получена в Врлгограде в 2005году. Однако максимальная дифференцирующая способность (2 ДССk =17,51) и максимальный дестабилизирующий эффект (Kek=9,41) проявились в Волгограде в 2005 году.
Если, используя результаты двух экспериментов, сравнить два предшественника – пар и зерновые по потенциальной способности дифференцировать сортовые популяции и обеспечивать эффективный отбор, то очевидно, что паровому предшественнику следует отдать предпочтение - пять анализирующих фонов из шести формирует пар и один зерновой предшественник. Однако коэффициент типичности среды (tk) в большинстве сравнений (5 из 6) был выше по зерновому предшественнику. По коэффициенту предсказуемости (Рk) это соотношение было одинаковым (3/3).
Наибольшая дифференцирующая способность малопродуктивных фонов, обнаруженная в условиях Волгограда, определяется лимитирующими факторами среды, действовавшими в экологических пунктах. В Волгограде средняя температура периода гаметогенеза была выше аналогичного показателя в Безенчуке в 2005 году на 2,6 С0, в 2006 году на 2,8С0. Учитывая высокую предсказуемость среды г.Волгограда (Рk = 0,21-0,39) и превалирование аддитивных эффектов генов, контролирующих жаростойкость процессов гаметогенеза и формирования зерна, определяющих величину признака «число зерен в колосе» - обеспечивающего полиморфизм в популяции, вполне реально включение этого пункта в систему совокупности сред для целенаправленной селекции. Особенно заметно преимущество отбора в условиях Волгограда из популяций, в которых одним из компонентов скрещивания был жаростойкий сорт Памяти Чеховича.
Совокупность изученных нами сред - Безенчук пар (БП), Безенчук зерновой предшественник (БЗ), Волгоград пар (ВП) по общей адаптивной способности (ОАСg) имеет следующую направленность экологического вектора БП БЗ ВП, по вариансе дифференцирующей способности среды (2ДССk) и по относительной дифференцирующей способности (Sek) ВП БП БЗ. В связи с этим целесообразно применение следующей схемы использования совокупности сред: репродукция F1 – F3 и отбор элитных колосьев в среде (ВП), испытания в селекционных и контрольном питомниках в среде (БП), испытания предварительное, конкурсное в средах ( БП, ВП, БЗ).
Результативным может быть отбор, проведенный как в ранних, начиная с F2, так и в поздних поколениях. В F2 отобраны - Безенчукская степная и Памяти Чеховича, в F4 - Безенчукская 205 и Марина, F6 – Безенчукская Нива. Технология селекционного процесса в лаборатории предусматривает в качестве обязательного приема отбор в ранних поколениях. При малочисленной популяции F2 ее размножение в следующих поколениях сопровождается увеличением числа отбираемых элитных колосьев. Отбор растений осуществляется на уровне растений, элитного колоса, эффективным может быть отбор только по признакам, контролируемым небольшим числом генов: высота растений, выполненность соломины, устойчивость к бурой ржавчине, мучнистой росе, окраска, опушение колоса, наличие желтого пигмента в зерне.
При изучении селекционного материала в гнездовом посеве (СП-1 года) довольно четко удается идентифицировать высокопродуктивные, приспособленные к данным условиям среды популяции, дающие большой выход ценных селекционных форм. Оценка перспективности популяций, выбор лучших из них по величине надземной массы, массы зерна, К.хоз и коэффициенту их вариации, позволяет селекционеру сконцентрировать усилия и повысить эффективность селекции. Малопродуктивные популяции прорабатываются с максимальным давлением отбора или бракуются полностью. Большой объем отборов из таких популяций возможен, если поставлена цель получения промежуточных форм для ступенчатой гибридизации. Чаще такие ситуации возникают при интрогрессии генов от других видов или от неадаптированного исходного материала.
Вероятность отбора генотипов адаптированных к конкретным условиям года, экопункта, превышающих среднепопуляционный уровень продуктивности на 20,0%, при бесповторном посеве на делянках 1,0-10,0м2, составляет 90,0%. Генотипы с превышением среднепопуляционного значения продуктивности в пределах 10-15%, могут с большой долей вероятности быть утеряны. Процедуры оценки и отбора в СП-2 и СП-3 годов одинаковы. На этих этапах достаточно хорошо идентифицируется морфотип (высота растений, вегетационный период, динамика кущения), устойчивость к болезням листьев, колоса, полеганию, выживаемость растений, озерненность колосьев, устойчивость к засухе и высоким температурам, морфологическая и ботаническая однородность агроценоза, выполненность и стекловидность зерна, степень развития черного зародыша.
Данные по урожайности и ее элементам в контрольном питомнике хорошо коррелируют с аналогичными показателями конкурсного сортоиспытания. К полевым и лабораторным оценкам на предыдущем этапе добавляются технологические исследования сортов по содержанию каротиноидов в зерне и реологическим свойствам клейковины, оцениваемым по показателю SDS седиментации.
Длительность изучения в предварительном сортоиспытании (ПСИ) 1 или реже 2 года, в конкурсном сортоиспытании (КСИ) 3 или 4 года, что позволяет достаточно точно оценить весь спектр урожайных и адаптивных возможностей сорта. Система межстанционного сортоиспытания дополняет, полученную информацию. На этапе ПСИ и КСИ можно довольно точно контролировать урожайность по элементам структуры и описывать ее формирование в зависимости от сорта, условий года и предшественника. Чистый пар хорошо дифференцирует сорта по признакам «К.хоз» и «масса 1000 зерен», предшественник «овес на зерно» - по числу колосков и зерен в колосе. Кроме того, в этих питомниках подробно изучаются параметры качества зерна и макарон. Полный анализ включает оценку физических качеств зерна, содержание белка, клейковины, каротиноидов, качества клейковины и макаронных изделий. Многолетнее изучение позволяет отбирать для передачи в систему государственного сортоиспытания сорта, стабильно формирующие высококачественное зерно и макароны.
8. Результаты селекции яровой твердой пшеницы и экономическая эффективность возделывания новых сортов.
За годы селекционной работы (1989-2009гг.) было создано и передано в систему Грсударственного сортоиспытания 8 сортов из них 4 включены в Государственный реестр селекционных достижений, допущенных к использованию, два сорта продолжают изучаться (табл.12). Для экономической оценки возделывания сортов яровой твердой пшеницы в Среднем Поволжье новые сорта были объединены в две группы. В первую группу включили группу сорта с выполненной соломиной - Безенчукская 205 и Марина, во вторую наиболее продуктивные высококачественные современные сорта с полой соломиной - Безенчукская степная, Безенчукская Нива.
Возделывание в Среднем Поволжье сортов твердой пшеницы с выполненной соломиной, устойчивых к хлебному пилильщику, может быть оправдано экономически, так как исключает снижение урожайности посевов и необходимость их обработки инсектицидами в случае эпизоотии вредителя.
В основу расчетов положены затраты возделывания твердой пшеницы по ресурсосберегающим технологиям с внесением удобрений в действующем веществе N30P30 и c применением гербицидов в фазу кущения. Уровень урожайности современных сортов при их возделывании по зерновым предшественникам и вышеприведенной технологии в большинстве экспериментов варьирует в пределах 15,0 –25,0 ц/га (А.П.Чичкин, В.А.Корчагин и др., 2005). Эффект от возделывания сортов с выполненной соломиной рассчитывали при 10% и 20% коэффициента вредоносности.
12. Сорта твердой пшеницы, созданные в 1989-2008гг.
| Сорт | Доля авторства, % | Годы Госсортоиспытания | Год включения в Госреестр РФ |
| Безкечукская 200 | 40,0% | 1999-2001 | 2002, 7,9 рег. |
| Безенчукская степная | 40,0% | 2001-2003 | 2004, 7,8,9 рег. |
| Безенчукская 202 | 40,0% | 2002-2004 | |
| Памяти Чеховича | 45,0% | 2003-2005 | |
| Безенчукская 205 | 50,0% | 2005-2007 | 2008, 7,8,9 рег. |
| Марина | 50,0% | 2005-2008 | 2009, 7,9 рег. |
| Безенчукская 207 | 50,0% | 2008-2009 | |
| Безенчукская Нива | 50,0% | 2009 |
В условиях с потенциалом урожайности 1,5т/га при коэффициенте вредоносности хлебного пилильщика 10,0% чистый доход с 1 га посевов снижается с 3,0 тыс. руб. до 2,25тыс. руб., а рентабельность с 50% до 38,5%.
В тех же условиях, но при коэффициенте вредоносности хлебного пилильщика 20,0%, величина чистого дохода составляет 1,5 тыс. руб., т.е. снижается вдвое, а рентабельность падает до 12,5%.
В условиях с потенциалом урожайности 2,5т/га и коэффициентах вредоносности 10,0% и 20,0% чистый доход с 1 га снижается с 6,75 тыс. руб. до 5,55 и 4,35 тыс. руб., рентабельность с 81,8% до 69,8% и 56,9% соответственно коэффициентам вредоносности.
Такие же расчеты были проведены для сортов Безенчукская степная и Безенчукская Нива при этом мы опирались на те же исходные экономические параметры, что и в предыдущем экономическом анализе эффективности возделывания сортов с выполненной соломиной. В качестве стандарта был взят сорт Безенчукская 182. Учитывая, что качество зерна и макарон новых сортов выше, чем у стандарта за основу при расчете экономической эффективности были использованы различия по урожайности, полученные в конкурсном сортоиспытании в 2005-2008 годы.
Приведенные расчеты показывают высокую эффективность возделывания новых сортов – чистый доход для сорта Безенчукская степная в расчете на 1000 га увеличивается до 340 тыс. рублей рентабельность на 2,0%, и для сорта Безенчукская Нива - 1 млн.480 тыс. рублей и 9,9% соответственно..
Учитывая, что экономические эффекты от качества зерна, которое по нашим оценкам выше у новых сортов, в расчеты не включены, эти цифры следует считать как минимально возможный экономический эффект от внедрения новых сортов в сельскохозяйственное производство
ВЫВОДЫ
1. На основе анализа многолетних данных вредоносности биотических и абиотических стрессоров актуальными в селекции на адаптивность признаны следующие направления (в порядке убывания значимости):
- жаростойкость процессов гаметогенеза и образования зерна
- устойчивость ростовых процессов к обезвоживанию растительных тканей и активность корневой системы
- устойчивость к листовым пятнистостям и корневым гнилям (Fusarium, Helminthosporium), мучнистой росе (Blumeria graminis) и бурой ржавчине (Puccinia recondita)
- устойчивость к полеганию.
2. Динамика листовой поверхности в онтогенезе твердой пшеницы в зависимости от сорта и условий среды имеет следующие особенности:
- в благоприятных по увлажнению условиях сорта всех периодов селекции развивают максимальную площадь листьев 4,0-5,0 м2/м2, что считается оптимальной величиной для зерновых культур. Ф.ч.пр. у лучших сортов превышает 8,0 г/м2*сут., полегание и болезни снижают эту величину до 2,0 г/м2*сутки и ниже;
- продуктивные сорта отзываются на улучшение условий среды увеличением площади листьев главных побегов в период трубкование – цветение - формирование зерна, чрезмерного увеличения листовой поверхности у современных сортов не наблюдается; в условиях сильной засухи положительное значение имеет увеличение площади листьев в период от трубкования и до созревания; повышенный ассимиляционный потенциал сортов имеет значение для увеличения зерновой продуктивности при условии устойчивости морфогенетических процессов (формирование колоса, гаметогенез, образование зерна, аттрагирующая способность колоса) к высоким температурам и другим стрессовым факторам
- засухоустойчивость оказывает определяющую роль на продолжительность активного функционирования ассимиляционного аппарата, не исключается контроль этого свойства сортов функциональным состоянием колоса;
- площадь листьев боковых побегов не имеет определяющего значения в формировании генотипических различий по урожайности, как боковых побегов, так и в целом посева с единицы площади.
3. Селекцией в Среднем Поволжье на разных её этапах реализованы следующие варианты ростовых функций в онтогенезе твердой пшеницы:
- значимое превалирование в ростовых процессах боковых побегов с момента начала их роста и независимо от конкретной динамики лимитирующих факторов среды;
- доминирование в ростовых функциях главного побега;
- значительное увеличение доли колоса в ростовых функциях современных продуктивных сортов от момента его формирования и до завершения роста стебля (цветение) с пролонгацией этого процесса до восковой спелости зерна;
- общий вывод, вытекающий из результатов изучения ростовых функций, состоит в том, что успешным может быть только тот селекционный сдвиг (трансгрессия) по относительному увеличению колоса, который скоррелирован с адекватным уровнем адаптивности.
4. Значительная реутилизация структурных ассимилятов наблюдается у всех сортов в условиях сильного полегания посевов и у незасухоустойчивых сортов в условиях сильной засухи; процесс селекции на продуктивность и адаптивность не привел к увеличению объёмов и доли реутилизации в урожае зерна.
5. Использование минеральных элементов в онтогенезе сортов связано со следующими процессами:
- динамика ежесуточного выноса азота сортами разной продуктивности соответствует приростам их надземной биомассы при этом основное преимущество интенсивных сортов по выносу в большинстве ситуаций наблюдается в предрепродуктивную и репродуктивную фазы.
- эффективность использования в ростовых процессах поглощенного азота определяется генетическими системами засухоустойчивости; отзывчивость продуктивных сортов во влажные годы обеспечивается высоким валовым выносом и потреблением минеральных веществ, что определяется уровнем адаптивности и продуктивности; фосфор в ростовых процессах надземной биомассы используется с одинаковой эффективностью всеми сортами независимо от условий среды.
- современные сорта твердой пшеницы с высокой эффективностью используют естественное плодородие почвы, но не обладают специфическими генными системами отзывчивости ростовых процессов на повышенный уровень питания и удобрения; поиск таких генетических систем – актуальная научно-исследовательская задача.
6. Взаимосвязь признаков продукционного процесса и элементов урожайности имеет следующие закономерности:
- стабильная взаимосвязь урожайности зерна агроценоза в контрастных условиях среды наблюдается с процессами морфогенеза колоса (функция роста колоса в цветение), К.хоз. и выносом фосфора в период трубкование - цветение.
- в благоприятных условиях наиболее тесно связан с урожаем зерна признак «число растений на 1м2», в условиях засухи - число зерен в колосе. Масса 1000 зерен и урожайность зерна агроценоза имеют независимый тип генотипического варьирования.
- признак «число зерен в колосе» стабильно и положительно коррелировал с выносом азота и фосфора в период цветение – созревание зерна; в связи с этим можно предположить, что отбор форм с высокоозерненным, крупным колосом опосредовано приводит к увеличению выноса макроэлементов в репродуктивный период и усилению мощности и активности корневой системы.
- содержание белка в зерне и урожайность зерна агроценоза являются взаимно компенсаторными признаками; стабилизация содержания белка в зерне в процессе селекции высокоурожайных адаптированных сортов может быть достигнута путем увеличения АУИ, который положительно связан с выносом азота и фосфора в период трубкование - цветение.
- сильный негативный эффект на урожайность, число зерен в колосе, К.хоз. и функцию роста стебля вносит функция роста боковых побегов к моменту цветения.
7. Процесс селекции твердой пшеницы в Среднем Поволжье сформировал коадаптированные блоки генов, которые, образовавшись в ранний период селекции, не только сохраняются, проходя циклы рекомбинации и отбора, но и усложняются, вбирая в себя новые потоки генетической информации; первый такой блок генов возник в результате межвидового скрещивания местных образцов твердой и мягкой пшеницы, затем значительное его обогащение произошло в результате включения генов харьковского и саратовского пулов.
8. Разработаны основные параметры трех моделей сортов среднеспелого биотипа яровой твердой пшеницы для Средневолжского региона
9. Реализация признаков моделей достигается применением выявленных и созданных источников и доноров генов:
- засухоустойчивости, жаростойкости, с повышенным уровнем адаптивной способности и селекционной ценности: Безенчукская 182, Безенчукская степная, Памяти Чеховича, Безенчукская 205, Марина, Безенчукская 207, Леукурум 1751, Безенчукская Нива, Гордеиформе 1732, в том числе базовых генотипов, несущих гены гомеостатичности и блоки коадаптированных генов – Безенчукская степная, Памяти Чеховича, Безенчукская 205;
- числа зерен в колосе – Безенчукская 182, Безенчукская степная, Безенчукский янтарь, Безенчукская 205, Марина, Памяти Чеховича, Безенчукская 207, Жемчужина Сибири, Валентина, Безенчукская Нива, Леукурум 1751, Леукурум 1753, Леукурум 1756, 688д-4, 653д-58, в том числе доноры генов признака в условиях высокотемпературного стресса – Памяти Чеховича и Безенчукская 205;
- выхода зерна из надземной биомассы (К.хоз ) – Памяти Чеховича, Безенчукская степная, Безенчукская 182, Марина, Безенчукская Нива, Гордеиформе 1732, Безенчукская 207, Омская янтарная, 688д-4, 653д-44, в том числе доноры генов признака в широком диапозоне сред - Памяти Чеховича, Безенчукская степная, Безенчукская 205;
- массы 1000 зерен – Саратовская золотистая, Валентина, Краснокутка 10, Краснокутка 12, Марина, Безенчукская 205, Безенчукская степная, Памяти Чеховича, Безенчукская Нива, в том числе доноры генов признака – Безенчукская степная, Безенчукская 205, Памяти Чеховича;
- натурной массы зерна: Валентина, Марина, Краснокутка 10, Безенчукская Нива, Безенчукская степная, Безенчукский янтарь;
- содержания белка, клейковины, урожайности и К.хоз: Памяти Чеховича, Безенчукская степная, Безенчукская 182;
- реологических свойств теста: Харьковская 9, Харьковская 13, Харьковская 15, WSMP 13, Vic, Гордеиформе 740, Гордеиформе 942, Гордеиформе 1732, Саратовская золотистая, Валентина, Ник, Безенчукская Нива, Марина;
- прочности макарон - Безенчукская 207, Памяти Чеховича, Безенчукская степная, Безенчукская 200, Марина, Безенчукская Нива, Саратовская золотистая, Гордеиформе 1732, Безенчукская 182;
- высокого содержания каротиноидных пигментов в зерне – Саратовская золотистая (2 гена –Yellow 1, Yellow 2), Памяти Чеховича (предположительно гены-Yellow 1, Yellow 2), Безенчукская 200 (Yellow 1), Гордеиформе 1434 (Yellow 2), Леукурум 1675 и ЛН-209-36 (ген или блок генов неаллельный Yellow 2);
- золотисто-желтого цвета макарон – Безенчукская 207, Безенчукская Нива, 653д-44, Саратовская золотистая;
- низкой потери сухих веществ при варке макарон – Памяти Чеховича, Саратовская золотистая, Валентина;
- устойчивости к болезням: Puccinia reconditа, Blumeria graminis и листовым пятнистостям – всего 78 образцов T.durum, T.dicoccum, T.cartlicum и T.timopheevi из коллекции ВИР, в том числе созданные в Самарском НИИСХ - Гордеиформе 1727, Гордеиформе 1738, Гордеиформе 1739, Леукурум 1751, Леукурум 1753, 495д-21, 495д-22, 495д-7, Безенчукская 205 доноры генов устойчивости к Blumeria graminis; Марина, Леукурум 1750 -доноры генов устойчивости к Puccinia reconditа; Леукурум 1690, Гордеиформе 1734, Безенчукская 205 - доноры генов устойчивости к Ustilago tritici; Памяти Чеховича, Гордеиформе 1674, 103д-4 - доноры генов устойчивости к листовым пятнистостям, болезням колоса;
- редукции высоты растений: Гордеиформе 1732, 646д-37 – ген Rht -1; Памяти Чеховича, 688д-4, 653д-44, 653д-58 – ген Rht Anh; Гордеиформе 944, Гордеиформе 1172 – ген Rht Az;
- выполненности соломины (SSt гены): Апуликум 401, k-49441 (Um 6001), k-50989 (Мелянопус 120-73), k-18344, k-8786, k-17386, k- 6862, 2246Б-1вс, 2246Б-7вс, 2246Б-10вс, 2111Б-1вс, 2111Б-2вс, 2111Б-3вс, 2111Б-4вс, Гордеиформе 1462Б-1, Гордеиформе 1462Б-1-1, Гордеиформе 1462Б-1-2, Гордеиформе 1398Б-12, Гордеиформе 1734, Гордеиформе 1741, Гордеиформе 1743, Гордеиформе 1744, Безенчукская 205, Марина;
10. Обоснована система создания и отбора родительских генотипов, состоящая из двух этапов - первый включает: а) создание базовых генотипов – используются принципы подбора пар по комплементарности продукционных процессов, эколого-географически отдаленные скрещивания, отдаленная гибридизация - предусматривают получение трансгрессий усиливающих общий гомеостаз формирования урожайности и потенциальную продуктивность; вновь созданные базовые генотипы могут иметь коммерческое значение; б) создание адаптированных доноров хозяйственно-ценных признаков - предусматривает широкое изучение новых отечественных и зарубежных сортов, коллекции твердой пшеницы и других видов из рода Triticum, применение метода беккроссов, сложной ступенчатой гибридизации при интрогрессии генов из других эколого-географических групп и видов; аналоги базовых генотипов могут иметь коммерческое значение; второй этап включает: а) использование в гибридизации сортов из групп базовых генотипов, адаптированных доноров, аналогов базовых генотипов с интрогрессированными генами, сортов из коллекции; б) обязательным атрибутом является создание коммерческих сортов.
11. Для оценки гомеоадаптивности сортов при испытаниях в различных средах наиболее целесообразно применение комплекса параметров А.В.Кильчевского, Л.В. Хотылевой (1997).
12. Совокупность изученных сред (БП, БЗ, ВП) имеет следующую направленность экологического вектора - по общей адаптивной способности (ОАСg) БП БЗ ВП, по дифференцирующей способности среды (2ДССk, Sek) ВП БП БЗ; в связи с этим целесообразно применение следующей схемы использования совокупности сред: репродукция F1 – F3 и отбор элитных колосьев в среде (ВП), испытания в селекционных и контрольном питомниках в соеде (БП), параллельное изучение сортов в питомниках предварительного и конкурсного испытаний в средах (БП, ВП, БЗ).
13. В условиях Безенчука, отбор из расщепляющихся поколений при достаточном объёме популяции целесообразно начинать в F2, ориентируясь на признаки, контролируемые небольшим числом генов и предпочитая хорошо озерненные, продуктивные колосья.
14. Созданы сорта, включенные в Государственный реестр селекционных достижений допущенных к использованию в РФ Безенчукская 200 (7и 9 регионы), Безенчукская степная (7,8,9 регионы), Безенчукская 205 (7,8,9 регионы), Марина (7,9 регионы). Безенчукская 207 и Безенчукская Нива проходят Госиспытание.
15. Возделывание новых сортов в зависимости от агроэкологической ситуации дает дополнительный доход по минимальным оценкам от 0,4 до 2,4 тыс. рублей на 1га посева из расчета сложившегося в 2008 году паритета цен на производство и реализацию зерна твердой пшеницы.
ПРЕДЛОЖЕНИЯ ДЛЯ ПРАКТИЧЕСКОЙ СЕЛЕКЦИИ И ПРОИЗВОДСТВА
- Использовать в практической селекции в Среднем Поволжье и на Урале, источники и доноры генов хозяйственно-ценных признаков, выделенные и созданные в процессе исследований.
- Целенаправленно, на основе засухоустойчивых, адаптированных к условиям Поволжья доноров генов редукции высоты растений – Гордеиформе 1732, 646д-37 (Rht-1), Памяти Чеховича, 688д-4, 653д-44, 653д-58 (Rht Anh), приступить к формированию сортов низкорослого и среднерослого морфотипов.
- Широко использовать в селекционной работе параметры разработанных моделей сортов.
- Интенсивно использовать в производстве высококачественного зерна сорта, включенные в Государственный реестр селекционных достижений – Безенчукская 200, Безенчукская степная, Безенчукская 205, Марина.
Список работ, опубликованных по теме диссертации
Монографии
- Вьюшков, А.А. Селекционно-генетическое улучшение яровой пшеницы / А.А. Вьюшков, П.Н. Мальчиков, В.В. Сюков, С.Н. Шевченко.- Самара, 2008.- 536 с.: табл., рис.+прил.- Библиогр.: с.534.. Мальчиков П.Н.
- Мальчиков П.Н. Формирование моделей сортов твердой пшеницы для Средневолжского региона России / П.Н.Мальчиков, А.А.Вьюшков, М.Г.Мясникова.- Самара. табл. рис.112с. (в печати).
Статьи, авторские свидетельства, патенты
- Мальчиков, П.Н. Продолжительность вегетационного периода как критерий адаптивности твердой пшеницы в Среднем Поволжье / П.Н. Мальчиков // Итоги и перспективы исследований в области селекции, семеноводства и ландшафтно-экологического земледелия: тез. докл. науч.-практ. конф., посвящ. 85-летию НИИСХ Юго-Востока и 100-летию … В.Н. Мамонтовой, 27-28 июня 1995 г.- Саратов, 1995.- С.50-52: табл.
- Вьюшков, А.А. Селекция яровой мягкой и твердой пшеницы / А.А. Вьюшков, В.В. Сюков, П.Н. Мальчиков // Вопросы повышения устойчивости зернового хозяйства в условиях Поволжского региона: тез. докл. науч.-практ. конф. Секция 1 / Поволжский НИИСС.- Кинель, 1997. - С.17-19: табл.
- Вьюшков, А.А. Создание и результаты изучения аналогов твердой пшеницы, различающихся по аллелям системы локусов Rht / А.А. Вьюшков, П.Н. Мальчиков // Развитие научного наследия академика Н.И.Вавилова: тез. междунар. науч. конф. / Саратовская ГСХА.- Саратов, 1997. - Ч.1.- С.42-45.
- Вьюшков, А.А. Результаты и перспективы селекции твердой пшеницы на адаптивность и качество в Самарском НИИСХ / А.А. Вьюшков, П.Н. Мальчиков, М.Г. Мясникова // Проблемы увеличения производства и повышения качества зерна в Российской Федерации: тез. докл. науч.-практ. конф…, 2-3 июля 1997 г. / НИИСХ Юго-Востока. - Саратов, 1997. - С.39-40.
- Мальчиков, П.Н. Наследование каротиноидных пигментов в зерне твердой пшеницы / П.Н. Мальчиков // Проблемы увеличения производства и повышения качества зерна в Российской Федерации: тез. докл. науч.-практ. конф. …, 2-3 июля 1997 г. / НИИСХ Юго-Востока. - Саратов, 1997.- С.41-42.
- Вьюшков, А.А. Селекция твердой пшеницы на устойчивость к стеблевому хлебному пилильщику / А.А. Вьюшков, П.Н. Мальчиков // Научные проблемы создания новых сортов сельскохозяйственных культур, адаптированных к современным условиям производства и переработки: материалы науч. сессии, 21-22 июля 1998 г. / Сев.- Зап. науч. центр. - СПб., 1998.- С.96-98.
- Мальчиков, П.Н. Определение числа генов, контролирующих содержание каротиноидных пигментов в зерне твердой пшеницы / П.Н. Мальчиков // Теоретические и прикладные проблемы генетики, селекции и семеноводства зерновых культур: тез. науч.-практ. конф., 24-27 марта 1998 г. / НИИСХ ЦРНЗ.- Немчиновка, 1998.- С.49.
- Мальчиков, П.Н. Оценка вариантов и перспектив реализации продуктивности и адаптивности твердой пшеницы / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова // Теоретические и прикладные проблемы генетики, селекции и семеноводства зерновых культур: тез. науч.-практ. конф., 24-27 марта 1998 г. / НИИСХ ЦРНЗ.- Немчиновка, 1998. - С.50.
- Мальчиков, П.Н. Эволюция продуктивности и адаптивности твердой пшеницы в процессе селекции в условиях Среднего Поволжья / П.Н. Мальчиков, А.А. Вьюшков // Научные проблемы создания новых сортов сельскохозяйственных культур, адаптированных к современным условиям производства и переработки: материалы науч. сессии, 21-22 июля 1998 г. / Сев.-Зап. науч. центр.- СПб., 1998.- С.89-91.
- Мальчиков, П.Н. Селекция твердой пшеницы по признакам качества / П.Н. Мальчиков, А.А. Вьюшков // Научные проблемы создания новых сортов сельскохозяйственных культур, адаптированных к современным условиям производства и переработки: материалы науч. сессии, 21-22 июля 1998 г. / Сев.-Зап. науч. центр. - СПб., 1998. - С.91-94.
- Создать на основе разработанной селекционно-физиологической модели методами рекомбиногенеза и экологической селекции высокопродуктивные сорта яровой твердой пшеницы с комплексом адаптивности к условиям Поволжья, проявляющие высокий уровень гомеостаза урожайности и качества макаронных изделий: отчет о НИР (заключ.) / Самарский НИИСХ; Лаб. селекции яровой твердой пшеницы; А.А. Вьюшков, П.Н. Мальчиков, М.Г. Мясникова.- № ГР 01960010517; Инв. № 02200108292.- Безенчук, 2000.- 10 с.- Библиогр.:с.9.
- Мальчиков, П.Н. Направления селекции твердой пшеницы на жаро-засухоустойчивость / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова // Актуальные проблемы развития прикладных исследований и пути повышения их эффективности в сельскохозяйственном производстве: материалы междунар. науч.-практ. конф., посвящ. 80-летию Татарского НИИСХ, 10-12 июля 2000 г.- Казань, 2001. - С.59-60.
- Мальчиков, П.Н. Изменение высоты растений в процессе селекции яровой твердой пшеницы на продуктивность / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова // Проблемы селекции полевых культур на адаптивность и качество в засушливых условиях: сб. науч. тр. / НИИСХ Юго-Востока.- Саратов, 2001.- С.54-56.
- Пат. 0883 РФ. Пшеница твердая яровая Безенчукская 200 / А.А. Вьюшков, П.Н. Мальчиков, М.Г. Мясникова, Л.В. Туварджиева (РФ). №9808990; Заявлено 10.12.98; Зарегистр. в Гос. реестре охраняемых селекц. достижений 15.02.2001.
- Мясникова, М.Г. Перспективы сортов яровой твердой пшеницы с выполненной соломиной в Среднем Поволжье / М.Г. Мясникова, А.А. Вьюшков, П.Н. Мальчиков // Агрономическая наука в начале XXI века: материалы 40-ой науч. конф. молодых ученых аспирантов и студентов агр. фак. …, 15-17 мая 2001 г. / Пензенская ГСХА.- Пенза, 2001.- С.187-188.
- Мальчиков, П.Н. Итоги селекционно-исследовательской работы на качество по яровой твердой пшенице в Самарском НИИСХ / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова // Пути повышения и стабилизации производства высококачественного зерна: сб. докл. междунар. науч.-практ. конф., посвящ. 80-летию … Кубанского ГАУ, 12-17 июня 2002 г. Краснодар, 2002.- С.104-109.
- Вьюшков, А.А. Твердой пшеницы много не бывает: О перспективах ее производства в Поволжье / А.А. Вьюшков, П.Н. Мальчиков // Агро-Информ. - 2002. - № 49.- Ноябрь- Декабрь. - С.17-18.
- Мясникова, М.Г. Опыт селекции сортов твердой пшеницы с выполненной соломиной / М.Г. Мясникова, А.А. Вьюшков, П.Н. Мальчиков // Репродуктивная биология, генетика и селекция: сб. науч. тр., посвящ. 90-летию … С.С. Хохлова / Саратовский ГУ. - Саратов, 2002. -С.120-123.
- Шевченко, С.Н. Отзывчивость сортов зерновых культур на применение удобрений в Среднем Поволжье / С.Н. Шевченко, А.П. Чичкин, А.Ф. Сухоруков, А.А. Вьюшков, В.В. Сюков, П.Н. Мальчиков // Сорт, удобрение и защита растений в системе высокопродуктивных технологий возделывания зерновых культур: материалы Всерос. симпозиума, 24-25 июня 2002 г. / НИИСХ ЦРНЗ, ВНИПТИХИМ.- М., 2002. -С.242-247.
- Вьюшков, А.А. Селекция яровой твердой пшеницы / А.А. Вьюшков, П.Н. Мальчиков // Вековой юбилей: Научно-исторический очерк о прошлом и настоящем Самарского НИИСХ им. Н.М. Тулайкова.- Самара: Изд-во «НТЦ», 2003.- С. 41-48.
- Вьюшков, А.А. Итоги селекции яровой твердой пшеницы / А.А. Вьюшков, П.Н. Мальчиков // Генетика, селекция и семеноводство сельскохозяйственных культур: сб. науч. тр.: к 100-летию Самарского НИИСХ.- Самара: Изд-во «НТЦ», 2003.- С.60-88: табл., рис.
- Вьюшков, А.А. Сорта яровой мягкой и твердой пшеницы / А.А. Вьюшков, В.В. Сюков, С.Н. Шевченко, П.Н. Мальчиков // Коммерческие сорта полевых культур Российской Федерации.- М., 2003.- С.85-89; С.106-110: рис.
- Мальчиков, П.Н. Перспективы селекции сортов твердой пшеницы устойчивых к хлебному пилильщику Cehpus pygmaeus L.: рек. / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова; Самарский НИИСХ. - Безенчук, 2005. -10с.: табл.
- Мальчиков, П.Н. Обоснование приоритетов в селекции твердой пшеницы в условиях лимитирования продукционного процесса абиотическими факторами (модель сорта): рек. / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова; Самарский НИИСХ.- Самара, 2005. -17с.: табл.
- Мальчиков, П.Н. Создание доноров генов редукции высоты растений, жаростойкости, процессов гаметогенеза и устойчивости твердой пшеницы к возбудителям грибковых болезней Blumeria graminis, Puccinia recondita, Ustilago tritici: метод. руковод. / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова; Самарский НИИСХ.- Безенчук, 2007.- 23 с.:табл.
- Мальчиков, П.Н. Селекция твердой пшеницы в Среднем Поволжье на устойчивость к грибным болезням / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова // Пути решения проблем повышения адаптивности, продуктивности и качества зерновых и кормовых культур: материалы междунар. науч.-практ. конф., посвящ. 100-летию Самарского НИИСХ … и 70-летию Поволжского НИИСС …. - Самара, 2003. - С. 37-38.
- Мальчиков, П.Н. Селекция твердой пшеницы на урожайность / П.Н. Мальчиков, А.А. Вьюшков // Генетика, селекция и семеноводство сельскохозяйственных культур: сб. науч. тр.: К 100-летию Самарского НИИСХ.- Самара: Изд-во «НТЦ», 2003. - С. 89 -118: табл. рис.- Библиогр.: с. 117-118.
- Мальчиков, П.Н. Лаборатория селекции и семеноводства яровой твердой пшеницы / П.Н. Мальчиков // Самарский научно-исследовательский институт сельского хозяйства имени академика Н.М. Тулайкова: 1903 - 2003: проспект: [К 100-летию со дня образования]. -Самара, 2003. -С. 16-17: фото.
- Мальчиков, П.Н. Сорта твердой пшеницы Самарского НИИСХ за 90 лет работы: их свойства и методы выведения / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова // Селекция и семеноводство сельскохозяйственных культур: сб. материалов VII Всерос. науч.-практ. конф., 18-19 февраля 2003 г. / Пензенская ГСХА.- Пенза, 2003.- С. 33-34.
- Мясникова, М.Г. Исходный селекционный материал в селекции сортов твердой пшеницы устойчивых к хлебному пилильщику CEPHUS PYGMAEUS L. / М.Г. Мясникова, П.Н. Мальчиков // Пути решения проблем повышения адаптивности, продуктивности и качества зерновых и кормовых культур: материалы междунар. науч.-практ. конф., посвящ. 100-летию Самарского НИИСХ … и 70-летию Поволжского НИИСС …. - Самара, 2003. - С. 35-36.
- Мальчиков, П.Н. Твердая пшеница Безенчукская 200 / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова // Селекция и семеноводство. - 2004. - № 4. - С. 8-9.
- Мальчиков, П.Н. Диверсификация системы сортов твердой пшеницы в Среднем Поволжье / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова // Стратегия адаптивной селекции полевых культур в связи с глобальным изменением климата: сб. науч. тр. по материалам Междунар. науч.-практ. конф. г.Саратов, 16-18 июня 2004г. / НИИСХ Юго-Востока. - Саратов, 2004. - С. 41-48: табл.
- Пат. 2527 Российская Федерация. Пшеница твердая яровая Безенчукская 202 / А.А. Вьюшков, П.Н. Мальчиков, М.Г. Мясникова, С.Е. Поротькин; заявитель и патентообладатель ГНУ Самарский НИИСХ.- № 9811633; заявл. 06.12.01; зарегистр. в Гос. реестре охраняемых селекц. достижений 18.02.05.
- Мясникова, М.Г. Селекционно-генетическая оценка признака выполненности соломины у твердой пшеницы / М.Г. Мясникова, А.А. Вьюшков, П.Н. Мальчиков // Пути мобилизации биологических ресурсов повышения продуктивности пашни, энергоресурсосбережения и производства конкурентоспособной продукции: материалы Всерос. науч.-практ. конф., посвящ. 85-летию ТатНИИСХ и 1000-летию Казани, 5-6 июля 2005 г. - Казань, 2005.- С. 141-146: табл.
- Паспорта доноров селекционно-ценных признаков сельскохозяйственных культур (выделенных и созданных в 2002-2005 гг.). Вып. 19: Пшеница, тритикале, рожь, ячмень, овес, кукуруза... / К.З. Будин, А.А. Вьюшков, Д.Е. Зубов, П.Н. Мальчиков, А.В. Милехин, М.Г. Мясникова, С.Е. Поротькин, В.В. Сюков, С.Н. Шевченко и др. / ВИР. - СПб., 2006. - 61 с.
- Мальчиков, П.Н. Совершенствование блока коадаптированных генов продуктивности и стабильности безенчукских сортов яровой твердой пшеницы / П.Н. Мальчиков, А.А. Вьюшков, М.Г. Мясникова, Н.П. Чиганцев // Достижения науки и техники АПК.- 2007.- № 8.- С. 4-7: табл.
- Мальчиков, П.Н. Селекционная оценка интегральных компонентов продукционного процесса у Triticum durum Desf. / П.Н. Мальчиков // Известия Самарского НЦ РАН: спец. вып.: «Развитие научного наследия академика Николая Максимовича Тулайкова»: (К 105-летию Самарского научно-исследовательского института сельского хозяйства им. Н.М. Тулайкова).- Самара: СамНЦ РАН, 2008. - С. 124-132: табл.- Библиогр.: с. 131-132.
- Мальчиков, П.Н. Наследование выполненности соломины у Triticum durum Desf / П.Н. Мальчиков, М.Г. Мясникова // Известия Самарского НЦ РАН: спец. вып.: «Развитие научного наследия академика Николая Максимовича Тулайкова»: (К 105-летию Самарского научно-исследовательского института сельского хозяйства им. Н.М. Тулайкова). - Самара: СамНЦ РАН, 2008. - С. 133-141: табл., рис.- Библиогр.: с.140-141.
- А.с. 42405 РФ. Пшеница твердая яровая Безенчукская 205 / А.А. Вьюшков, П.Н. Мальчиков, М.Г. Мясникова (РФ). - № 9553783; заявл. 16.12.04; зарегистр. в Гос. реестре охраняемых селекц. достижений 14.05.08.
- Пат. 3915 Российская Федерация. Пшеница твердая яровая Безенчукская 205 / А.А. Вьюшков, П.Н. Мальчиков, М.Г. Мясникова; заявитель и патентообладатель ГНУ Самарский НИИСХ.- № 9553783; заявл. 16.12.04; зарегистр. в Гос. реестре охраняемых селекц. достижений 14.05.08.
- Основные пути повышения эффективности растениеводства Самарской области: науч.-практ. рек. / С.Н. Шевченко, А.В. Милехин, В.А. Корчагин, А.А. Вьюшков, А.П. Чичкин, А.Ф. Сухоруков, В.В. Сюков, П.Н. Мальчиков и др.; Самарский НИИСХ.- Самара, 2008. - 131 с.: табл., рис.+прил.
- Мальчиков П.Н. Доноры генов гомеостаза и элементов продуктивности твердой пшеницы /П.Н.Мальчиков // Нива Поволжья (в печати)
- Мальчиков П.Н. Перспективы редукции высоты растений в селекции яровой твердой пшеницы./П.Н.Мальчиков // Нива Поволжья (в печати)
- Мальчиков П.Н. Система взаимоотношений фотосинтеза, ростовых процессов, потребления минеральных веществ при формировании генотипических различий по крожайности яровой твердой пшеницы /П.Н.Мальчиков // Достижения науки и техники АПК.- 2009.-№7.
- Мальчиков П.Н. Проблемы селекции высокобелковистых и урожайных сортов твердой пшеницы // Достижения науки и техники АПК.-2009.- №8.
- Мальчиков П.Н. Яровая твердая пшеница Безенчукская 205 /П.Н.Мальчиков, А.А.Вьюшков, М.Г.Мясникова // Зерновое хозяйство.- 2009. №4.
- Мальчиков П.Н. Яровая твердая пшеница Безенчукская степная /П.Н.Мальчиков, А.А.Вьюшков, М.Г.Мясникова //Зерновое хозяйство.- 2009 №4.
- Мальчиков П.Н. Доноры генов редукции высоты растений твердой пшеницы /П.Н.Мальчиков // Самара: СамНЦ РАН, 2009. (в печати).
- Мальчиков П.Н. Подбор родительских сортов для гибридизации в селекции яровой твердой пшеницы / П.Н.Мальчиков // Достижения науки и техники АПК.-2009.- №11.
- Мальчиков П.Н. Новые сорта яровой твердой пшеницы / П.Н.Мальчиков, А.А.Вьшков, М.Г.Мясеикрва // Достижения науки и техники АПК.-2009.- №11.